
The Effects of Natural Epigenetic Therapies in 3D Ovarian Cancer and Patient-Derived Tumor Explants: New Avenues in Regulating the Cancer Secretome

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Therapeutic Agents
2.2. Tissue Culture
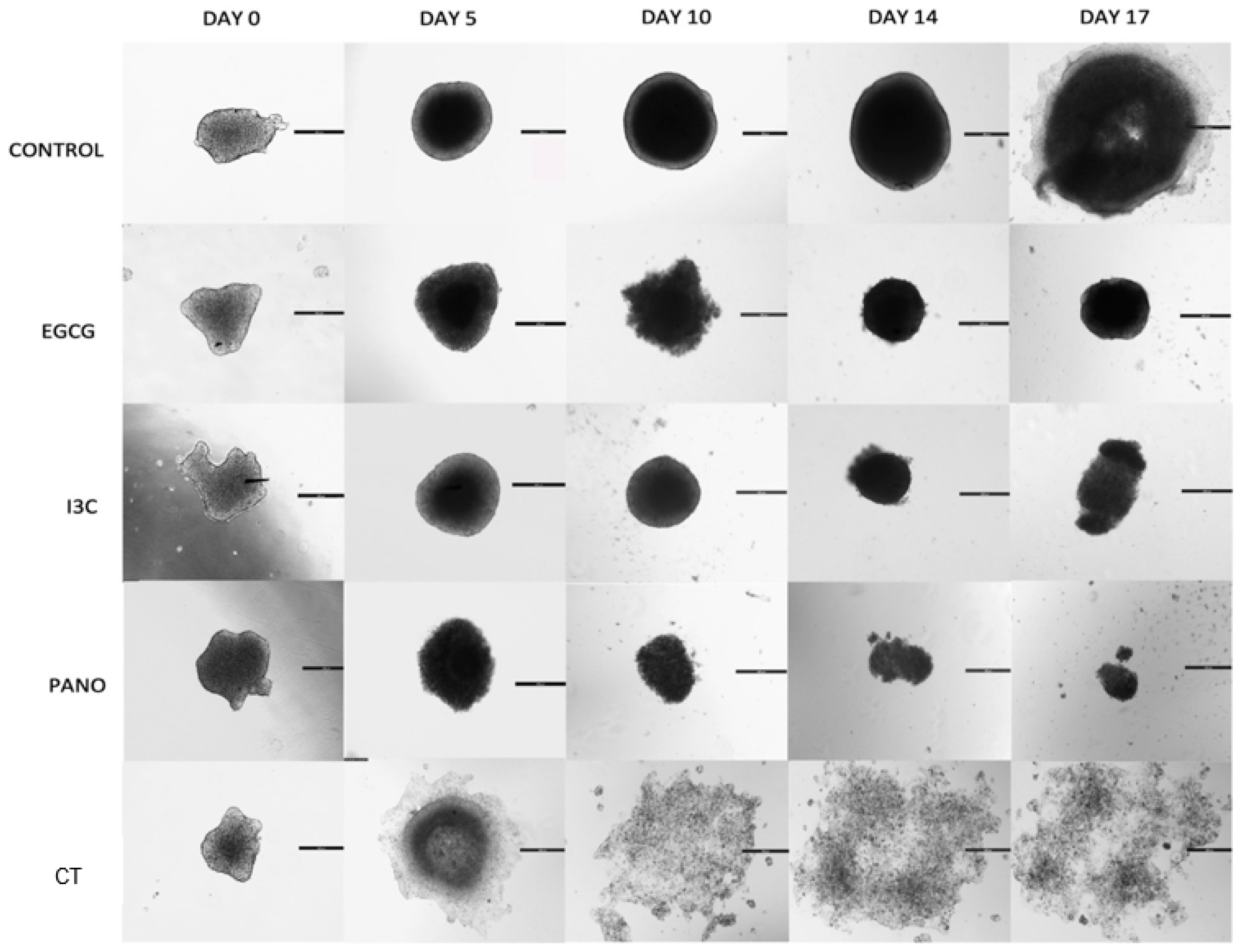
2.2.1. Establishing 3D Tissue Cultures
2.2.2. Measurement of Spheroid and Organoid Growth in 3D Culture
2.3. Cellular Assays
2.3.1. Cell Viability, Apoptosis, and Necrosis
2.3.2. Proliferation and Regrowth Ability of Cells
2.3.3. Migration and Invasion
2.3.4. Observation of Colony Formation Following Treatment in 3D Culture
2.4. Tissue Assays
2.4.1. Cancer Tissue Preparation and Treatments
2.4.2. Elisa
2.4.3. Protein Array
2.5. Statistical Analysis
3. Results
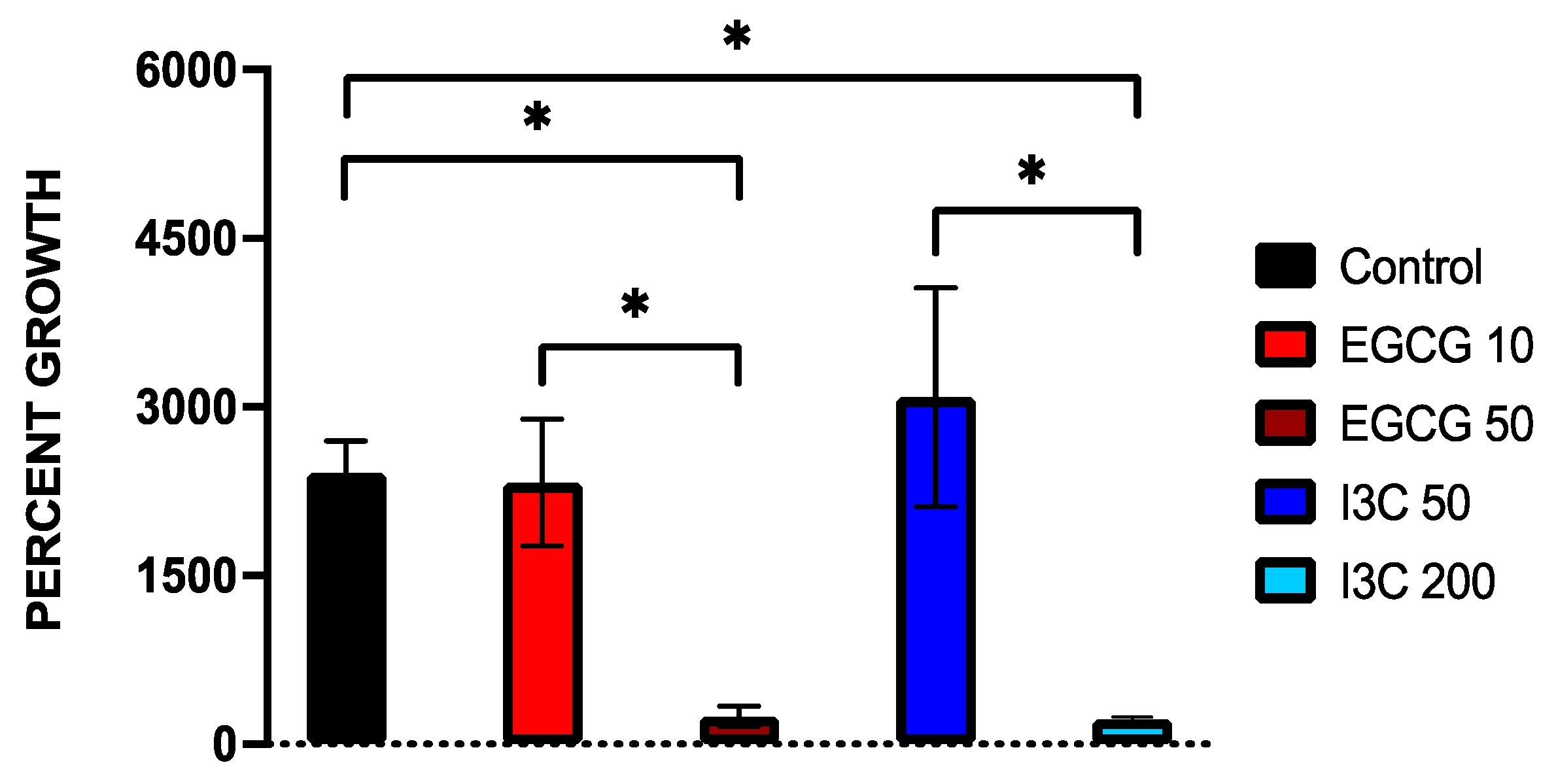
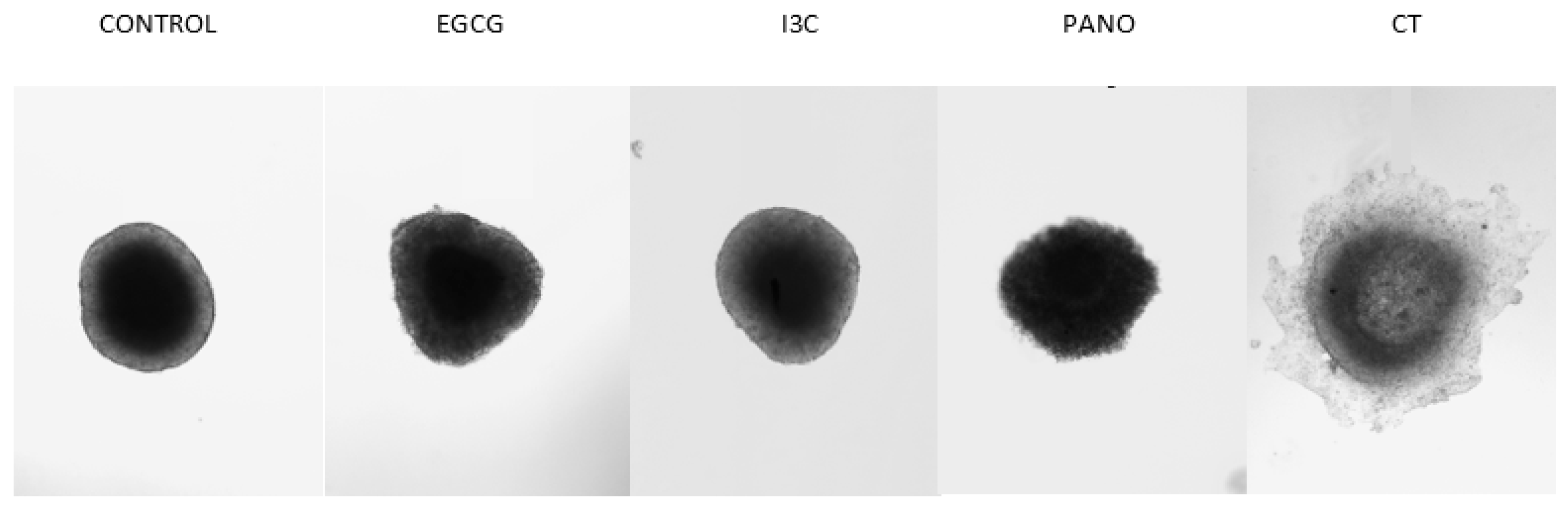
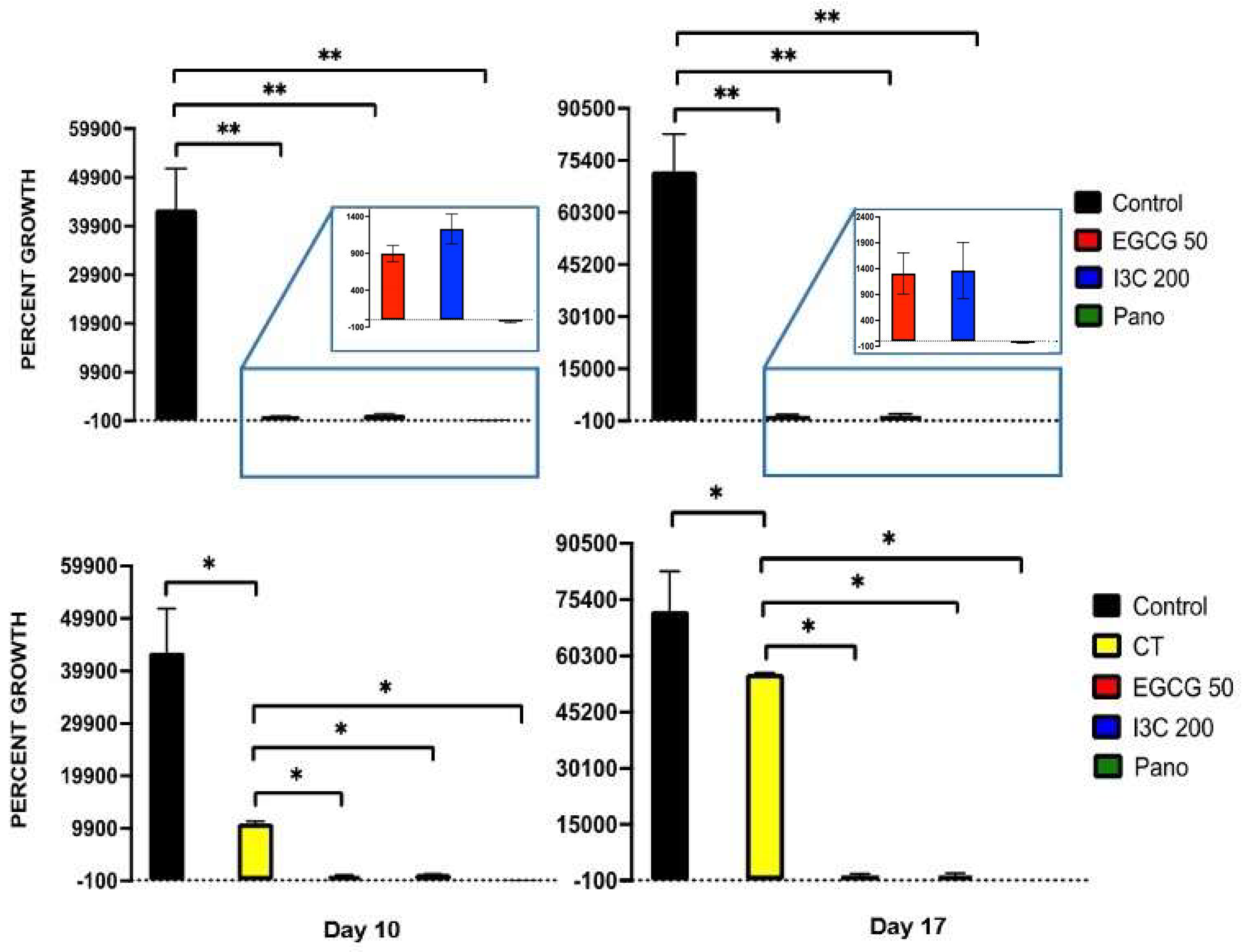
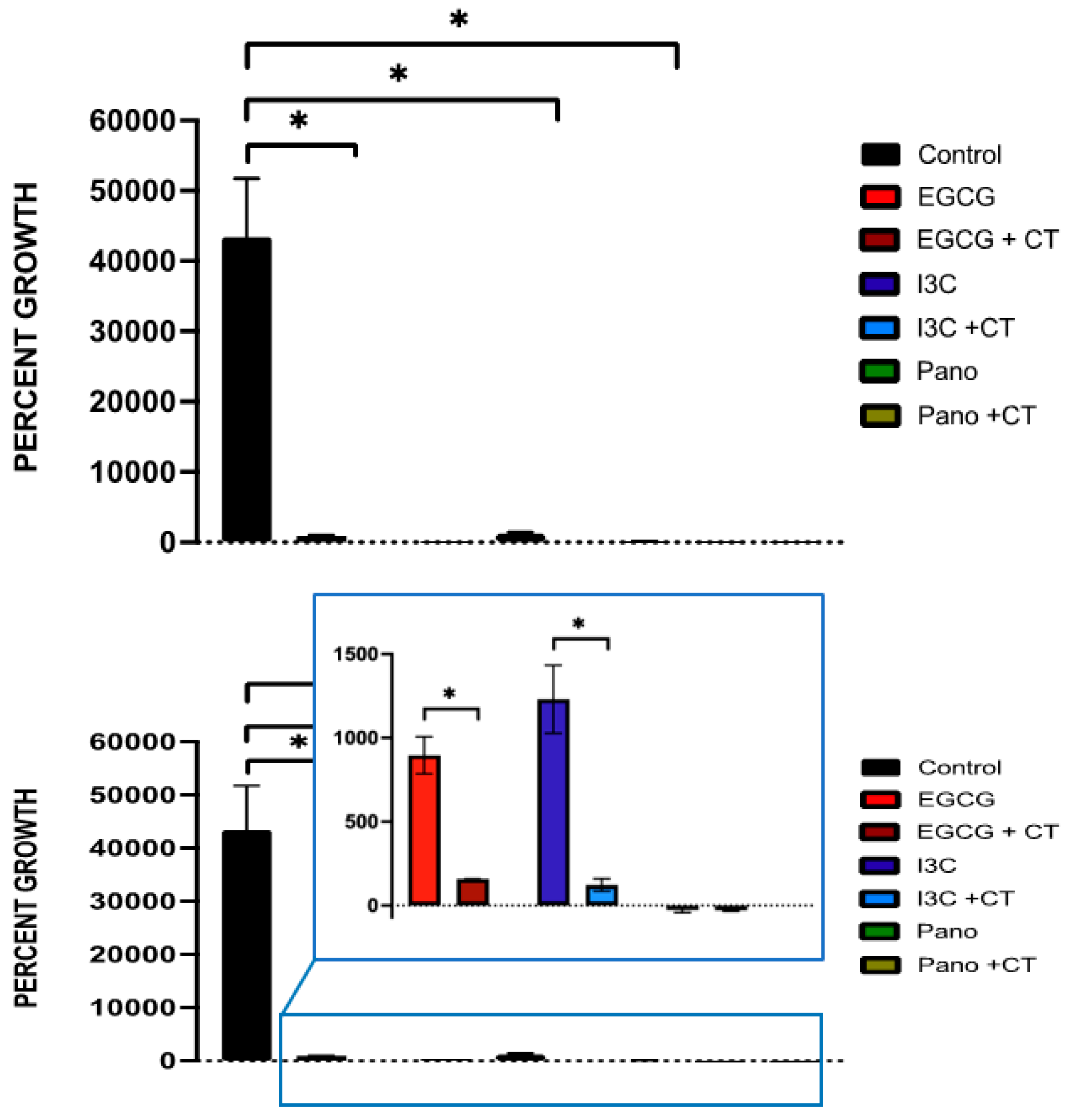
3.1. Dose-Dependent Effect of Natural Epigenetic Therapies on 3D Ovarian Cancer Growth
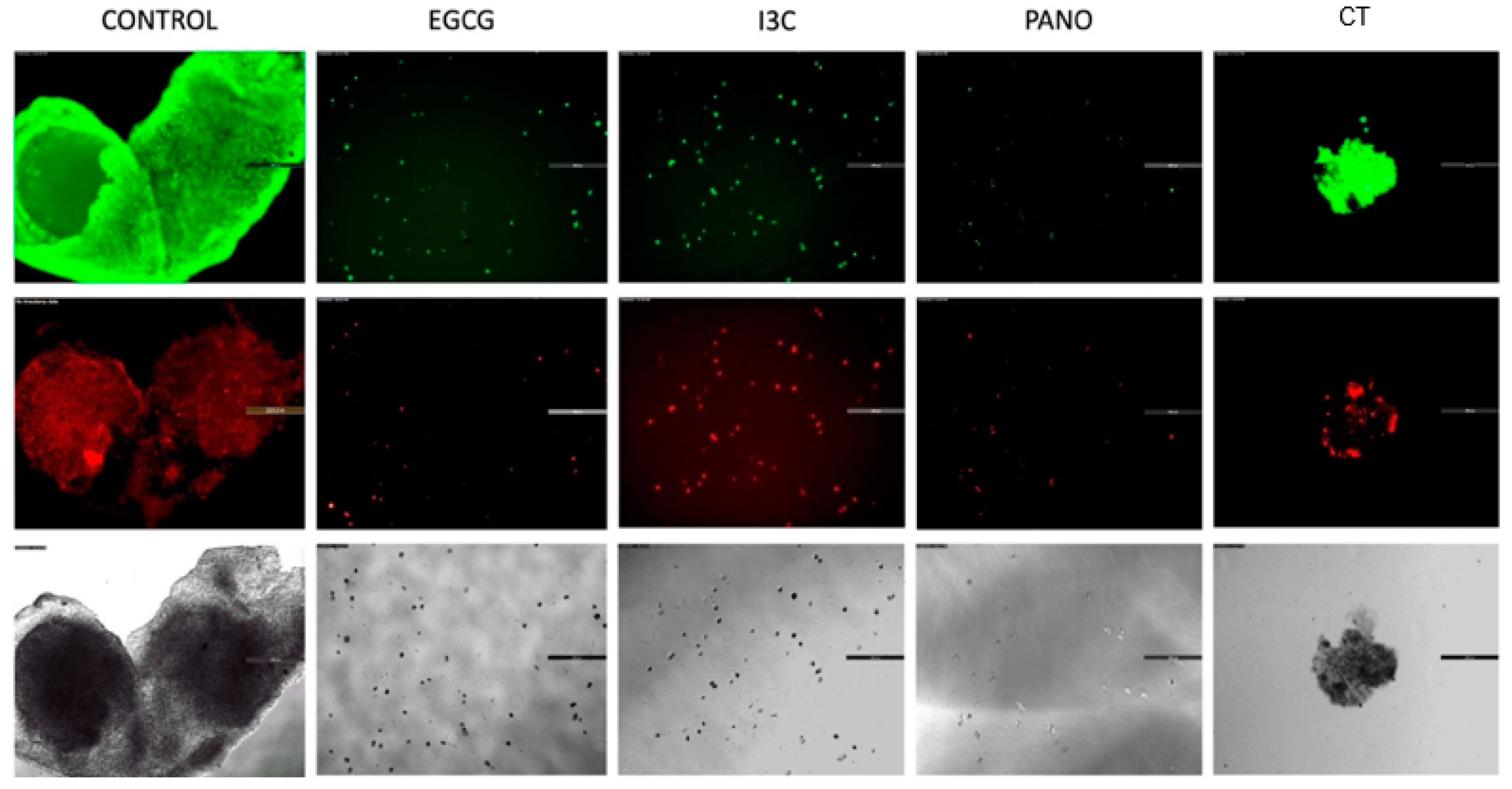
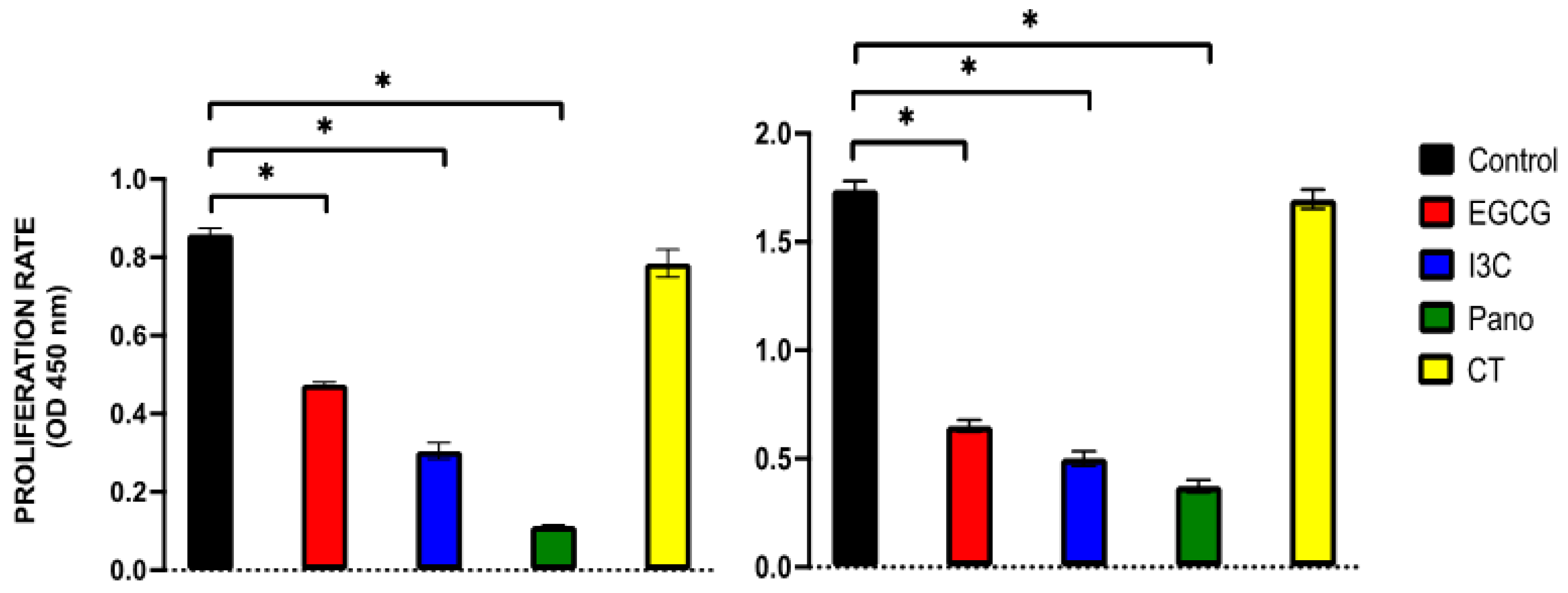
3.2. Apoptosis and Necrosis Assays in Pre-Treated Spheroids
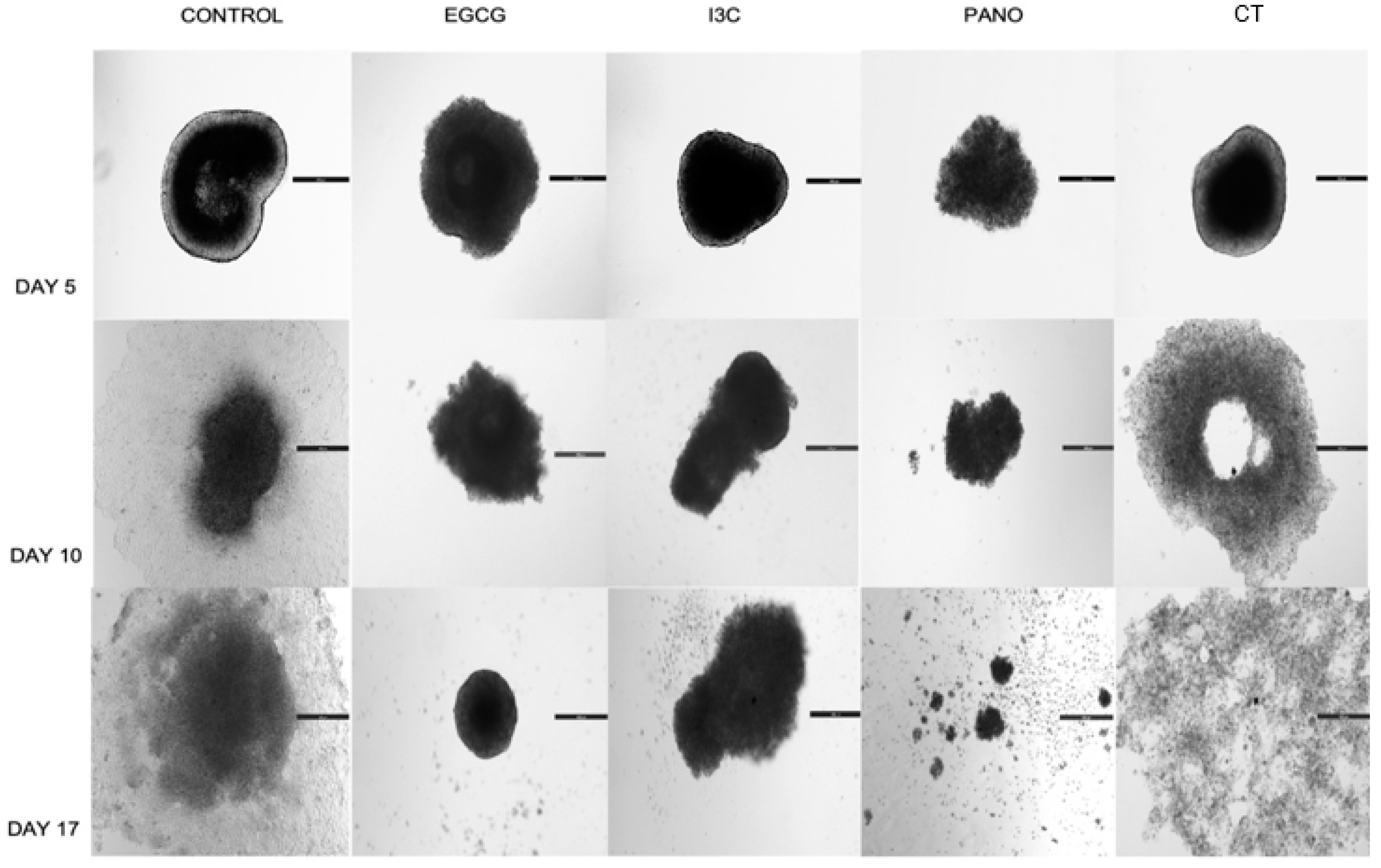
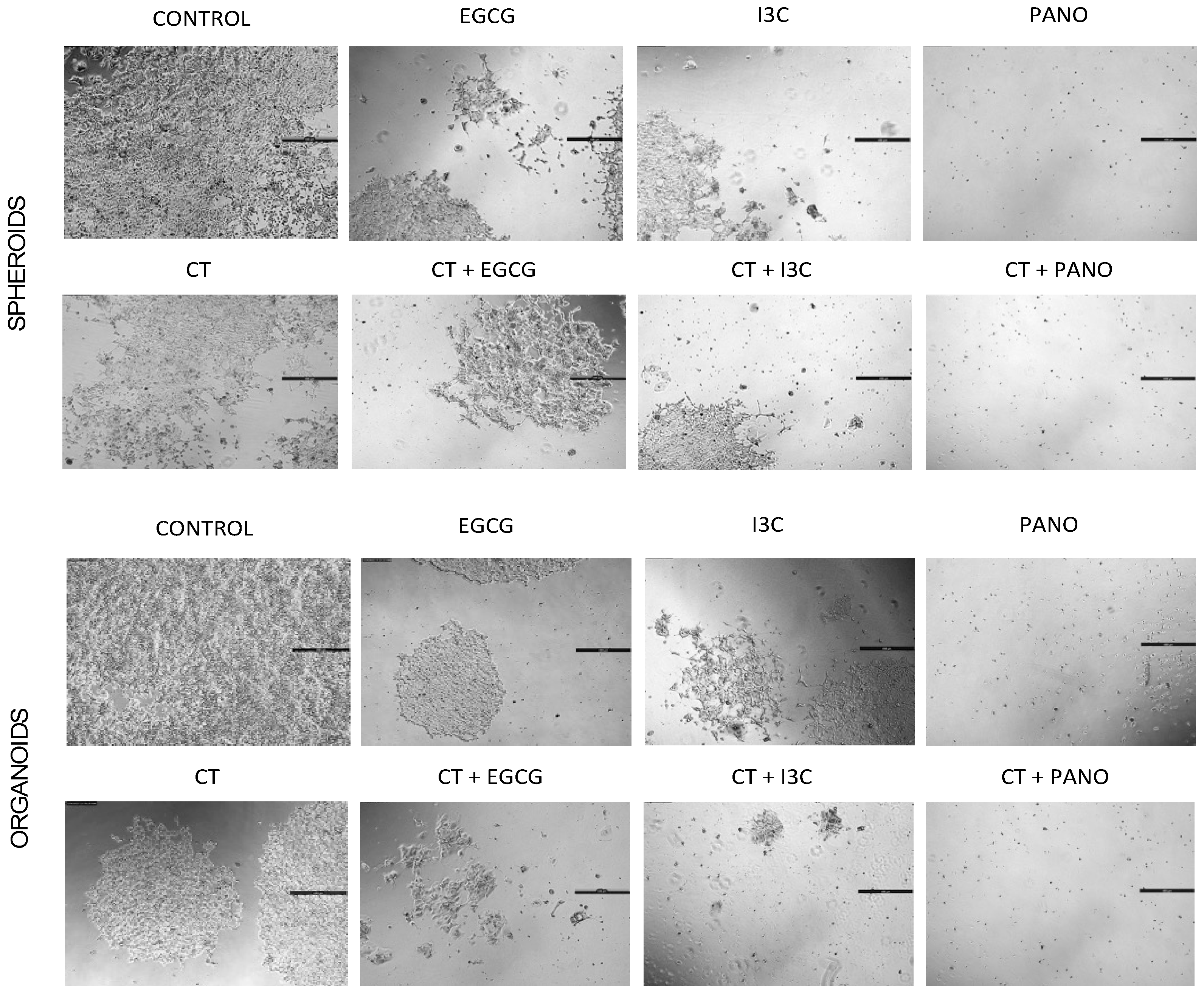
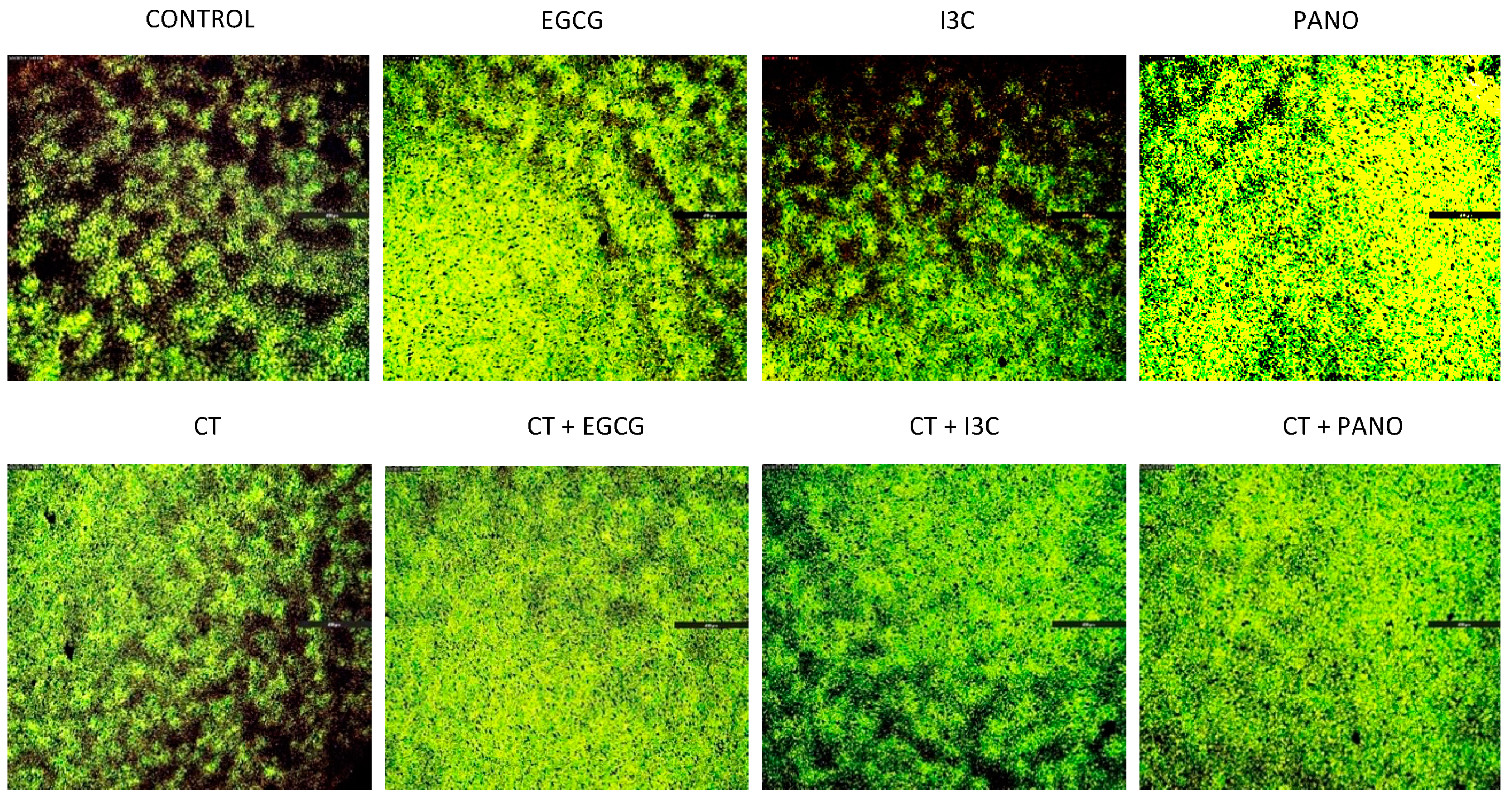
3.3. Lateral Spread of Microdisease
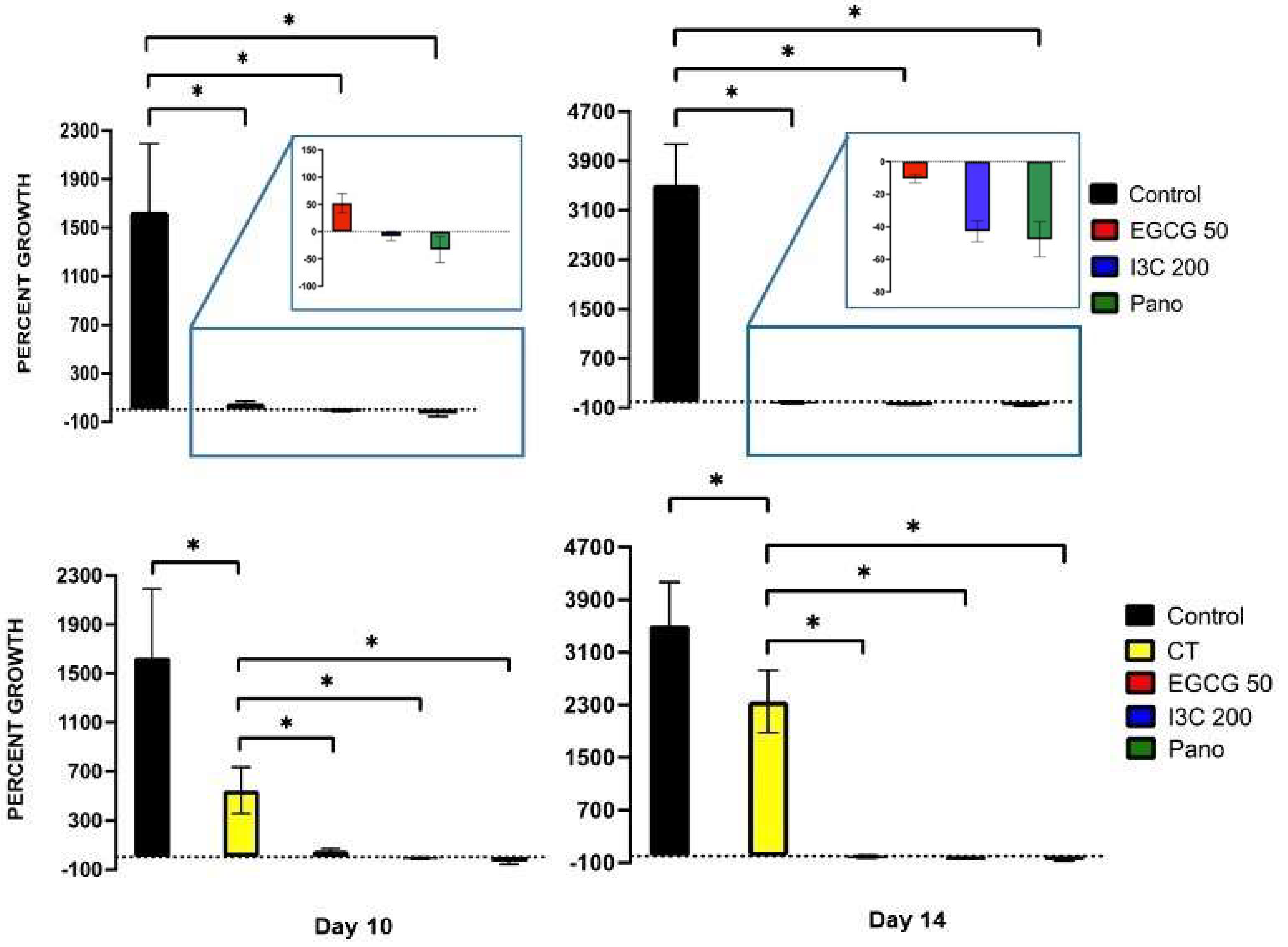
3.4. Can Epigenetic Therapies Affect Standard Chemotherapy Efficiency in the Treatment of 3D Ovarian Cancer?
3.5. Does the Tumorigenic Microenvironment Affect the Efficiency of Epigenetic Therapies?
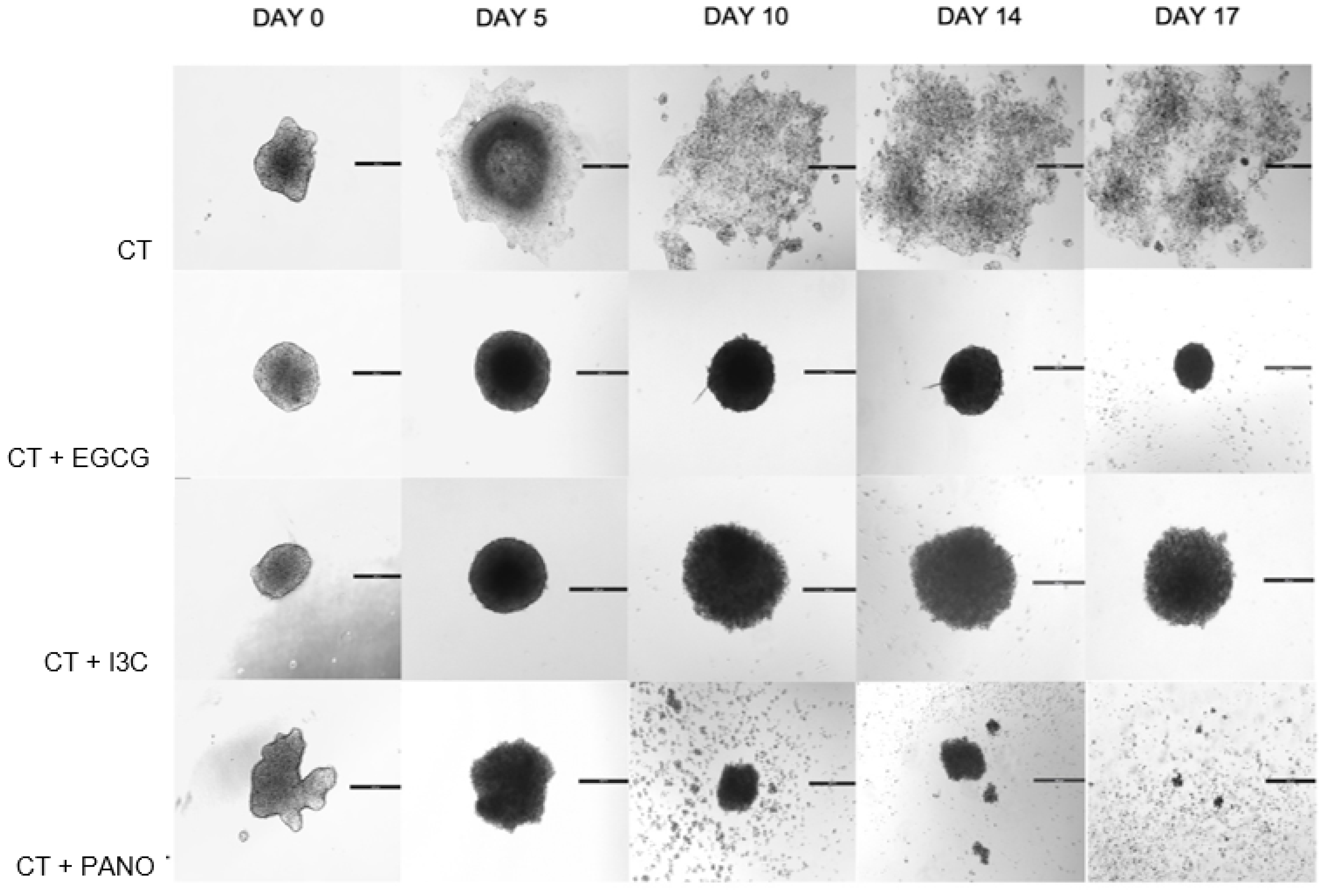
3.6. Do Epigenetic Therapies Suppress Post-Treatment Regrowth, Metastasis, and Migration of 3D Derived Ovarian Cancer Cells?
3.6.1. Post-Treatment Ability of 3D-Derived Spheroid and Organoid Cells to Regrow and Proliferate
3.6.2. Post-Treatment Invasion and Migration Ability of 3D-Derived Spheroid and Organoid Cells Following Epigenetic Treatments
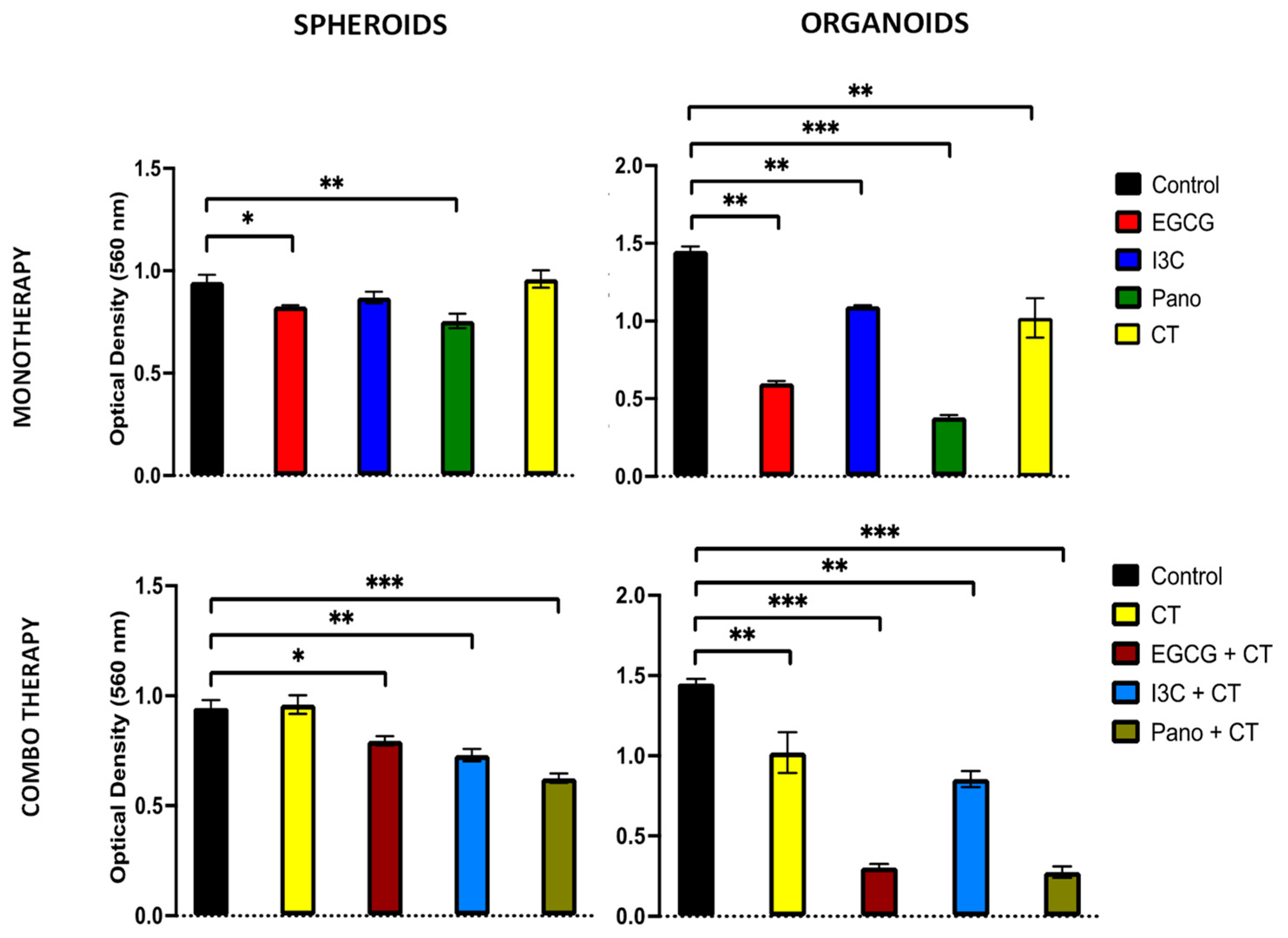
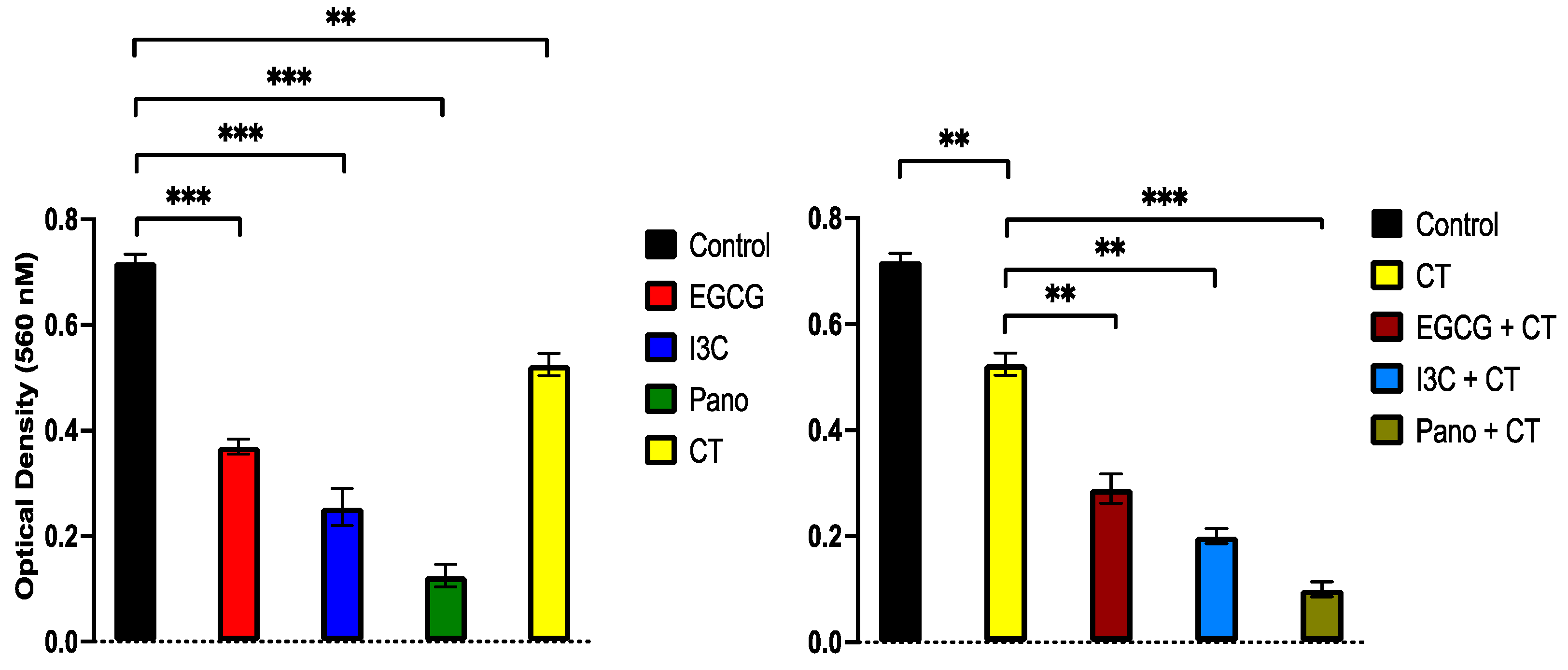
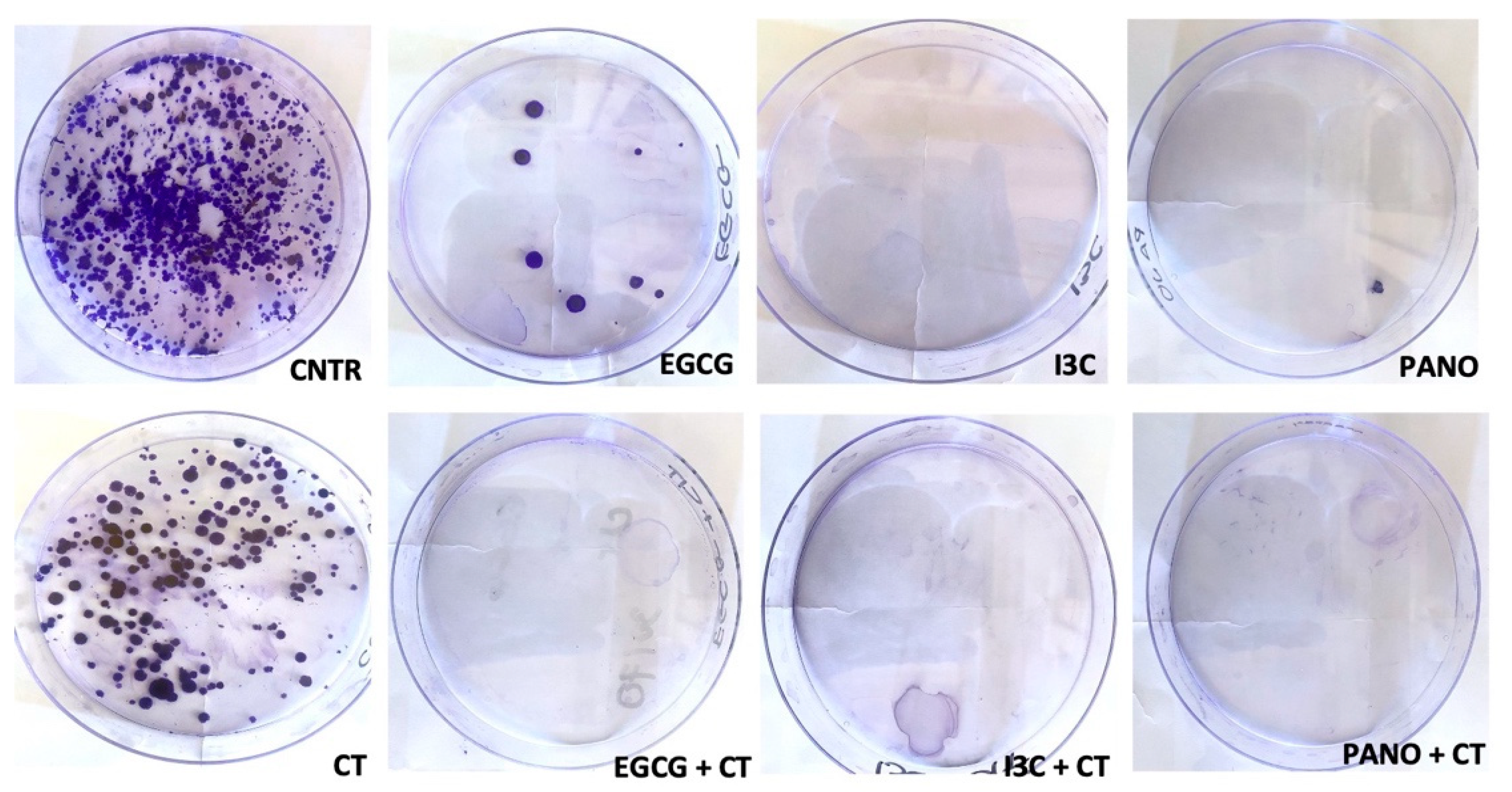
3.6.3. Post-Treatment Ability of 3D-Derived Cells to Form Colony Forming Units (CFU)
3.7. Can Epigenetic Treatments Alter the Expression of Protumorigenic Markers in Ovarian Cancer Human Specimens?
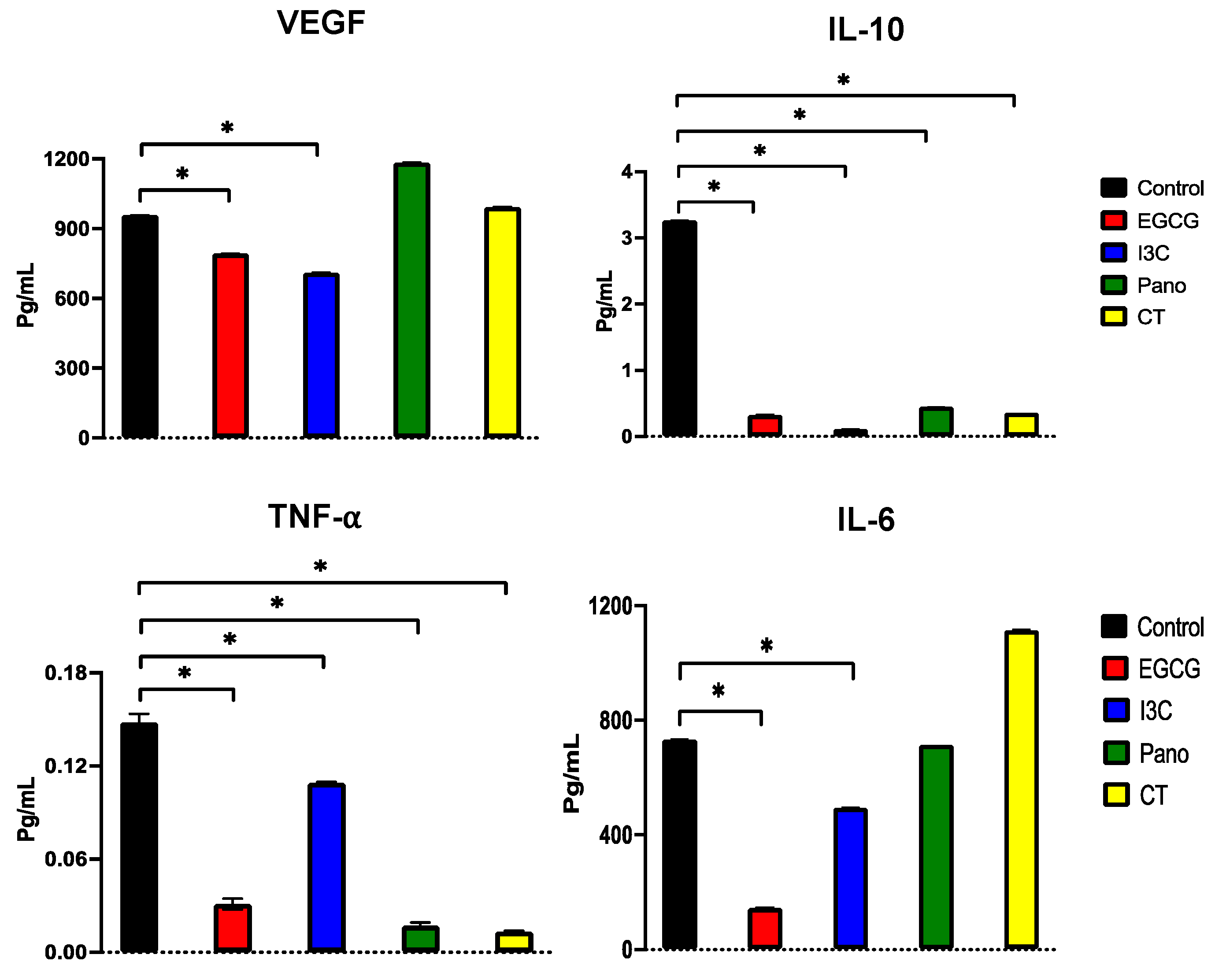
3.7.1. Effects on the Secretion of Protumorigenic Factors by ELISA
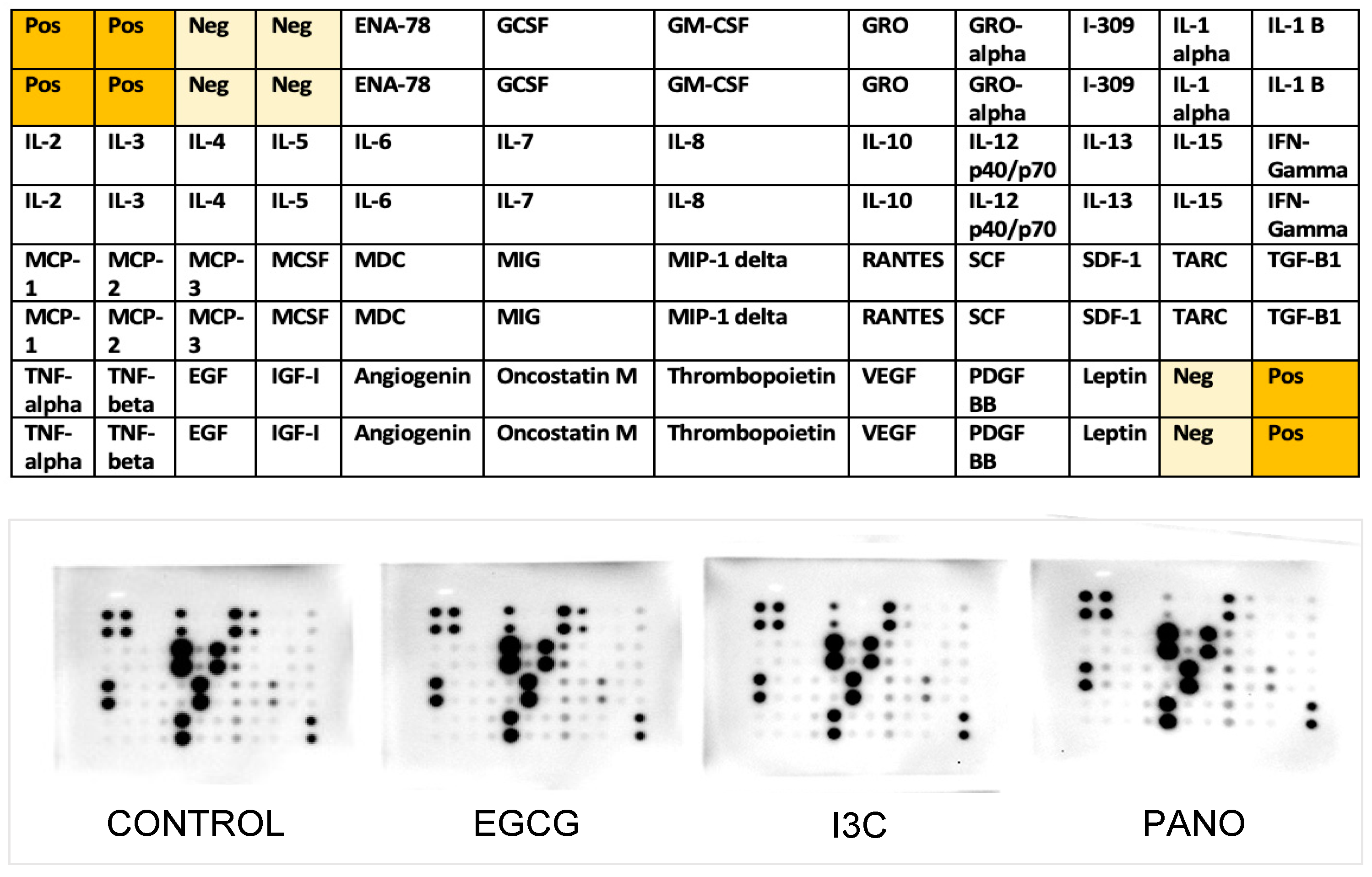
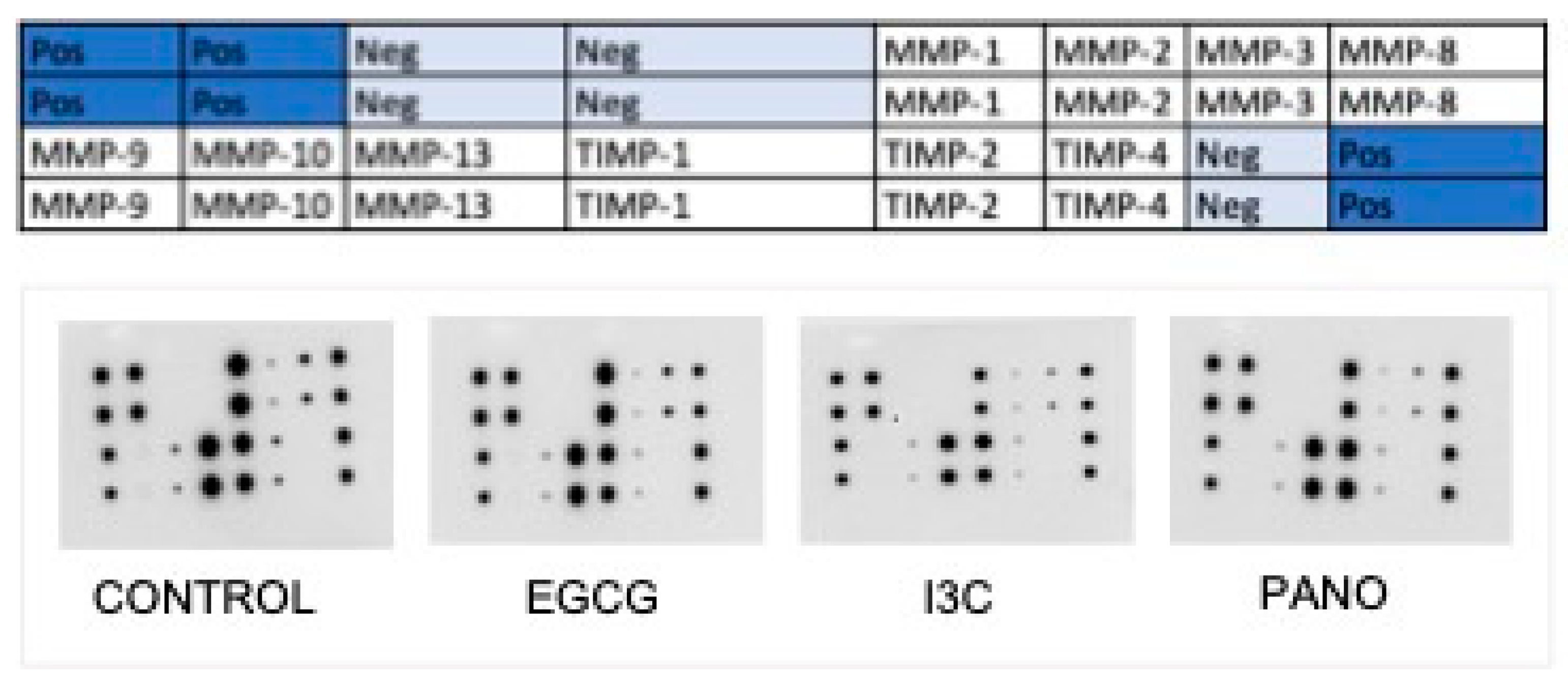
3.7.2. Effects on the Secretion of Cytokines, Growth Factors, MMPs, and TIMPs by Protein Array
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Cancer Society. Cancer Statistics Center. Available online: http://cancerstatisticscenter.cancer.org (accessed on 5 January 2023).
- National Comprehensive Cancer Network. Ovarian Cancer including Fallopian Tube Cancer and Primary Peritoneal Cancer (Version 1.2023). Available online: http://www.nccn.org/professionals/physician_gls/pdf/ovariancancer.pdf (accessed on 10 January 2023).
- Torre, L.A.; Trabert, B.; DeSantis, C.E.; Miller, K.D.; Samimi, G.; Runowicz, C.D.; Gaudet, M.M.; Jemal, A.; Siegel, R.L. Ovarian cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 284–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchetti, C.; Pisano, C.; Facchini, G.; Bruni, G.S.; Magazzino, F.P.; Losito, S.; Pignata, S. First-line treatment of advanced ovarian cancer: Current research and perspectives. Expert Rev. Anticancer. Ther. 2010, 10, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Pokhriyal, R.; Hariprasad, R.; Kumar, L.; Hariprasad, G. Chemotherapy Resistance in Advanced Ovarian Cancer Patients. Biomark. Cancer 2019, 11, 1179299X19860815. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Lin, Y.G.; Roman, L.D.; Sood, A.K. Overcoming platinum resistance in ovarian carcinoma. Expert Opin. Investig. Drugs 2010, 19, 1339–1354. [Google Scholar] [CrossRef] [PubMed]
- Chi, D.S.; Berchuck, A.; Dizon, D.S.; Yashar, C. Principles and Practice of Gynecologic Oncology, 7th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2017. [Google Scholar]
- Balch, C.; Huang, T.H.; Brown, R.; Nephew, K.P. The epigenetics of ovarian cancer drug resistance and resensitization. Am. J. Obstet. Gynecol. 2004, 191, 1552–1572. [Google Scholar] [CrossRef]
- Smith, H.J.; Straughn, J.M.; Buchsbaum, D.J.; Arend, R.C. Epigenetic therapy for the treatment of epithelial ovarian cancer: A clinical review. Gynecol. Oncol. Rep. 2017, 20, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, A.Z.; Afroze, S.H.; Osuji, G.A.; Kayani, S.Y.; Colon, N.; Pantho, A.F.; Kuehl, T.J.; Pilkinton, K.A.; Uddin, M.N. Epigenetic Modifications in Ovarian Cancer: A Review. J. Cancer Treat. Diagn. 2020, 4, 17–35. [Google Scholar] [CrossRef]
- Moufarrij, S.; Dandapani, M.; Arthofer, E.; Gomez, S.; Srivastava, A.; Lopez-Acevedo, M.; Villagra, A.; Chiappinelli, K.B. Epigenetic therapy for ovarian cancer: Promise and progress. Clin. Epigenetics 2019, 11, 7. [Google Scholar] [CrossRef]
- Jones, P.A.; Ohtani, H.; Chakravarthy, A.; De Carvalho, D.D. Epigenetic therapy in immune-oncology. Nat. Rev. Cancer 2019, 19, 151–161. [Google Scholar] [CrossRef]
- Rivenbark, A.G.; Stolzenburg, S.; Beltran, A.S.; Yuan, X.; Rots, M.G.; Strahl, B.D.; Blancafort, P. Epigenetic reprogramming of cancer cells via targeted DNA methylation. Epigenetics 2012, 7, 350–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gujar, H.; Weisenberger, D.J.; Liang, G. The Roles of Human DNA Methyltransferases and Their Isoforms in Shaping the Epigenome. Genes 2019, 10, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucidi, A.; Buca, D.; Ronsini, C.; Tinari, S.; Bologna, G.; Buca, D.; Leombroni, M.; Liberati, M.; D’Antonio, F.; Scambia, G.; et al. Role of Extracellular Vesicles in Epithelial Ovarian Cancer: A Systematic Review. Int. J. Mol. Sci. 2020, 21, 8762. [Google Scholar] [CrossRef] [PubMed]
- Avilés, D.; Warshal, D.; Krill, L.; Ostrovsky, O. Extracellular Vesicles and Ovarian Cancer. In Extracellular Vesicles—Role in Diseases Pathogenesis and Therapy; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Nowicka, A.; Marini, F.C.; Solley, T.N.; Elizondo, P.B.; Zhang, Y.; Sharp, H.J.; Broaddus, R.; Kolonin, M.; Mok, S.C.; Thompson, M.S.; et al. Human omental-derived adipose stem cells increase ovarian cancer proliferation, migration, and chemoresistance. PLoS ONE 2013, 8, e81859. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Wang, Q.; Lau, W.B.; Lau, B.; Xu, L.; Zhao, L.; Yang, H.; Feng, M.; Xuan, Y.; Yang, Y.; et al. Tumor microenvironment: The culprit for ovarian cancer metastasis? Cancer Lett. 2016, 377, 174–182. [Google Scholar] [CrossRef]
- Lis, R.; Touboul, C.; Raynaud, C.M.; Malek, J.A.; Suhre, K.; Mirshahi, M.; Rafii, A. Mesenchymal cell interaction with ovarian cancer cells triggers pro-metastatic properties. PLoS ONE 2012, 7, e38340. [Google Scholar] [CrossRef] [Green Version]
- Nieman, K.M.; Kenny, H.A.; Penicka, C.V.; Ladanyi, A.; Buell-Gutbrod, R.; Zillhardt, M.R.; Romero, I.L.; Carey, M.S.; Mills, G.B.; Hotamisligil, G.S.; et al. Adipocytes promote ovarian cancer metastasis and provide energy for rapid tumor growth. Nat. Med. 2011, 17, 1498–1503. [Google Scholar] [CrossRef] [Green Version]
- Sookram, J.; Zheng, A.; Linden, K.M.; Morgan, A.B.; Brown, S.A.; Ostrovsky, O. Epigenetic therapy can inhibit growth of ovarian cancer cells and reverse chemoresistant properties acquired from metastatic omentum. Int. J. Gynaecol. Obstet. Off. Organ Int. Fed. Gynaecol. Obstet. 2019, 145, 225–232. [Google Scholar] [CrossRef]
- Kapur, S.K.; Katz, A.J. Review of the adipose derived stem cell secretome. Biochimie 2013, 95, 2222–2228. [Google Scholar] [CrossRef]
- Kelly, T.K.; De Carvalho, D.D.; Jones, P.A. Epigenetic modifications as therapeutic targets. Nat. Biotechnol. 2010, 28, 1069–1078. [Google Scholar] [CrossRef] [Green Version]
- Jin, N.; George, T.L.; Otterson, G.A.; Verschraegen, C.; Wen, H.; Carbone, D.; Herman, J.; Bertino, E.M.; He, K. Advances in epigenetic therapeutics with focus on solid tumors. Clin. Epigenetics 2021, 13, 83. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Nephew, K.P. Ovarian Cancer Stem Cells: Role in Metastasis and Opportunity for Therapeutic Targeting. Cancers 2019, 11, 934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matei, D.E.; Nephew, K.P. Epigenetic therapies for chemoresensitization of epithelial ovarian cancer. Gynecol. Oncol. 2010, 116, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Hunter, K.; Aikins, J.; Brown, S.; Ostrovsky, O. Use of Epigenetic Therapy Shortens Duration of Standard Chemotherapy for Ovarian Cancer with Minimal Toxicity to Normal Tissue. J. Cancer Sci. Clin. Ther. 2020, 4, 100–114. [Google Scholar] [CrossRef]
- Bilbao, M.; Katz, C.; Kass, S.L.; Smith, D.; Hunter, K.; Warshal, D.; Aikins, J.K.; Ostrovsky, O. Epigenetic Therapy Augments Classic Chemotherapy in Suppressing the Growth of 3D High-Grade Serous Ovarian Cancer Spheroids over an Extended Period of Time. Biomolecules 2021, 11, 1711. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hu, W.; Shen, D.Y.; Kavanagh, J.J.; Fu, S. Azacitidine enhances sensitivity of platinum-resistant ovarian cancer cells to carboplatin through induction of apoptosis. Am. J. Obstet. Gynecol. 2009, 200, 177.e1–177.e9. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.Y.; Liao, W.S.; Lu, Z.; Bornmann, W.G.; Hennessey, V.; Washington, M.N.; Rosner, G.L.; Yu, Y.; Ahmed, A.A.; Bast, R.C., Jr. Decitabine and suberoylanilide hydroxamic acid (SAHA) inhibit growth of ovarian cancer cell lines and xenografts while inducing expression of imprinted tumor suppressor genes, apoptosis, G2/M arrest, and autophagy. Cancer 2011, 117, 4424–4438. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.; Hu, W.; Iyer, R.; Kavanagh, J.J.; Coleman, R.L.; Levenback, C.F.; Sood, A.K.; Wolf, J.K.; Gershenson, D.M.; Markman, M.; et al. Phase 1b-2a study to reverse platinum resistance through use of a hypomethylating agent, azacitidine, in patients with platinum-resistant or platinum-refractory epithelial ovarian cancer. Cancer 2011, 117, 1661–1669. [Google Scholar] [CrossRef]
- Helland, Ø.; Popa, M.; Bischof, K.; Gjertsen, B.T.; McCormack, E.; Bjørge, L. The HDACi Panobinostat Shows Growth Inhibition Both In Vitro and in a Bioluminescent Orthotopic Surgical Xenograft Model of Ovarian Cancer. PLoS ONE 2016, 11, e0158208. [Google Scholar] [CrossRef] [Green Version]
- Garrett, L.A.; Growdon, W.B.; Rueda, B.R.; Foster, R. Influence of a novel histone deacetylase inhibitor panobinostat (LBH589) on the growth of ovarian cancer. J. Ovarian Res. 2016, 9, 58. [Google Scholar] [CrossRef] [Green Version]
- Weng, J.R.; Tsai, C.H.; Kulp, S.K.; Chen, C.S. Indole-3-carbinol as a chemopreventive and anti-cancer agent. Cancer Lett. 2008, 262, 153–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidel, C.; Schnekenburger, M.; Dicato, M.; Diederich, M. Histone deacetylase modulators provided by Mother Nature. Genes Nutr. 2012, 7, 357–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royston, K.J.; Tollefsbol, T.O. The Epigenetic Impact of Cruciferous Vegetables on Cancer Prevention. Curr. Pharmacol. Rep. 2015, 1, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Trudel, D.; Labbé, D.P.; Bairati, I.; Fradet, V.; Bazinet, L.; Têtu, B. Green tea for ovarian cancer prevention and treatment: A systematic review of the in vitro, in vivo and epidemiological studies. Gynecol. Oncol. 2012, 126, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.Z.; Wang, Y.; Ai, N.; Hou, Z.; Sun, Y.; Lu, H.; Welsh, W.; Yang, C.S. Tea polyphenol (-)-epigallocatechin-3-gallate inhibits DNA methyltransferase and reactivates methylation-silenced genes in cancer cell lines. Cancer Res. 2003, 63, 7563–7570. [Google Scholar]
- Katz, E.; Nisani, S.; Chamovitz, D.A. Indole-3-carbinol: A plant hormone combatting cancer. F1000Research 2018, 7, F1000 Faculty Rev-689. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.J.; Shim, J.Y.; Zhu, B.T. Mechanisms for the inhibition of DNA methyltransferases by tea catechins and bioflavonoids. Mol. Pharmacol. 2005, 68, 1018–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, J.; Shi, W.; Guo, H.; Long, W.; Wang, Y.; Qi, J.; Liu, J.; Xu, Y. The Inhibitory Effect of (-)-Epigallocatechin-3-Gallate on Breast Cancer Progression via Reducing SCUBE2 Methylation and DNMT Activity. Molecules 2019, 24, 2899. [Google Scholar] [CrossRef] [Green Version]
- Nandakumar, V.; Vaid, M.; Katiyar, S.K. (-)-Epigallocatechin-3-gallate reactivates silenced tumor suppressor genes, Cip1/p21 and p16INK4a, by reducing DNA methylation and increasing histones acetylation in human skin cancer cells. Carcinogenesis 2011, 32, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Zwergel, C.; Valente, S.; Mai, A. DNA Methyltransferases Inhibitors from Natural Sources. Curr. Top. Med. Chem. 2016, 16, 680–696. [Google Scholar] [CrossRef]
- Huang, Y.J.; Wang, K.L.; Chen, H.Y.; Chiang, Y.F.; Hsia, S.M. Protective Effects of Epigallocatechin Gallate (EGCG) on Endometrial, Breast, and Ovarian Cancers. Biomolecules 2020, 10, 1481. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.H.; Lavrenov, S.N.; Sundar, S.N.; Nguyen, D.H.; Tseng, M.; Marconett, C.N.; Kung, J.; Staub, R.E.; Preobrazhenskaya, M.N.; Bjeldanes, L.F.; et al. 1-Benzyl-indole-3-carbinol is a novel indole-3-carbinol derivative with significantly enhanced potency of anti-proliferative and anti-estrogenic properties in human breast cancer cells. Chem. Biol. Interact. 2010, 186, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Cheng, P.H.; Rao, X.M.; McMasters, K.M.; Zhou, H.S. Indole-3-carbinol (I3C) increases apoptosis, represses growth of cancer cells, and enhances adenovirus-mediated oncolysis. Cancer Biol. Ther. 2014, 15, 1256–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiselev, V.I.; Ashrafyan, L.A.; Muyzhnek, E.L.; Gerfanova, E.V.; Antonova, I.B.; Aleshikova, O.I.; Sarkar, F.H. A new promising way of maintenance therapy in advanced ovarian cancer: A comparative clinical study. BMC Cancer 2018, 18, 904. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, F.H.; Li, Y. Indole-3-carbinol and prostate cancer. J. Nutr. 2004, 134, 3493S–3498S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, G.J.; Zhang, Z.; Wen, X.D.; Yu, C.; Calway, T.; Yuan, C.S.; Wang, C.Z. Epigallocatechin Gallate (EGCG) is the most effective cancer chemopreventive polyphenol in green tea. Nutrients 2012, 4, 1679–1691. [Google Scholar] [CrossRef] [PubMed]
- Kelm, J.M.; Timmins, N.E.; Brown, C.J.; Fussenegger, M.; Nielsen, L.K. Method for generation of homogeneous multicellular tumor spheroids applicable to a wide variety of cell types. Biotechnol. Bioeng. 2003, 83, 173–180. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Knoblich, J.A. Organogenesis in a dish: Modeling development and disease using organoid technologies. Science 2014, 345, 1247125. [Google Scholar] [CrossRef]
- Kim, J.; Koo, B.K.; Knoblich, J.A. Human organoids: Model systems for human biology and medicine. Nat. Rev. Mol. Cell Biol. 2020, 21, 571–584. [Google Scholar] [CrossRef]
- Madden, E.C.; Gorman, A.M.; Logue, S.E.; Samali, A. Tumour Cell Secretome in Chemoresistance and Tumour Recurrence. Trends Cancer 2020, 6, 489–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López de Andrés, J.; Griñán-Lisón, C.; Jiménez, G.; Marchal, J.A. Cancer stem cell secretome in the tumor microenvironment: A key point for an effective personalized cancer treatment. J. Hematol. Oncol. 2020, 13, 136. [Google Scholar] [CrossRef] [PubMed]
- Escalona, R.M.; Kannourakis, G.; Findlay, J.K.; Ahmed, N. Expression of TIMPs and MMPs in Ovarian Tumors, Ascites, Ascites-Derived Cells, and Cancer Cell Lines: Characteristic Modulatory Response before and after Chemotherapy Treatment. Front. Oncol. 2022, 11, 796588. [Google Scholar] [CrossRef] [PubMed]
- Nash, M.A.; Ferrandina, G.; Gordinier, M.; Loercher, A.; Freedman, R.S. The role of cytokines in both the normal and malignant ovary. Endocr. Relat. Cancer 1999, 6, 93–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, D.; Matte, I.; Garde-Granger, P.; Bessette, P.; Piché, A. Ascites IL-10 Promotes Ovarian Cancer Cell Migration. Cancer Microenviron. Off. J. Int. Cancer Microenviron. Soc. 2018, 11, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Hart, K.M.; Byrne, K.T.; Molloy, M.J.; Usherwood, E.M.; Berwin, B. IL-10 immunomodulation of myeloid cells regulates a murine model of ovarian cancer. Front. Immunol. 2011, 2, 29. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, R.C.; Zhang, X.L.; Niu, X.L.; Qu, Y.; Li, L.Z.; Meng, X.Y. Interleukin-8 secretion by ovarian cancer cells increases anchorage-independent growth, proliferation, angiogenic potential, adhesion and invasion. Cytokine 2012, 59, 145–155. [Google Scholar] [CrossRef]
- Zhang, R.; Roque, D.M.; Reader, J.; Lin, J. Combined inhibition of IL-6 and IL-8 pathways suppresses ovarian cancer cell viability and migration and tumor growth. Int. J. Oncol. 2022, 60, 50. [Google Scholar] [CrossRef]
- Zhou, W.; Sun, W.; Yung, M.M.H.; Dai, S.; Cai, Y.; Chen, C.W.; Meng, Y.; Lee, J.B.; Braisted, J.C.; Xu, Y.; et al. Autocrine activation of JAK2 by IL-11 promotes platinum drug resistance. Oncogene 2018, 37, 3981–3997. [Google Scholar] [CrossRef] [PubMed]
- Souza-Fonseca-Guimaraes, F.; Cursons, J.; Huntington, N.D. The Emergence of Natural Killer Cells as a Major Target in Cancer Immunotherapy. Trends Immunol. 2019, 40, 142–158. [Google Scholar] [CrossRef]
- Juurikka, K.; Butler, G.S.; Salo, T.; Nyberg, P.; Åström, P. Role of MMP8 in Cancer: A Systematic Review. Int. J. Mol. Sci. 2019, 20, 4506. [Google Scholar] [CrossRef] [Green Version]
- Al-Alem, L.; Curry, T.E., Jr. Ovarian cancer: Involvement of the matrix metalloproteinases. Reproduction 2015, 150, R55–R64. [Google Scholar] [CrossRef] [Green Version]
- Freese, K.E.; Kokai, L.; Edwards, R.P.; Philips, B.J.; Sheikh, M.A.; Kelley, J.; Comerci, J.; Marra, K.G.; Rubin, J.P.; Linkov, F. Adipose-derived stems cells and their role in human cancer development, growth, progression, and metastasis: A systematic review. Cancer Res. 2015, 75, 1161–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klopp, A.H.; Zhang, Y.; Solley, T.; Amaya-Manzanares, F.; Marini, F.; Andreeff, M.; Debeb, B.; Woodward, W.; Schmandt, R.; Broaddus, R.; et al. Omental adipose tissue-derived stromal cells promote vascularization and growth of endometrial tumors. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 771–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bimonte, S.; Cascella, M. The Potential Roles of Epigallocatechin-3-Gallate in the Treatment of Ovarian Cancer: Current State of Knowledge. Drug Des. Dev. Ther. 2020, 14, 4245–4250. [Google Scholar] [CrossRef] [PubMed]
- Siblini, H.; Al-Hendy, A.; Segars, J.; González, F.; Taylor, H.S.; Singh, B.; Flaminia, A.; Flores, V.A.; Christman, G.M.; Huang, H.; et al. Assessing the Hepatic Safety of Epigallocatechin Gallate (EGCG) in Reproductive-Aged Women. Nutrients 2023, 15, 320. [Google Scholar] [CrossRef] [PubMed]
- National Toxicology Program. Toxicology studies of indole-3-carbinol in F344/N rats and B6C3F1/N mice and toxicology and carcinogenesis studies of indole-3-carbinol in Harlan Sprague Dawley rats and B6C3F1/N mice (gavage studies). Natl. Toxicol. Program Tech. Rep. Ser. 2017, 584, NTP-TR-584. [Google Scholar] [CrossRef]
- Wong, G.Y.; Bradlow, L.; Sepkovic, D.; Mehl, S.; Mailman, J.; Osborne, M.P. Dose-ranging study of indole-3-carbinol for breast cancer prevention. J. Cell. Biochem. Suppl. 1997, 28–29, 111–116. [Google Scholar] [CrossRef]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GF/ CYTOKINE | FUNCTION | TREATMENTS | ||||||
|---|---|---|---|---|---|---|---|---|
| CONTROL | EGCG | I3C | PANO | |||||
| MEAN, SD | MEAN, SD | EFFECT | MEAN, SD | EFFECT | MEAN, SD | EFFECT | ||
| IL-10 | Induces cancer proliferation, migration, and immunosuppression | 183 ± 5 | 42 ± 4 | 77%  | 50 ± 9.3 | 73% | 40 ± 1.5 | 78% |
| IL-8 | Stimulates ovarian cancer growth and proliferation | 187± 0.25 | 150 ± 0.46 | 20% | 167 ± 5.7 | 11% | 139 ± 5 | 25% |
| IL-11 | Promotes chemoresistance, proliferation and survival of metastatic ovarian cancer cells | 36 ± 3.6 | 18 ± 2.7 | 50% | 2.9 ± 2.4 | 92% | 0.8 ± 0.2 | 98% |
| IL-6R | Correlated with aggressive behavior of high-grade ovarian cancer and poor prognosis when activated by IL-6 | 35.2 ± 3.6 | 41.6 ± 4.23 | 18%  | 21.9 ± 2 | 38% | 34.78 ± 1.6 | 22% |
| TNF-RI | Stimulates release of IL-6 and VEGF, induces peritoneal cancer spread | 136 ± 2.2 | 30.5 ± 4.6 | 78% | 7.8 ± 2.5 | 94% | 12.2 ± 1.3 | 91% |
| RANTES (CCL5) | Alters T-cells and NK cells function, induces cancer progression and resistance | 136 ± 2.4 | 5.33 ± 1.3 | 96% | 38.8 ± 0.91 | 71% | 31.7 ± 1.46 | 77% |
| MCP-1 | Facilitates proliferation, migration, angiogenesis and immunosuppression | 134 ± 2.3 | 135.5 ± 4.9 | 11% | 80.6 ± 4.3 | 40% | 81.7 ± 1.4 | 39% |
| MMPs/TIMPs | FUNCTION | TREATMENTS | ||||||
|---|---|---|---|---|---|---|---|---|
| CONTROL | EGCG | I3C | PANO | |||||
| MEAN, SD | MEAN, SD | EFFECT | MEAN, SD | EFFECT | MEAN, SD | EFFECT | ||
| MMP-1 | Degradation of ECM | 31,359 ± 518 | 33,999 ± 2477 | 8% | 22,850 ± 1061 | 27% | 30,443 ± 2035 | 3% |
| MMP-3 | Remodeling of ECM | 10,777 ± 752 | 10,209 ± 588 | 5% | 5391 ± 109 | 50% | 5083 ± 283 | 53% |
| MMP-8 | Stimulates inflammation, correlates with prognosis | 23,777 ± 1229 | 16,732 ± 1456 | 30% | 19,900 ± 2347 | 16% | 25,445 ± 486 | 7% |
| MMP-13 | Stimulates metastasis, correlates with prognosis | 4467 ± 331 | 3094 ± 411 | 31% | 2871 ± 910 | 36% | 2950 ± 186 | 34% |
| TIMP-1 | Inhibits metalloproteinase activity, cancer proliferation, migration, invasion | 30,750 ± 1132 | 32,433 ± 1988 | 5% | 32,011 ± 2774 | 4% | 32,509 ± 988 | 6% |
| TIMP-2 | Inhibits metalloproteinase activity, cancer proliferation, migration, invasion | 23,552 ± 1099 | 25,728 ± 1834 | 9% | 28,211 ± 4007 | 19% | 33,889 ± 636 | 43% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kelly, R.; Aviles, D.; Krisulevicz, C.; Hunter, K.; Krill, L.; Warshal, D.; Ostrovsky, O. The Effects of Natural Epigenetic Therapies in 3D Ovarian Cancer and Patient-Derived Tumor Explants: New Avenues in Regulating the Cancer Secretome. Biomolecules 2023, 13, 1066. https://doi.org/10.3390/biom13071066
Kelly R, Aviles D, Krisulevicz C, Hunter K, Krill L, Warshal D, Ostrovsky O. The Effects of Natural Epigenetic Therapies in 3D Ovarian Cancer and Patient-Derived Tumor Explants: New Avenues in Regulating the Cancer Secretome. Biomolecules. 2023; 13(7):1066. https://doi.org/10.3390/biom13071066
Chicago/Turabian StyleKelly, Rebeca, Diego Aviles, Catriona Krisulevicz, Krystal Hunter, Lauren Krill, David Warshal, and Olga Ostrovsky. 2023. "The Effects of Natural Epigenetic Therapies in 3D Ovarian Cancer and Patient-Derived Tumor Explants: New Avenues in Regulating the Cancer Secretome" Biomolecules 13, no. 7: 1066. https://doi.org/10.3390/biom13071066
APA StyleKelly, R., Aviles, D., Krisulevicz, C., Hunter, K., Krill, L., Warshal, D., & Ostrovsky, O. (2023). The Effects of Natural Epigenetic Therapies in 3D Ovarian Cancer and Patient-Derived Tumor Explants: New Avenues in Regulating the Cancer Secretome. Biomolecules, 13(7), 1066. https://doi.org/10.3390/biom13071066