
Bacterial Spore-Based Delivery System: 20 Years of a Versatile Approach for Innovative Vaccines


Abstract
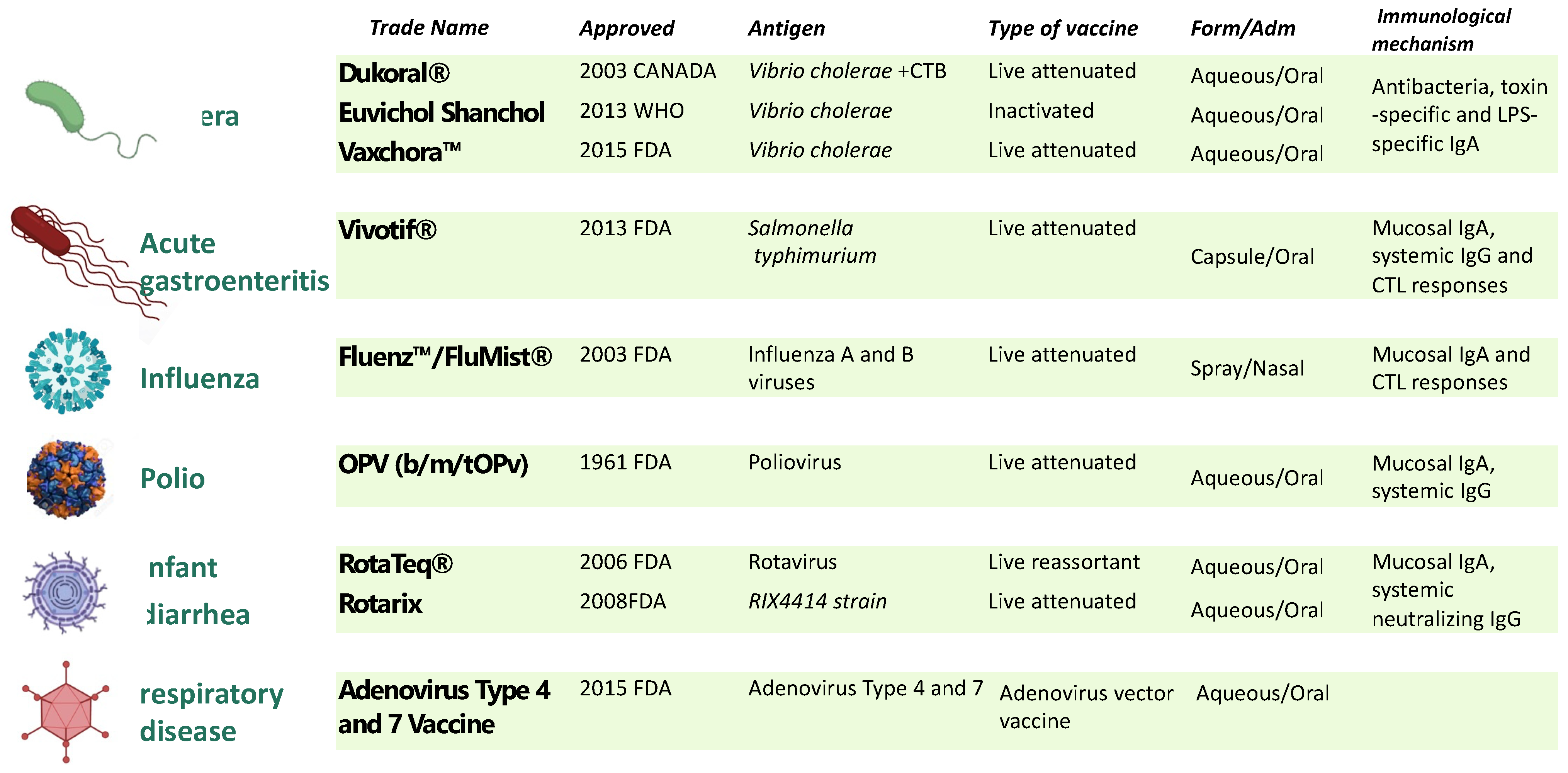
:1. Introduction
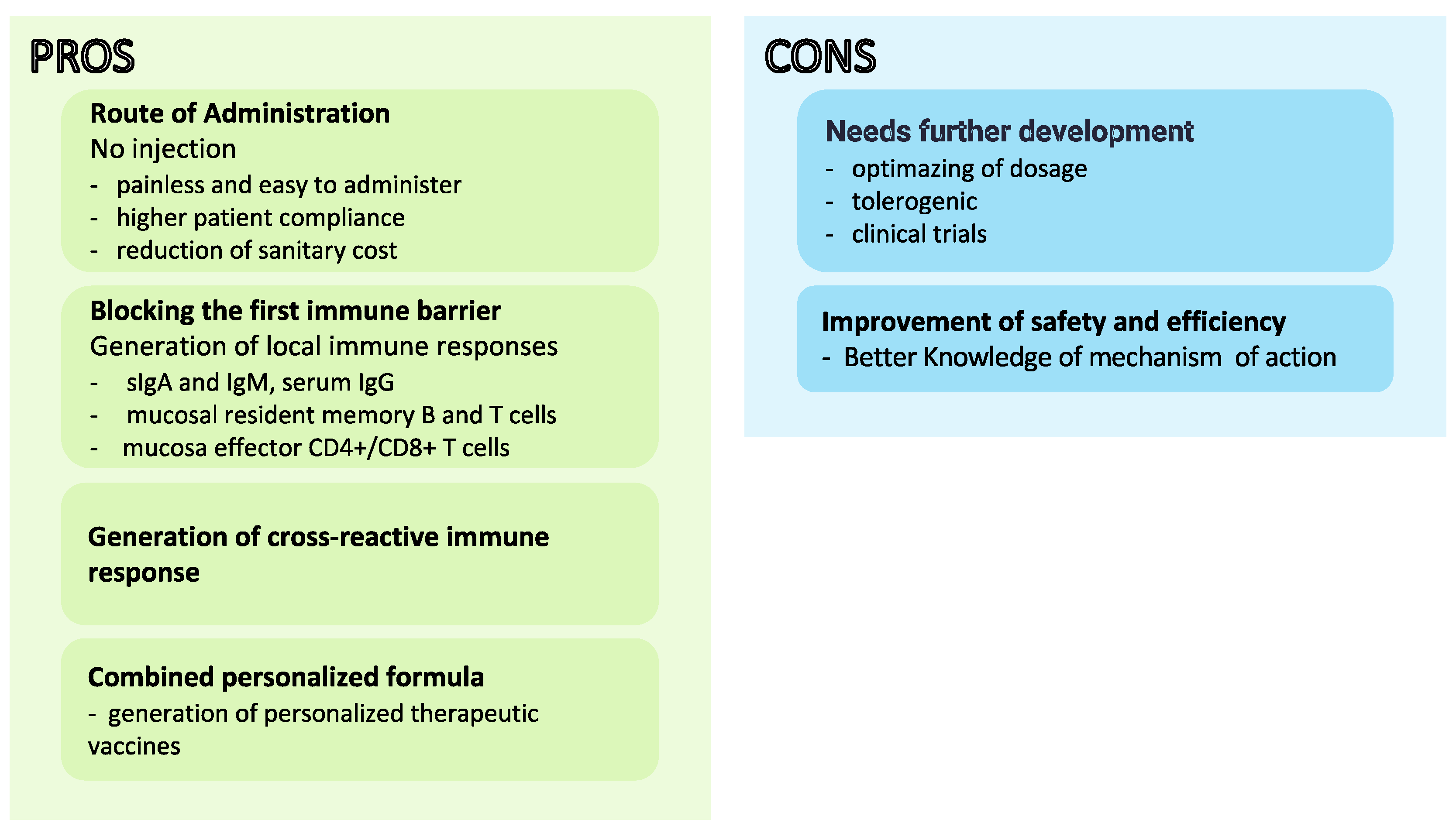
2. Bacterial Spores as a Mucosal Vaccine Delivery System
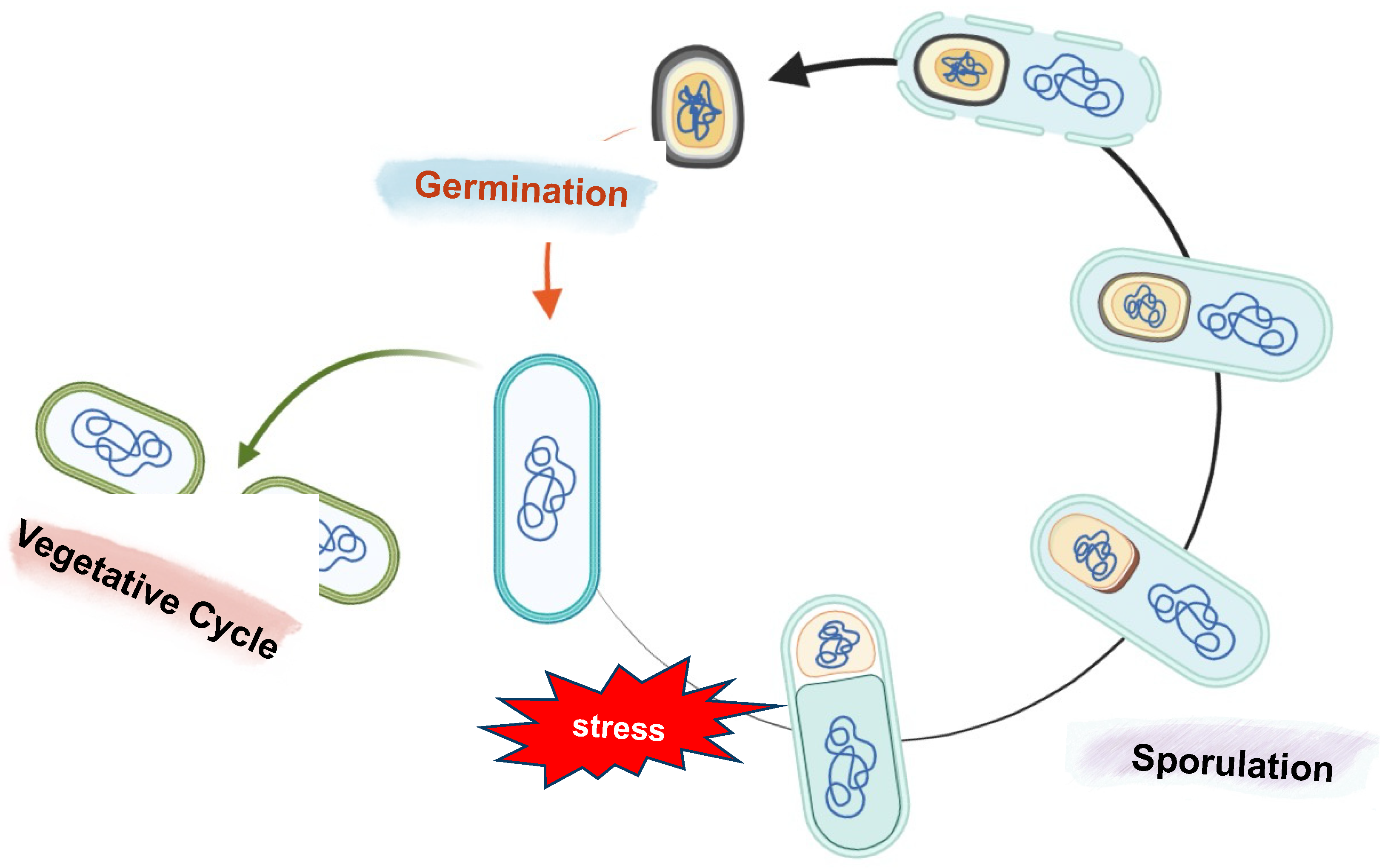
2.1. Bacterial Spores, Sporulation, and Intestinal Life Cycle
2.2. Spore Surface
2.3. Bacterial Spore as a Recombinant Vaccine Platform
- (1)
- The extreme stability due to the well-documented resistance of the bacterial spore to high temperatures, acidic pH, and the presence of chemicals and enzymes [52,53]. Guaranteeing the high stability of the vaccine carrier system from production to administration to the patient is a crucial requirement of vaccine development. Bacillus spores are more stable than vegetative cells during the processing and storage stages of commercial preparations, making them a suitable candidate for vaccine formulations [92]. Moreover, stability at extreme temperatures is preferred in the development of mucosal vaccines, mainly for those intended for use in developing countries, where poor distribution and storage conditions are the main limitations [6,91,93];
- (2)
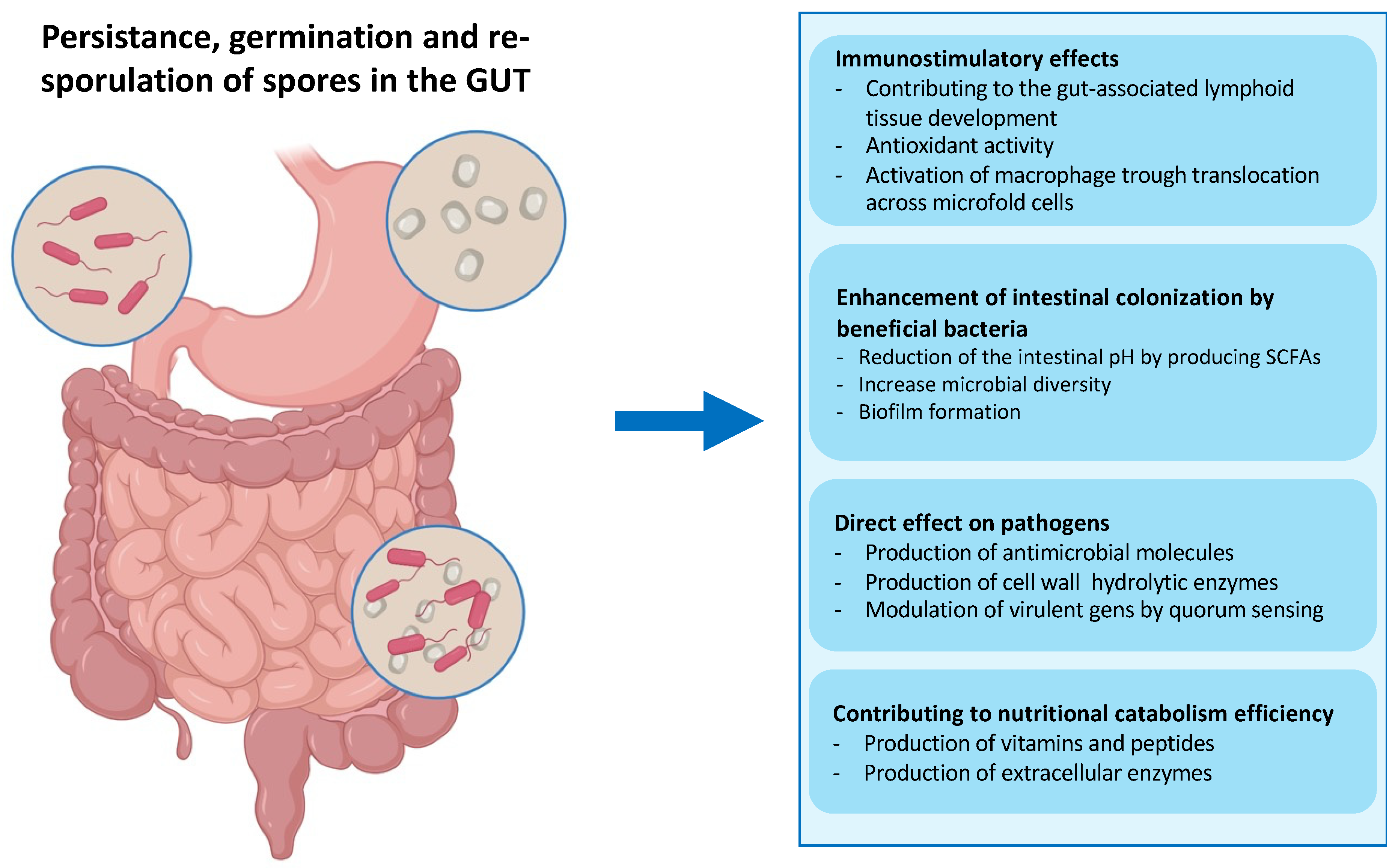
- The exceptional safety record of several spore former species used worldwide in probiotic preparations for human and animal use, as dietary supplements and growth promoters [66,67,68,69,70,92,94]. As mentioned above, several spore former species are part of the animal gut microbiota, which have a role in the development of the immune system, protection against intestinal pathogens, induction of cytoprotective responses, and of anti-oxidative stress responses in epithelial cells (Figure 4) [64,65,66,94]. This safety record is an essential requirement if the display system is intended for the delivery of antigen molecules to human mucosal surfaces;
- (3)
- All known coat proteins are synthesized in the mother compartment during sporulation [54,56,72]. Consequently, coat components and the antigens fused to them do not need to undergo a cell wall translocation step to expose the recombinant proteins externally, thus overcoming the size limitation often encountered with cell-based display systems [52,91,95];
- (4)
2.4. Spore-Based Vaccine Design Strategy
Anchor Proteins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anchor [Refs] | Target Pathogen | Antigen | Fusion Method | Linker | Application |
|---|---|---|---|---|---|
| CotB | |||||
| [52] | Clostridium tetani | C-term fragment of the tetanus toxin, TTFC | C-term, N-term, sandwich | - | Oral vaccination for tetanus |
| [99] | Clostridium difficile | fagellin protein, FliD | C-terminal | GGGEA; AAKGGG | C. difficile oral vaccine |
| [108] | C-term repeat domains of toxins A and B, TcdA-TcdB | C-terminal | - | ||
| [106] | Clostridium perfringens | C-term of alpha toxin gene, Cpa247-370 fused to the GST gene | C-terminal | - | Oral and Nasal Vaccine against necrotic enteritis |
| [95,105] | Bacillus anthracis | anthrax-protective antigen, PA | C-terminal | - | Anthrax vaccine |
| [107] | Helicobacter acinonychis | urease subunit alpha, UreA | C-terminal | GGGEAA; AKGGG | Anti-Helicobacter vaccine |
| [121] | Helicobacter pylori | vacuolating cytotoxin A, CagA | C-terminal N-terminal | GGGGS | Anti-Helicobacter vaccine |
| [122] | Mycobacterium tuberculosis | immunodominant secretory antigen, MPT64 | C-terminal | - | Nasal Vaccine against tuberculosis |
| [100] | Streptococcus mutans | truncated P1 protein | N-terminal | NR | S. mutans vaccine |
| [109] | White spot syndrome virus | major envelope proteins, VP28 | C-terminal | - | Oral vaccine for shrimps |
| [123] | Influenza virus | ectodomain of influenza virus M2 protein | C-terminal | - | Oral Influenza vaccine |
| [124] | Adjuvant | human IL-2 | C-terminal | Adjuvant to H. pylori vaccine | |
| CotC | |||||
| [110] | Clostridium tetani | C-term fragment of the tetanus toxin, TTFC | C-terminal N-terminal | - | Oral vaccination for tetanus |
| [108] | Clostrium difficile | C-term repeat domains of toxins A and B TcdA-TcdB | C-terminal | - | C. dìfficile oral vaccine |
| [110] | Escherichia coli | heat-labile enterotoxin B, LTB | C-terminal N-terminal | - | E. coli vaccine |
| [125] | Salmonella serovar pullorum | outer membrane protein (porin), OmpC | C-terminal | - | Salmonella vaccine |
| [95,105] | Bacillus anthracis | anthrax-protective antigen, PA | C-terminal | - | Anthrax vaccine |
| [67] | Helicobacter acinonychis | urease subunit alpha, UreA | C-terminal | - | Anti-Helicobacter vaccine |
| [124] | Helicobacter pylori | urease subunit beta, UreB | C-terminal | - | Oral vaccine for H. pylori |
| [121] | vacuolating cytotoxin A, CagA | C-terminal N-terminal | GGGGS | ||
| [126] | cholera toxin B subunit, CTB and UreB | C-terminal | - | ||
| [112] | Clonorchis sinensis | tegumental protein 20.8 kD, TP20.8 | C-terminal | - | Liver flukes vaccine |
| [112] | tegumental protein 22.3 kDa, CsTP22.3 | C-terminal | - | ||
| [113] | cysteine proteases, CsCP | C-terminal | - | ||
| [114] | leucine aminopeptidase 2, CsLAP2 | C-terminal | - | ||
| [115] | enolase | C-terminal | - | ||
| [116] | paramyosin antigen, CsPmy | C-terminal | |||
| [117] | serpin, CsSer-3 | C-terminal | |||
| [127] | Schistosoma japonicum | 26 kDa full- length GST protein, SjGST | C-terminal | - | Liver flukes oral vaccine |
| [128] | grass carp reovirus | major outer capsid protein, VP4 | C-terminal | - | Grass carp reovirus vaccine |
| [129] | White spot syndrome virus | major envelope proteins, VP28 and VP62 | C-terminal | - | Oral vaccine for shrimps |
| [130] | Bombyx mori | nucleopolyhedrovirus, GP64 | C-terminal | - | Bombyx mori vaccine |
| CotG | |||||
| [107] | Helicobacter acinonychis | urease subunit alpha, UreA | C-terminal | - | Anti-Helicobacter vaccine |
| [99] | Clostridium difficile | fagellin protein, FliD | C-terminal | GGGEA AAKGGG | Oral vaccine against C. difficile |
| [121] | Helicobacter pylori | vacuolating cytotoxin A, CagA | C-terminal N-terminal | GGGGS | Anti-Helicobacter vaccine |
| [120] | Transmissible gastroenteritis virus | transmissible gastroenteritis virus spike, TGEV-S | C-terminal | Transmissible gastroenteritis vaccine | |
| [118] | - | Streptavidin | C-terminal | GGGGS | - |
| CotZ | |||||
| [99] | Clostridium difficile | fagellin protein, FliD | C-terminal | GGGEA AAKGGG | Oral vaccine against C. difficile |
| [107] | Helicobacter acinonychis | urease subunit alpha, UreA | C-terminal | GGGGS | Anti-Helicobacter vaccine |
| [131] | Helicobacter pylori | vacuolating cytotoxin A, CagA | C-terminal N-terminal | GGGEAAAKGGG | Anti-Helicobacter vaccine |
| CgeA | |||||
| [121] | Helicobacter pylori | vacuolating cytotoxin A, CagA | C-terminal N-terminal | GGGEAAAKGGG | Anti-Helicobacter vaccine |
2.5. Strategies for Optimizing Antigen Exposure on Spore Surface
2.5.1. Linker Peptides to Increase Stability and Flexibility of Recombinant Protein
2.5.2. Multi-Antigen Spore-Based Mucosal Vaccine
2.5.3. Non-Recombinant Display of Antigens on the Spore Surface
3. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dey, J.; Mahapatra, S.R.; Lata, S.; Patro, S.; Misra, N.; Suar, M. Exploring Klebsiella pneumoniae capsule polysaccharide proteins to design multiepitope subunit vaccine to fight against pneumonia. Exp Rev. Vaccines. 2022, 21, 569–587. [Google Scholar] [CrossRef] [PubMed]
- Woodworth, J.S.; Clemmensen, H.S.; Battey, H.; Dijkman, K.; Lindenstrøm, T.; Laureano, R.S.; Taplitz, R.; Morgan, J.; Aagaard, C.; Rosenkrands, I.; et al. A Mycobacterium tuberculosis-specific subunit vaccine that provides synergistic immunity upon co-administration with Bacillus Calmette-Guérin. Nat. Commun. 2021, 12, 6658. [Google Scholar] [CrossRef]
- Roukens, A.H.E.; van Halem, K.; de Visser, A.W.; Visser, L.G. Long-Term Protection After Fractional-Dose Yellow Fever Vaccination: Follow-up Study of a Randomized, Controlled, Noninferiority Trial. Ann. Intern Med. 2018, 169, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Klein, N.P.; Abu-Elyazeed, R.; Cheuvart, B.; Janssens, W.; Mesaros, N. Immunogenicity and safety following primary and booster vaccination with a hexavalent diphtheria, tetanus, acellular pertussis, hepatitis B, inactivated poliovirus and Haemophilus influenzae type b vaccine: A randomized trial in the United States. Hum. Vaccines Immunother. 2019, 15, 809–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Global Task Force on Cholera Control: Overview of Ending Cholera—A Global Roadmap to 2030; WHO: Geneva, Switzerland, 2018.
- Pavot, V.; Rochereau, N.; Genin, C.; Verrier, B.; Paul, S. New insights in mucosal vaccine development. Vaccine 2012, 30, 142–154. [Google Scholar] [CrossRef]
- Chowdhury, F.; Ali Syed, K.; Akter, A.; Rahman Bhuiyan, T.; Tauheed, I.; Khaton, F.; Biswas, R.; Ferdous, J.; Al Banna, H.; Ross, A.G.; et al. A phase I/II study to evaluate safety, tolerability and immunogenicity of Hillchol®, an inactivated single Hikojima strain based oral cholera vaccine, in a sequentially age descending population in Bangladesh. Vaccine 2021, 39, 4450–4457. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, T.R.; Choudhury, F.K.; Khanam, F.; Saha, A.; Sayeed, M.A.; Salma, U.; Lundgren, A.; Sack, D.A.; Svennerholm, A.M.; Qadri, F. Evaluation of immune responses to an oral typhoid vaccine, Ty21a, in children from 2 to 5 years of age in Bangladesh. Vaccine 2014, 32, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Carter, N.J.; Curran, M.P. Live Attenuated Influenza Vaccine (FluMist®; Fluenz™). Drugs 2011, 71, 1591–1622. [Google Scholar] [CrossRef]
- Yeh, M.T.; Bujaki, E.; Dolan, P.T.; Smith, M.; Wahid, R.; Konz, J.; Weiner, A.J.; Bandyopadhyay, A.S.; Van Damme, P.; De Coster, I.; et al. Engineering the Live-Attenuated Polio Vaccine to Prevent Reversion to Virulence. Cell Host Microbe 2020, 27, 736–751.e8. [Google Scholar] [CrossRef]
- Vesikari, T.; Itzler, R.; Karvonen, A.; Korhonen, T.; Van Damme, P.; Behre, U.; Bona, G.; Gothefors, L.; Heaton, P.M.; Dallas, M.; et al. RotaTeq, a pentavalent rotavirus vaccine: Efficacy and safety among infants in Europe. Vaccine 2019, 28, 345–351. [Google Scholar] [CrossRef]
- Collins, N.D.; Adhikari, A.; Yang, Y.; Kuschner, R.A.; Karasavvas, N.; Binn, L.N.; Walls, S.D.; Graf, P.C.F.; Myers, C.A.; Jarman, R.G.; et al. Live Oral Adenovirus Type 4 and Type 7 Vaccine Induces Durable Antibody Response. Vaccines 2020, 8, 411. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, E.W. Mucosal vaccines—Fortifying the frontiers. Nat. Rev. Immunol. 2022, 2, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Kotomina, T.; Korenkov, D.; Matyushenko, V.; Prokopenko, P.; Rudenko, L.; Isakova-Sivak, I. Live attenuated influenza vaccine viral vector induces functional cytotoxic T-cell immune response against foreign CD8+ T-cell epitopes inserted into NA and NS1 genes using the 2A self-cleavage site. Hum. Vaccines Immunother. 2018, 14, 2964–2970. [Google Scholar] [CrossRef] [Green Version]
- Tsakiri, M.; Naziris, N.; Demetzos, C. Innovative vaccine platforms against infectious diseases: Under the scope of the COVID-19 pandemic. Int. J. Pharm. 2021, 610, 121212. [Google Scholar] [CrossRef] [PubMed]
- Buddy Creech, C.; Jimenez-Truque, N.; Kown, N.; Sokolow, K.; Brady, E.J.; Yoder, S.; Solovay, K.; Rubin, K.; Noviello, S.; Hensel, E.; et al. Safety and immunogenicity of live, attenuated intranasal Bordetella pertussis vaccine (BPZE1) in healthy adults. Vaccine 2022, 40, 6740–6746. [Google Scholar] [CrossRef]
- Pei, Z.; Jiang, X.; Yang, Z.; Ren, X.; Gong, H.; Reeves, M.; Sheng, J.; Wang, Y.; Pan, Z.; Liu, F.; et al. Oral Delivery of a Novel Attenuated Salmonella Vaccine Expressing Influenza A Virus Proteins Protects Mice against H5N1 and H1N1 Viral Infection. PLoS ONE 2015, 10, e0129276. [Google Scholar] [CrossRef]
- 10Smith, G.L.; MacKett, M.; Moss, B. Infectious vaccinia virus recombinants that express hepatitis B virus surface antigen. Nature 1983, 302, 490–495. [Google Scholar] [CrossRef]
- Leong, K.H.; Ramsay, A.J.; Boyle, D.B.; Ramshaw, I.A. Selective induction of immune responses by cytokines coexpressed in recombinant fowlpox virus. J. Virol. 1994, 68, 8125–8130. [Google Scholar] [CrossRef] [Green Version]
- Moss, B.; Smith, G.L.; Gerin, J.L.; Purcell, R.H. Live recombinant vaccinia virus protects chimpanzees against hepatitis B. Nature 1984, 311, 67–69. [Google Scholar] [CrossRef]
- Draper, S.J.; Heeney, J.L. Viruses as vaccine vectors for infectious diseases and cancer. Nat. Rev. Microbiol. 2012, 8, 62–73. [Google Scholar] [CrossRef]
- Tan, H.X.; Gilbertson, B.P.; Jegaskanda, S.; Alcantara, S.; Amarasena, T.; Stambas, J.; McAuley, J.L.; Kent, S.J.; De Rose, R. Recombinant influenza virus expressing HIV-1 p24 capsid protein induces mucosal HIV-specific CD8 T-cell responses. Vaccine 2016, 34, 1172–1179. [Google Scholar] [CrossRef]
- Jas, D.; Coupier, C.; Toulemonde, C.E.; Guigal, P.M.; Poulet, H. Three-year duration of immunity in cats vaccinated with a canarypox-vectored recombinant rabies virus vaccine. Vaccine 2012, 30, 6991–6996. [Google Scholar] [CrossRef] [PubMed]
- Esaki, M.; Godoy, A.; Rosenberger, J.K.; Rosenberger, S.C.; Gardin, Y.; Yasuda, A.; Dorsey, K.M. Protection and antibody response caused by turkey herpesvirus vector newcastle disease vaccine. Avian Dis. 2013, 57, 750–755. [Google Scholar] [CrossRef] [PubMed]
- FDA. First FDA-Approved Vaccine for the Prevention of Ebola Virus Disease, Marking a Critical Milestone in Public Health Preparedness and Response. 2020. Available online: https://www.fda.gov/news-events/press-announcements/first-fda-approved-vaccine-prevention (accessed on 19 December 2019).
- Afkhami, S.; D’Agostino, M.R.; Zhang, A.; Stacey, H.D.; Marzok, A.; Kang, A.; Singh, R.; Bavananthasivam, J.; Ye, G.; Luo, X.; et al. Respiratory mucosal delivery of next-generation COVID-19 vaccine provides robust protection against both ancestral and variant strains of SARS-CoV-2. Cell 2022, 185, 896–915. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Sayedahmed, E.E.; Singh, V.K.; Mishra, A.; Dorta-Estremera, S.; Nookala, S.; Canaday, D.H.; Chen, M.; Wang, J.; Sastry, K.J.; et al. A recombinant bovine adenoviral mucosal vaccine expressing mycobacterial antigen-85B generates robust protection against tuberculosis in mice. Cell reports. Medicine 2021, 2, 100372. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.; Wang, L.R.; Gómez-Román, V.R.; Davis-Warren, A.; Montefiori, D.C.; Kalyanaraman, V.S.; Venzon, D.; Zhao, J.; Kan, E.; Rowell, T.J.; et al. Replicating Rather than Nonreplicating Adenovirus- Human Immunodeficiency Virus Recombinant Vaccines Are Better at Eliciting Potent Cellular Immunity and Priming High-titer Antibodies. J. Virol. 2005, 79, 10200–10209. [Google Scholar] [CrossRef] [Green Version]
- Cárcamo-Calvo, R.; Muñoz, C.; Buesa, J.; Rodríguez-Díaz, J.; Gozalbo-Rovira, R. The Rotavirus Vaccine Landscape, an Update. Pathogens 2021, 10, 520. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Yang, Y.; Liang, Z.; Tian, Y.; Liu, B.; Gao, Z.; Jia, L.; Chen, L.; Wang, Q. Effectiveness of Lanzhou lamb rotavirus vaccine in preventing gastroenteritis among children younger than 5 years of age. Vaccine 2019, 37, 3611–3616. [Google Scholar] [CrossRef]
- Mielcarek, N.; Alonso, S.; Locht, C. Nasal vaccination using live bacterial vectors. Adv. Drug Deliv. Rev. 2001, 51, 55–69. [Google Scholar] [CrossRef]
- Chin’ombe, N.; Ruhanya, V. Recombinant Salmonella Bacteria Vectoring HIV/AIDS Vaccines. Open Virol. J. 2013, 7, 121–126. [Google Scholar] [CrossRef] [Green Version]
- Detmer, A.; Glenting, J. Live bacterial vaccines-a review and identification of potential hazards. Microb. Cell Factories 2006, 23, 5–23. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Berzofsky, J.A. Oral vaccines: Directed safe passage to the front line of defense. Gut Microbes 2013, 4, 246–252. [Google Scholar] [CrossRef] [Green Version]
- Bontempi, I.; Leal, K.; Prochetto, E.; Díaz, G.; Cabrera, G.; Bortolotti, A.; Morbidoni, H.R.; Borsuk, S.; Dellagostin, O.; Marcipar, I. Recombinant Mycobacterium bovis BCG is a promising platform to develop vaccines against Trypansoma cruzi infection. Clin. Exp. Immunol. 2020, 201, 306–316. [Google Scholar] [CrossRef]
- Kanno, A.I.; Goulart, C.; Rofatto, H.K.; Oliveira, S.C.; Leite, L.C.C.; McFadden, J. New Recombinant Mycobacterium bovis BCG Expression Vectors: Improving Genetic Control over Mycobacterial Promoters. Appl. Environ. Microbiol. 2016, 82, 2240–2246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruhn, K.W.; Craft, N.; Miller, J.F. Listeria as a vaccine vector. Microbes Infect. 2007, 9, 1226–12352007. [Google Scholar] [CrossRef] [PubMed]
- Frankel, F.R.; Hegde, S.; Lieberman, J.; Paterson, Y. Induction of Cell-Mediated Immune Responses to Human Immunodeficiency Virus Type 1 Gag Protein by Using Listeria monocytogenes as a Live Vaccine Vector. J. Immunol. 1995, 155, 4775–4782. [Google Scholar] [CrossRef]
- Curtiss, R., III; Xin, W.; Li, Y.; Kong, W.; Wanda, S.Y.; Gunn, B.; Wang, S. New technologies in using recombinant attenuated Salmonella vaccine vectors. Crit. Rev. Immunol. 2010, 30, 255–270. [Google Scholar] [CrossRef] [Green Version]
- Zhi, Y.; Ji, H.J.; Guo, H.; Lim, J.H.; Byun, E.B.; Kim, W.S.; Seo, H.S. Salmonella Vaccine Vector System for Foot-and-Mouth Disease Virus and Evaluation of Its Efficacy with Virus-Like Particles. Vaccines 2021, 9, 22. [Google Scholar] [CrossRef]
- Roland, K.L.; Tinge, S.A.; Killeen, K.P.; Kochi, S.K. Recent advances in the development of live, attenuated bacterial vectors. Curr. Opin. Mol. Ther. 2007, 7, 62–72. [Google Scholar]
- Nuorti, J.P.; Whitney, C.G.; Centers for Disease Control and Prevention (CDC). Prevention of pneumococcal disease among infants and children—Use of 13-valent pneumococcal conjugate vaccine and 23-valent pneumococcal polysaccharide vaccine—Recommendations of the Advisory Committee on Immunization Practices (ACIP). MMWR. Recommendations and reports: Morbidity and mortality weekly report. Recomm. Rep. 2010, 59, 1–18. [Google Scholar]
- Sundararaman, A.M.; Ray, P.V.; Ravindra, J.; Halami, P.M. Role of probiotics to combat viral infections with emphasison COVID-19. Appl. Microbiol. Biotechnol. 2020, 104, 8089–8104. [Google Scholar] [CrossRef] [PubMed]
- Niu, H.; Xing, J.H.; Zou, B.S.; Shi, C.W.; Huang, H.B.; Jiang, Y.L.; Wang, J.Z.; Cao, X.; Wang, N.; Zeng, Y.; et al. Immune Evaluation of Recombinant Lactobacillus plantarum With Surface Display of HA1-DCpep in Mice. Front. Immunol. 2021, 12, 800965. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.H.; Yang, W.T.; Yang, G.L.; Zhang, X.K.; Liu, Y.Y.; Zhang, L.J.; Ye, L.P.; Hu, J.T.; Xing, X.; Qi, C.; et al. Lactobacillus plantarum vaccine vector expressing hemagglutinin provides protection against H9N2 challenge infection. Virus Res. 2016, 211, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, B.; Zhang, D.; Liu, S.; Ren, J. The construction of recombinant Lactobacillus casei vaccine of PEDV and its immune responses in mice. BMC Vet Res. 2021, 17, 184. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Yang, G.; Wang, Q.; Wang, Z.; Yang, W.; Gu, W.; Shi, C.; Wang, J.; Huang, H.; Wang, C. Molecular mechanisms underlying protection against H9N2 influenza virus challenge in mice by recombinant Lactobacillus plantarum with surface displayed HA2-LTB. J. Biotechnol. 2017, 259, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Fu, T.; Hao, J.; Li, L.; Tian, M.; Jin, N.; Ren, L.; Li, C. A recombinant Lactobacillus plantarum strain expressing the spike protein of SARS-CoV-2. Int. J. Biol. Macromol. 2020, 160, 36–740. [Google Scholar] [CrossRef]
- Xu, J.; Ren, Z.; Cao, K.; Li, X.; Yang, J.; Luo, X.; Zhu, L.; Wang, X.; Ding, L.; Liang, J.; et al. Boosting Vaccine-Elicited Respiratory Mucosal and Systemic COVID-19 Immunity in Mice With the Oral Lactobacillus plantarum. Front. Nutr. 2021, 8, 78924. [Google Scholar] [CrossRef]
- Vetráková, A.; Chovanová, R.K.; Rechtoríková, R.; Krajčíková, D.; Barák, I. Bacillus subtilis spores displaying RBD domain of SARS-CoV-2 spike protein. Comput. Struct. Biotechnol. J. 2023, 21, 1550–1556. [Google Scholar] [CrossRef]
- Katsande, P.M.; Fernández-Bastit, L.; Ferreira, W.T.; Vergara-Alert, J.; Hess, M.; Lloyd-Jones, K.; Hong, H.A.; Segales, J.; Cutting, S.M. Heterologous Systemic Prime-Intranasal Boosting Using a Spore SARS-CoV-2 Vaccine Confers Mucosal Immunity and Cross-Reactive Antibodies in Mice as well as Protection in Hamsters. Vaccines 2022, 10, 1900. [Google Scholar] [CrossRef]
- Isticato, R.; Cangiano, G.; Tran, H.T.; Ciabattini, A.; Medaglini, D.; Oggioni, M.R.; De Felice, M.; Pozzi, G.; Ricca, E. Surface display of recombinant proteins on Bacillus subtilis spores. J. Bacteriol. 2001, 183, 6294–6301. [Google Scholar] [CrossRef] [Green Version]
- Tan, I.S.; Ramamurthi, K.S. Spore formation in Bacillus subtilis. Environ. Microbiol. Rep. 2014, 6, 212–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chastanet, A.; Vitkup, D.; Yuan, G.C.; Norman, T.M.; Liu, J.S.; Losick, R.M. Broadly heterogeneous activation of the master regulator for sporulation in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2010, 107, 8486–8491. [Google Scholar] [CrossRef] [Green Version]
- González-Pastor, J.E.; Hobbs, E.C.; Losick, R. Cannibalism by sporulating bacteria. Science 2003, 301, 510–513. [Google Scholar] [CrossRef] [PubMed]
- Losick, R.; Stragier, P. Crisscross regulation of cell-type-specific gene expression during development in B. subtilis. Nature 1992, 355, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Ogura, M.; Nii, S.; Hirooka, K. Dual Regulation of Bacillus subtilis kinB Gene Encoding a Sporulation Trigger by SinR through Transcription Repression and Positive Stringent Transcription Control. Front. Microbiol. 2017, 8, 2502. [Google Scholar] [CrossRef] [Green Version]
- Swarge, B.; Abhyankar, W.; Jonker, M.; Hoefsloot, H.; Kramer, G.; Setlow, P.; Brul, S.; de Koning, L.J. Integrative Analysis of Proteome and Transcriptome Dynamics during Bacillus subtilis Spore Revival. mSphere 2020, 5, e00463-20. [Google Scholar] [CrossRef]
- Zammuto, V.; Fuchs, F.M.; Fiebrandt, M.; Stapelmann, K.; Ulrich, N.J.; Maugeri, T.L.; Pukall, R.; Gugliandolo, C.; Moeller, R. Comparing Spore Resistance of Bacillus Strains Isolated from Hydrothermal Vents and Spacecraft Assembly Facilities to Environmental Stressors and Decontamination Treatments. Astrobiology 2018, 18, 1425–1434. [Google Scholar] [CrossRef]
- Tarrah, A.; Pakroo, S.; Carlot, M.; Nesto, C.; Cirillo, A.; Lombardi, A.; Corich, V.; Giacomini, A. Thermal resistance and high-performance microwave decontamination assessment of Bacillus endospores isolated from food-grade herbal extracts. PLoS ONE 2021, 16, e0261988. [Google Scholar] [CrossRef]
- Cortesão, M.; Fuchs, F.M.; Commichau, F.M.; Eichenberger, P.; Schuerger, A.C.; Nicholson, W.L.; Setlow, P.; Moeller, R. Bacillus subtilis Spore Resistance to Simulated Mars Surface Conditions. Front. Microbiol. 2019, 10, 333. [Google Scholar] [CrossRef] [Green Version]
- Setlow, P. Germination of spores of Bacillus species: What we know and do not know. J. Bacteriol 2014, 196, 1297–1305. [Google Scholar] [CrossRef] [Green Version]
- Koopman, N.; Remijas, L.; Seppen, J.; Setlow, P.; Brul, S. Mechanisms and Applications of Bacterial Sporulation and Germination in the Intestine. Int. J. Mol. Sci. 2022, 23, 3405. [Google Scholar] [CrossRef] [PubMed]
- Casula, G.; Cutting, S.M. Bacillus probiotics: Spore germination in the gastrointestinal tract. Appl. Environ. Microbiol. 2002, 68, 2344–2352. [Google Scholar] [CrossRef] [Green Version]
- Duc, L.H.; Hong, A.H.; Nguyen, Q.U.; Cutting, S.M. Intracellular fate and immunogenicity of B. subtilis spores. Vaccine 2004, 22, 1873–1885. [Google Scholar] [CrossRef] [PubMed]
- Bernardeau, M.; Lehtinen, M.J.; Forssten, S.D.; Nurminen, P. Importance of the gastrointestinal life cycle of Bacillus for probiotic functionality. J. Food Sci. Technol 2017, 54, 2570–2584. [Google Scholar] [CrossRef]
- Tam, N.K.; Uyen, N.Q.; Hong, H.A.; Duc, H.; Hoa, T.T.; Serra, C.R.; Henriques, A.O.; Cutting, S.M. The intestinal life cycle of Bacillus subtilis and close relatives. J. Bacteriol. 2006, 188, 2692–2700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Li, Z.; Gu, X.; Zhao, J.; Guo, T.; Kong, J. Probiotic Bacillus subtilis LF11 Protects Intestinal Epithelium Against Salmonella Infection. Front. Cell Infect. Microbiol. 2022, 12, 837886. [Google Scholar] [CrossRef] [PubMed]
- Shivangi, S.; Devi, P.B.; Ragul, K.; Shetty, P.H. Probiotic Potential of Bacillus Strains Isolated from an Acidic Fermented Food Idli. Probiotics Antimicrob. Proteins 2020, 12, 1502–1513. [Google Scholar] [CrossRef]
- Kotowicz, N.; Bhardwaj, R.K.; Ferreira, W.T.; Hong, H.A.; Olender, A.; Ramirez, J.; Cutting, S.M. Safety and probiotic evaluation of two Bacillus strains producing antioxidant compounds. Benef. Microbes 2019, 10, 759–771. [Google Scholar] [CrossRef]
- Driks, A. Bacillus subtilis spore coat. Microbiol. Mol. Biol. Rev. 1999, 63, 1–20. [Google Scholar] [CrossRef] [Green Version]
- McKenney, P.T.; Eichenberger, P. Dynamics of spore coat morphogenesis in Bacillus subtilis. Mol. Microbiol. 2012, 83, 245–260. [Google Scholar] [CrossRef] [Green Version]
- Imamura, D.; Kuwana, R.; Takamatsu, H.; Watabe, K. Proteins involved in formation of the outermost layer of Bacillus subtilis spores. J. Bacterial. 2011, 193, 4075–4080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cangiano, G.; Mazzone, A.; Baccigalupi, L.; Isticato, R.; Eichenberger, P.; De Felice, M.; Ricca, E. Direct and indirect control of late sporulation genes by GerR of Bacillus subtilis. J. Bacterial. 2010, 192, 3406–3413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krajčíková, D.; Bugárová, V.; Barák, I. Interactions of Bacillus subtilis Basement Spore Coat Layer Proteins. Microorganisms 2021, 9, 285. [Google Scholar] [CrossRef] [PubMed]
- Ursem, R.; Swarge, B.; Abhyankar, W.R.; Buncherd, H.; de Koning, L.J.; Setlow, P.; Brul, S.; Kramer, G. Identification of Native Cross-Links in Bacillus subtilis Spore Coat Proteins. J. Proteome Res. 2021, 20, 1809–1816. [Google Scholar] [CrossRef] [PubMed]
- Shuster, B.; Khemmani, M.; Abe, K.; Huang, X.; Nakaya, Y.; Maryn, N.; Buttar, S.; Gonzalez, A.N.; Driks, A.; Sato, T.; et al. Contributions of crust proteins to spore surface properties in Bacillus subtilis. Mol. Microbiol. 2019, 111, 825–843. [Google Scholar] [CrossRef]
- Freitas, C.; Plannic, J.; Isticato, R.; Pelosi, A.; Zilhão, R.; Serrano, M.; Baccigalupi, L.; Ricca, E.; Elsholz, A.K.W.; Losick, R.; et al. A protein phosphorylation module patterns the Bacillus subtilis spore outer coat. Mol. Microbiol. 2020, 114, 934–951. [Google Scholar] [CrossRef]
- Isticato, R.; Pelosi, A.; Zilhão, R.; Baccigalupi, L.; Henriques, A.O.; De Felice, M.; Ricca, E. CotC-CotU heterodimerization during assembly of the Bacillus subtilis spore coat. J. Bacteriol. 2008, 190, 1267–1275. [Google Scholar] [CrossRef] [Green Version]
- Isticato, R.; Sirec, T.; Vecchione, S.; Crispino, A.; Saggese, A.; Baccigalupi, L.; Notomista, E.; Driks, A.; Ricca, E. The Direct Interaction between Two Morphogenetic Proteins Is Essential for Spore Coat Formation in Bacillus subtilis. PLoS ONE 2015, 10, e0141040. [Google Scholar] [CrossRef] [Green Version]
- Isticato, R.; Pelosi, A.; De Felice, M.; Ricca, E. CotE binds to CotC and CotU and mediates their interaction during spore coat formation in Bacillus subtilis. J. Bacteriol. 2010, 192, 949–954. [Google Scholar] [CrossRef] [Green Version]
- Isticato, R.; Sirec, T.; Giglio, R.; Baccigalupi, L.; Rusciano, G.; Pesce, G.; Zito, G.; Sasso, A.; De Felice, M.; Ricca, E. Flexibility of the programme of spore coat formation in Bacillus subtilis: Bypass of CotE requirement by over-production of CotH. PLoS ONE 2013, 8, e74949. [Google Scholar] [CrossRef]
- Cangiano, G.; Sirec, T.; Panarella, C.; Isticato, R.; Baccigalupi, L.; De Felice, M.; Ricca, E. The sps gene products affect germination, hydrophobicity and protein adsorption of Bacillus subtilis spores. Appl. Environ. Microbiol. 2014, 80, 7293–7302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Wan, Q.; Krajcikova, D.; Tang, J.; Tzokov, S.B.; Barak, I.; Bullough, P.A. Diverse supramolecular structures formed by self-assembling proteins of the Bacillus subtilis spore coat. Mol. Microbiol. 2015, 97, 347–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesce, G.; Rusciano, G.; Sasso, A.; Isticato, R.; Sirec, T.; Ricca, E. Surface charge and hydrodynamic coefficient measurements of Bacillus subtilis spore by optical tweezers. Colloids Surf. B 2014, 116, 568–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, T.; Krzewinski, F.; Yamakawa, N.; Lemy, C.; Hamiot, A.; Brunet, L.; Lacoste, A.S.; Knirel, Y.; Guerardel, Y.; Faille, C. The sps Genes Encode an Original Legionaminic Acid Pathway Required for Crust Assembly in Bacillus subtilis. mBio 2020, 11, e01153-20. [Google Scholar] [CrossRef] [PubMed]
- Bressuire-Isoard, C.; Bornard, I.; Henriques, A.O.; Carlin, F.; Broussolle, V. Sporulation Temperature Reveals a Requirement for CotE in the Assembly of both the Coat and Exosporium Layers of Bacillus cereus Spores. Appl. Environ. Microbiol. 2015, 82, 232–243. [Google Scholar] [CrossRef] [Green Version]
- Stewart, G.C. The Exosporium Layer of Bacterial Spores: A Connection to the Environment and the Infected Host. MMBR Rev. 2015, 79, 437–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vittoria, M.; Saggese, A.; Di Gregorio Barletta, G.; Castaldi, S.; Isticato, R.; Baccigalupi, L.; Ricca, E. Sporulation efficiency and spore quality in a human intestinal isolate of Bacillus cereus. Res. Microbiol. 2023, 104030. [Google Scholar] [CrossRef]
- Isticato, R.; Ricca, E. Spore Surface Display. Microbiol Spectrum 2014, 2, 351–366. [Google Scholar] [CrossRef]
- Lin, P.; Yuan, H.; Du, J.; Liu, K.; Liu, H.; Wang, T. Progress in research and application development of surface display technology using Bacillus subtilis spores. Appl. Microbiol. Biotechnol. 2020, 104, 2319–2331. [Google Scholar] [CrossRef]
- Sirec, T.; Cangiano, G.; Baccigalupi, L.; Ricca, E.; Isticato, R. The spore surface of intestinal isolates of Bacillus subtilis. FEMS Microbiol. Lett. 2014, 358, 194–201. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Wu, Z.; Sun, H.; Zhao, L.; Shang, M.; Shi, M.; Jiang, H.; Lin, Z.; Zhou, X.; Li, X.; et al. The storage stability of Bacillus subtilis spore displaying cysteine protease of Clonorchis sinensis and its effect on improving the gut microbiota of mice. Appl. Microbiol. Biotechnol. 2021, 105, 2513–2526. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Kim, W.S.; Paik, H.D. Bacillus strains as human probiotics: Characterization, safety, microbiome, and probiotic carrier. Food Sci. Biotechnol. 2019, 28, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Le, H.D.; Hong, H.A.; Atkins, H.S.; Flick-Smith, H.C.; Durrani, Z.; Rijpkema, S.; Titball, R.W.; Cutting, S.M. Immunization against anthrax using Bacillus subtilis spores expressing the anthrax protective antigen. Vaccine 2007, 25, 346–355. [Google Scholar] [CrossRef]
- Scheiblhofer, S.; Laimer, J.; Machado, Y.; Weiss, R.; Thalhamer, J. Influence of protein fold stability on immunogenicity and its implications for vaccine design. Expert Rev. Vaccines 2017, 16, 479–489. [Google Scholar] [CrossRef] [Green Version]
- Aung, A.; Cui, A.; Maiorino, L.; Amini, A.P.; Gregory, J.R.; Bukenya, M.; Zhang, Y.; Lee, H.; Cottrell, C.A.; Morgan, D.M.; et al. Low protease activity in B cell follicles promotes retention of intact antigens after immunization. Science 2023, 379, eabn8934. [Google Scholar] [CrossRef]
- Turk, B. Targeting proteases: Successes, failures and future prospects. Nat. Rev. Drug Discov. 2006, 5, 785–799. [Google Scholar] [CrossRef]
- Negri, A.; Potocki, W.; Iwanicki, A.; Obuchowski, M.; Hinc, K. Expression and display of Clostridium difficile protein FliD on the surface of Bacillus subtilis spores. J. Med. Microbiol. 2013, 62, 1379–1385. [Google Scholar] [CrossRef]
- Tavares, M.B.; Silva, B.M.; Cavalcante, R.C.; Souza, R.D.; Luiz, W.B.; Paccez, J.D.; Crowley, P.J.; Brady, L.J.; Ferreira, L.C.; Ferreira, R.C. Induction of neutralizing antibodies in mice immunized with an amino-terminal polypeptide of Streptococcus mutans P1 protein produced by a recombinant Bacillus subtilis strain. FEMS Immunol. Med. Microbiol. 2010, 59, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Saggese, A.; Di Gregorio Barletta, G.; Vittoria, M.; Donadio, G.; Isticato, R.; Baccigalupi, L.; Ricca, E. CotG Mediates Spore Surface Permeability in Bacillus subtilis. mBio 2022, 13, e0276022. [Google Scholar] [CrossRef]
- Zilhão, R.; Serrano, M.; Isticato, R.; Ricca, E.; Moran, C.P., Jr.; Henriques, A.O. Interactions among CotB, CotG, and CotH during assembly of the Bacillus subtilis spore coat. J. Bacteriol. 2004, 186, 1110–1119. [Google Scholar] [CrossRef] [Green Version]
- Giglio, R.; Fani, R.; Isticato, R.; De Felice, M.; Ricca, E.; Baccigalupi, L. Organization and evolution of the cotG and cotH genes of Bacillus subtilis. J. Bacteriol. 2011, 193, 6664–6673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciabattini, A.; Parigi, R.; Isticato, R.; Oggioni, M.R.; Pozzi, G. Oral priming of mice by recombinant spores of Bacillus subtilis. Vaccine 2004, 22, 4139–4143. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Kim, J.A.; Kim, C.H.; Choi, S.K.; Pan, J.G. Bacillus subtilis spore vaccines displaying protective antigens induce functional antibodies and protective potency. BMC Vet. Res. 2020, 16, 259. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.H.; Hong, H.A.; Clark, G.C.; Titball, R.W.; Cutting, S.M. Recombinant Bacillus subtilis expressing the Clostridium perfringens alpha toxoid is a candidate orally delivered vaccine against necrotic enteritis. Infect. Immun. 2008, 76, 5257–5265. [Google Scholar] [CrossRef] [Green Version]
- Hinc, K.; Isticato, R.; Dembek, M.; Karczewska, J.; Iwanicki, A.; Peszyńska-Sularz, G.; De Felice, M.; Obuchowski, M.; Ricca, E. Expression and display of UreA of Helicobacter acinonychis on the surface of Bacillus subtilis spores. Microb. Cell Factories 2010, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.A.; Hitri, K.; Hosseini, S.; Kotowicz, N.; Bryan, D.; Mawas, F.; Wilkinson, A.J.; van Broekhoven, A.; Kearsey, J.; Cutting, S.M. Mucosal antibodies to the C terminus of toxin A prevent colonization of Clostridium difficile. Infect. Immun. 2017, 85, e01060-16. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.T.; Pham, C.K.; Pham, H.T.; Pham, H.L.; Nguyen, A.H.; Dang, L.T.; Huynh, H.A.; Cutting, S.M.; Phan, T.N. Bacillus subtilis spores expressing the VP28 antigen: A potential oral treatment to Litopenaeus vannamei against white spot syndrome. FEMS Microbiol. Lett. 2014, 358, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Mauriello, E.M.; Duc, H.; Isticato, R.; Cangiano, G.; Hong, H.A.; De Felice, M.; Ricca, E.; Cutting, S.M. Display of heterologous antigens on the Bacillus subtilis spore coat using CotC as a fusion partner. Vaccine 2004, 22, 1177–1187. [Google Scholar] [CrossRef]
- Isticato, R.; Di Mase, D.S.; Mauriello, E.M.; De Felice, M.; Ricca, E. Amino terminal fusion of heterologous proteins to CotC increases display efficiencies in the Bacillus subtilis spore system. Bio Tech. 2007, 42, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Xia, H.; Hu, X.; Huang, Y.; Li, Y.; Li, L.; Ma, C.; Chen, X.; Hu, F.; Xu, J.; et al. Oral administration of a Bacillus subtilis spore-based vaccine expressing Clonorchis sinensis tegumental protein 22.3 kDa confers protection against Clonorchis sinensis. Vaccine 2008, 26, 1817–1825. [Google Scholar] [CrossRef]
- Tang, Z.; Shang, M.; Chen, T.; Ren, P.; Sun, H.; Qu, H.; Lin, Z.; Zhou, L.; Yu, J.; Jiang, H.; et al. The immunological characteristics and probiotic function of recombinant Bacillus subtilis spore expressing Clonorchis sinensis cysteine protease. Parasites Vectors 2016, 9, 648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, H.; Xu, Y.; Sun, H.; Lin, J.; Yu, J.; Tang, Z.; Shen, J.; Liang, C.; Li, S.; Chen, W.; et al. Systemic and local mucosal immune responses induced by orally delivered Bacillus subtilis spore expressing leucine aminopeptidase 2 of Clonorchis sinensis. Parasitol. Res. 2014, 113, 3095–3103. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chen, T.; Xie, Z.; Liang, P.; Qu, H.; Shang, M.; Mao, Q.; Ning, D.; Tang, Z.; Shi, M.; et al. Oral delivery of Bacillus subtilis spore expressing enolase of Clonorchis sinensis in rat model: Induce systemic and local mucosal immune responses and has no side effect on liver function. Parasitol. Res. 2015, 114, 2499–2505. [Google Scholar] [CrossRef]
- Sun, H.; Lin, Z.; Zhao, L.; Chen, T.; Shang, M.; Jiang, H.; Tang, Z.; Zhou, X.; Shi, M.; Zhou, L.; et al. Bacillus subtilis spore with surface display of paramyosin from Clonorchis sinensis potentializes a promising oral vaccine candidate. Parasites Vectors 2018, 11, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.; Sun, H.; Ma, Y.; Zhou, X.; Jiang, H.; Wang, X.; Song, J.; Tang, Z.; Bian, Q.; Zhang, Z.; et al. Evaluation of immune response to Bacillus subtilis spores expressing Clonorchis sinensis serpin3. Parasitology 2020, 147, 1080–1087. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, C.S.; Kim, B.G. Spore-displayed streptavidin: A live diagnostic tool in biotechnology. Biochem. Biophys. Res. Commun. 2005, 331, 210–214. [Google Scholar] [CrossRef]
- Kim, J.; Schumann, W. Display of proteins on Bacillus subtilis endospores. Cell Mol. Life Sci. 2009, 66, 3127–3313. [Google Scholar] [CrossRef]
- Mou, C.; Zhu, L.; Xing, X.; Lin, J.; Yang, Q. Immune responses induced by recombinant Bacillus subtilis expressing the spike protein of transmissible gastroenteritis virus in pigs. Antivir. Res. 2016, 131, 74–84. [Google Scholar] [CrossRef]
- Iwanicki, A.; Piatek, I.; Stasilojć, M.; Grela, A.; Lega, T.; Obuchowski, M.; Hinc, K. A system of vectors for Bacillus subtilis spore surface display. Microb. Cell Factories 2014, 13, 30. [Google Scholar] [CrossRef] [Green Version]
- Sibley, L.; Reljic, R.; Radford, D.S.; Huang, J.M.; Hong, H.A.; Cranenburgh, R.M.; Cutting, S.M. Recombinant Bacillus subtilis spores expressing MPT64 evaluated as a vaccine against tuberculosis in the murine model. FEMS Microbiol. Lett. 2014, 358, 170–179. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Miao, Y.; Guo, Y.; Qiu, H.; Sun, S.; Kou, Z.; Yu, H.; Li, J.; Chen, Y.; Jiang, S.; et al. Development of a heat-stable and orally delivered recombinant M2e-expressing B. subtilis spore-based influenza vaccine. Hum. Vaccines Immunother. 2014, 10, 3649–3658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinc, K.; Stasiłojć, M.; Piątek, I.; Peszyńska-Sularz, G.; Isticato, R.; Ricca, E.; Obuchowski, M.; Iwanicki, A. Mucosal adjuvant activity of IL-2 presenting spores of Bacillus subtilis in a murine model of Helicobacter pylori vaccination. PLoS ONE 2014, 9, e95187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, X.; Liu, M.; Pan, K.; Yang, J. Surface display of OmpC of Salmonella serovar Pullorum on Bacillus subtilis spores. PLoS ONE 2018, 13, e0191627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Gong, S.; Li, X.M.; Yang, Y.; Guan, R.; Zhou, S.; Yao, S.; Xie, Y.; Ou, Z.; Zhao, J.; et al. Expression of Helicobacter pylori urease B on the surface of Bacillus subtilis spores. J. Med. Microbiol. 2015, 64, 104–105. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Hu, X.; Wu, Z.; Xiong, S.; Zhou, Z.; Wang, X.; Xu, J.; Lu, F.; Yu, X. Immunogenicity of self-adjuvanticity oral vaccine candidate based on use of Bacillus subtilis spore displaying Schistosoma japonicum 26 KDa GST protein. Parasitol. Res. 2009, 105, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Bian, Q.; Zeng, W.; Ren, P.; Sun, H.; Lin, Z.; Tang, Z.; Zhou, X.; Wang, Q.; Wang, Y.; et al. Oral delivery of Bacillus subtilis spores expressing grass carp reovirus VP4 protein produces protection against grass carp reovirus infection. Fish Shellfish. Immunol. 2018, 84, 768–780. [Google Scholar] [CrossRef]
- Valdez, A.; Yepiz-Plascencia, G.; Ricca, E.; Olmos, J. First Litopenaeus vannamei WSSV 100% oral vaccination protection using CotC::Vp26 fusion protein displayed on Bacillus subtilis spores surface. J. Appl. Microbiol. 2014, 117, 347–357. [Google Scholar] [CrossRef]
- Li, G.; Tang, Q.; Chen, H.; Yao, Q.; Ning, D.; Chen, K. Display of Bombyx mori nucleopolyhedrovirus GP64 on the Bacillus subtilis spore coat. Curr. Microbiol. 2011, 62, 1368–1373. [Google Scholar] [CrossRef]
- Hinc, K.; Iwanicki, A.; Obuchowski, M. New stable anchor protein and peptide linker suitable for successful spore surface display in B. subtilis. Microb. Cell Factories 2013, 12, 22. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Li, G.; Zhang, C.; Xing, X.H. A study on the effects of linker flexibility on acid phosphatase PhoC-GFP fusion protein using a novel linker library. Enzym. Microb. Technol. 2016, 83, 1–6. [Google Scholar] [CrossRef]
- Guérout-Fleury, A.M.; Frandsen, N.; Stragier, P. Plasmids for ectopic integration in Bacillus subtilis. Gene 1996, 180, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Middleton, R.; Hofmeister, A. New shuttle vectors for ectopic insertion of genes into Bacillus subtilis. Plasmid 2004, 51, 238–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stasiłojć, M.; Hinc, K.; Peszyńska-Sularz, G.; Obuchowski, M.; Iwanicki, A. Recombinant Bacillus subtilis Spores Elicit Th1/Th17-Polarized Immune Response in a Murine Model of Helicobacter pylori Vaccination. Mol. Biotech. 2015, 57, 685–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.M.; Hong, H.A.; Van Tong, H.; Hoang, T.H.; Brisson, A.; Cutting, S.M. Mucosal delivery of antigens using adsorption to bacterial spores. Vaccine 2010, 28, 1021–1030. [Google Scholar] [CrossRef]
- Bonavita, R.; Isticato, R.; Maurano, F.; Ricca, E.; Rossi, M. Mucosal immunity induced by gliadin-presenting spores of Bacillus subtilis in HLA-DQ8-transgenic mice. Immunol. Lett. 2015, 165, 84–89. [Google Scholar] [CrossRef]
- Isticato, R.; Sirec, T.; Treppiccione, L.; Maurano, F.; De Felice, M.; Rossi, M.; Ricca, E. Non-recombinant display of the B subunit of the heat labile toxin of Escherichia coli on wild type and mutant spores of Bacillus subtilis. Microb. Cell Factories 2013, 12, 98. [Google Scholar] [CrossRef] [Green Version]
- Maia, A.R.; Reyes-Ramírez, R.; Pizarro-Guajardo, M.; Saggese, A.; Castro-Córdova, P.; Isticato, R.; Ricca, E.; Paredes-Sabja, D.; Baccigalupi, L. Induction of a Specific Humoral Immune Response by Nasal Delivery of Bcla2ctd of Clostridioides difficile. Int. J. Mol. Sci. 2020, 21, 1277. [Google Scholar] [CrossRef] [Green Version]
- Santos, F.D.S.; Mazzoli, A.; Maia, A.R.; Saggese, A.; Isticato, R.; Leite, F.; Iossa, S.; Ricca, E.; Baccigalupi, L. A probiotic treatment increases the immune response induced by the nasal delivery of spore-adsorbed TTFC. Microb. Cell Factories 2020, 19, 42. [Google Scholar] [CrossRef] [Green Version]
- de Almeida, M.E.M.; Alves, K.C.S.; de Vasconcelos, M.G.S.; Pinto, T.S.; Glória, J.C.; Chaves, Y.O.; Neves, W.L.L.; Tarragô, A.M.; de Souza Neto, J.N.; Astolfi-Filho, S.; et al. Bacillus subtilis spores as delivery system for nasal Plasmodium falciparum circumsporozoite surface protein immunization in a murine model. Sci. Rep. 2022, 12, 1531. [Google Scholar] [CrossRef]
- Ricca, E.; Baccigalupi, L.; Cangiano, G.; De Felice, M.; Isticato, R. Mucosal vaccine delivery by non-recombinant spores of Bacillus subtilis. Microb. Cell Factories 2014, 13, 115. [Google Scholar] [CrossRef] [Green Version]
- Lanzilli, M.; Donadio, G.; Fusco, F.A.; Sarcinelli, C.; Limauro, D.; Ricca, E.; Isticato, R. Display of the peroxiredoxin Bcp1 of Sulfolobus solfataricus on probiotic spores of Bacillus megaterium. New Biotechnol. 2018, 46, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Donadio, G.; Lanzilli, M.; Sirec, T.; Ricca, E.; Isticato, R. Localization of a red fluorescence protein adsorbed on wild type and mutant spores of Bacillus subtilis. Microb. Cell Factories 2016, 15, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isticato, R.; Lanzilli, M.; Petrillo, C.; Donadio, G.; Baccigalupi, L.; Ricca, E. Bacillus subtilis builds structurally and functionally different spores in response to the temperature of growth. Environ. Microbiol. 2020, 22, 170–182. [Google Scholar] [CrossRef]
- Petrillo, C.; Castaldi, S.; Lanzilli, M.; Saggese, A.; Donadio, G.; Baccigalupi, L.; Ricca, E.; Isticato, R. The temperature of growth and sporulation modulates the efficiency of spore-display in Bacillus subtilis. Microb. Cell Factories 2020, 19, 185. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isticato, R. Bacterial Spore-Based Delivery System: 20 Years of a Versatile Approach for Innovative Vaccines. Biomolecules 2023, 13, 947. https://doi.org/10.3390/biom13060947
Isticato R. Bacterial Spore-Based Delivery System: 20 Years of a Versatile Approach for Innovative Vaccines. Biomolecules. 2023; 13(6):947. https://doi.org/10.3390/biom13060947
Chicago/Turabian StyleIsticato, Rachele. 2023. "Bacterial Spore-Based Delivery System: 20 Years of a Versatile Approach for Innovative Vaccines" Biomolecules 13, no. 6: 947. https://doi.org/10.3390/biom13060947
APA StyleIsticato, R. (2023). Bacterial Spore-Based Delivery System: 20 Years of a Versatile Approach for Innovative Vaccines. Biomolecules, 13(6), 947. https://doi.org/10.3390/biom13060947