
Galectin-9 as a Potential Modulator of Lymphocyte Adhesion to Endothelium via Binding to Blood Group H Glycan

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Reagents
2.2. Galectins and Galectin-Specific Antibodies
2.3. Cell Culture
2.4. Enzymatic Defucosylation of EA.hy 926
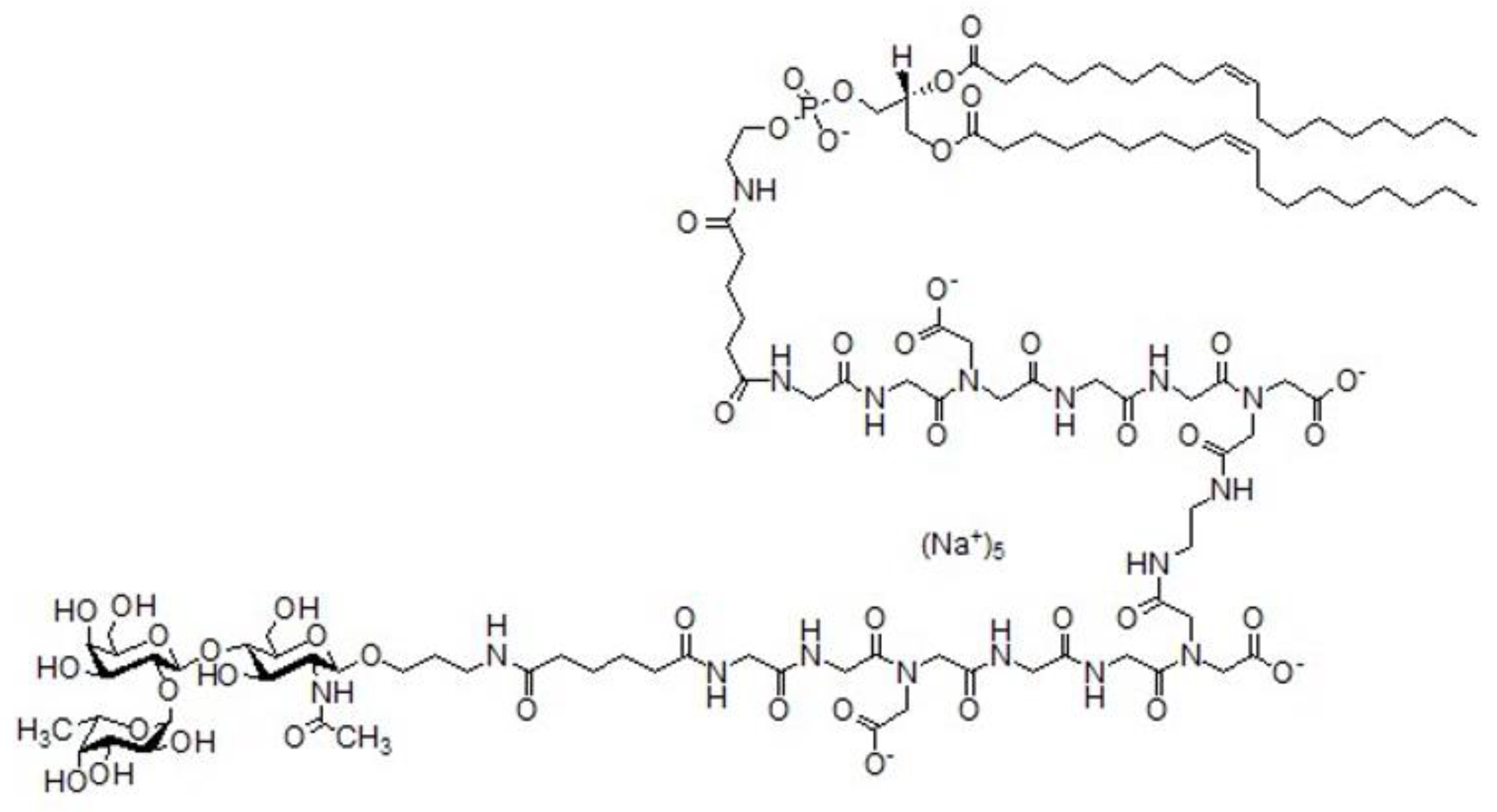
2.5. Insertion of FSL-H (Type 2) into EA.hy 926 Cells
2.6. Binding of Galectins to EA.hy 926 Cells
2.7. Flow Cytometry
2.8. Detection of FSL-H (Type 2) in the Cell Membrane
2.9. Cell Adhesion Assay
2.10. Statistical Analysis
3. Results
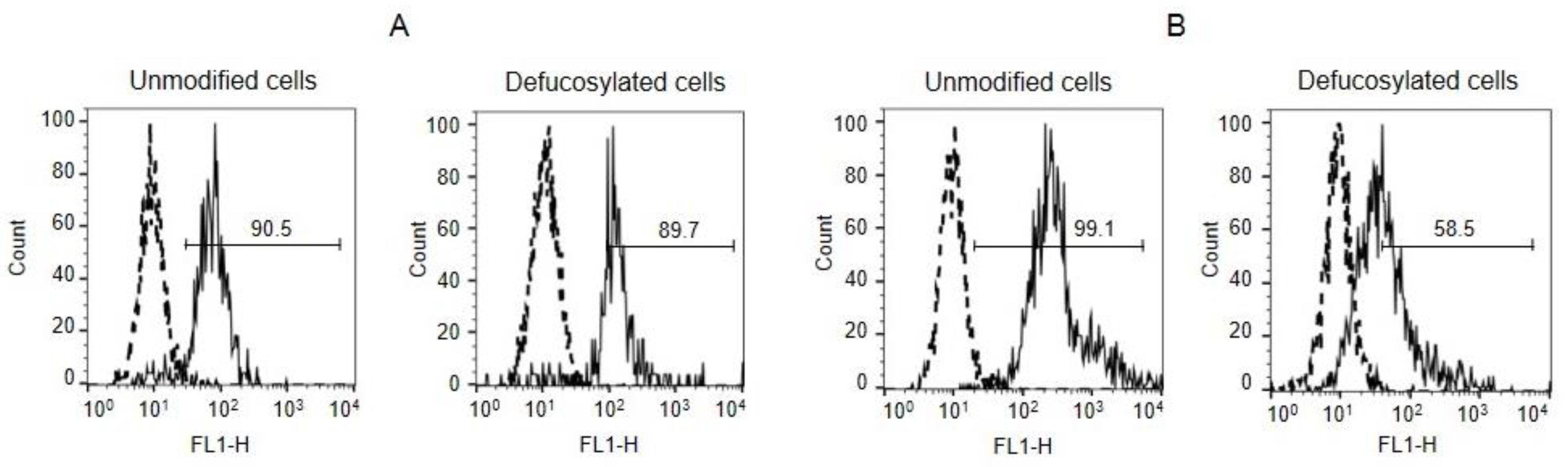
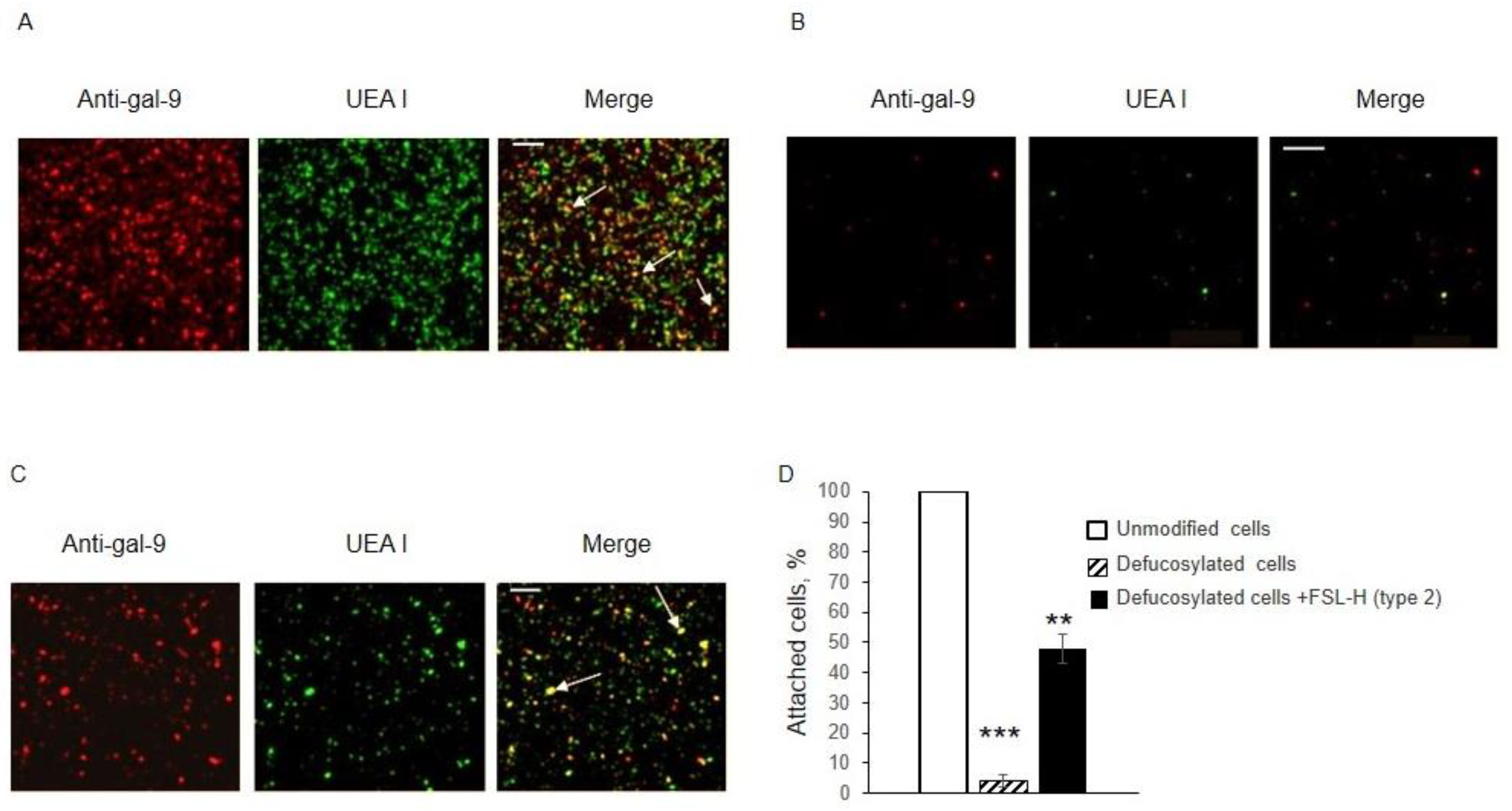
3.1. Binding of Galectins to Unmodified and Fucosidase-Treated EA.hy 926 Cells
3.2. Binding of Gal-9 to Defucosylated EA.hy 926 Cells after Their “Refucosylation” with FSL-H (Type 2)
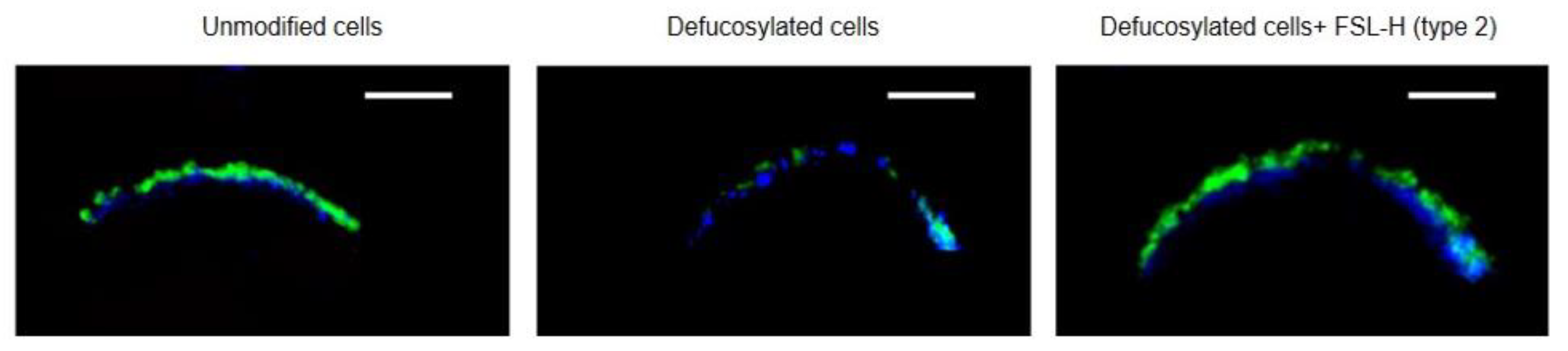
3.3. Localization of Inserted FSL-H (Type 2)
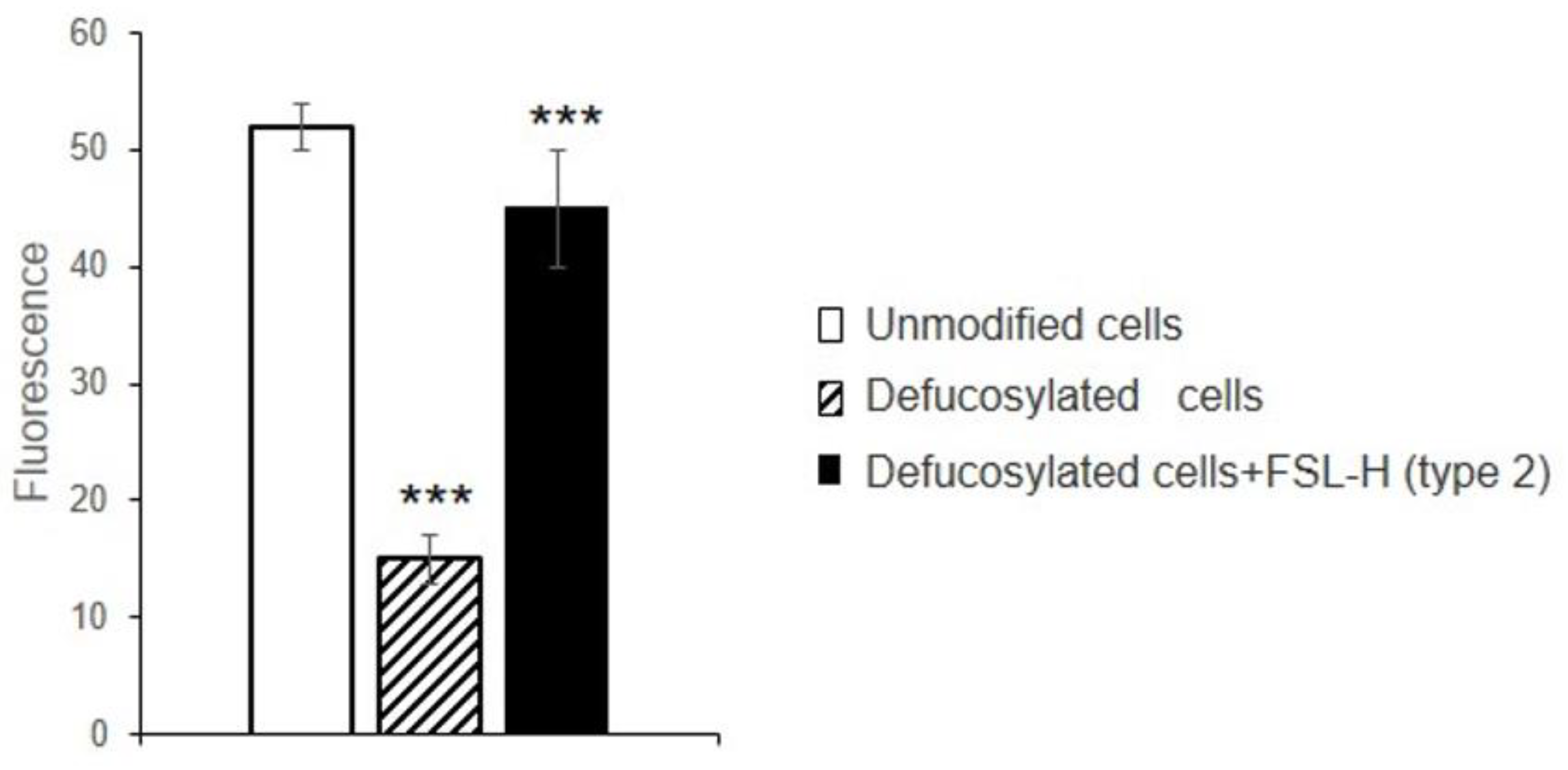
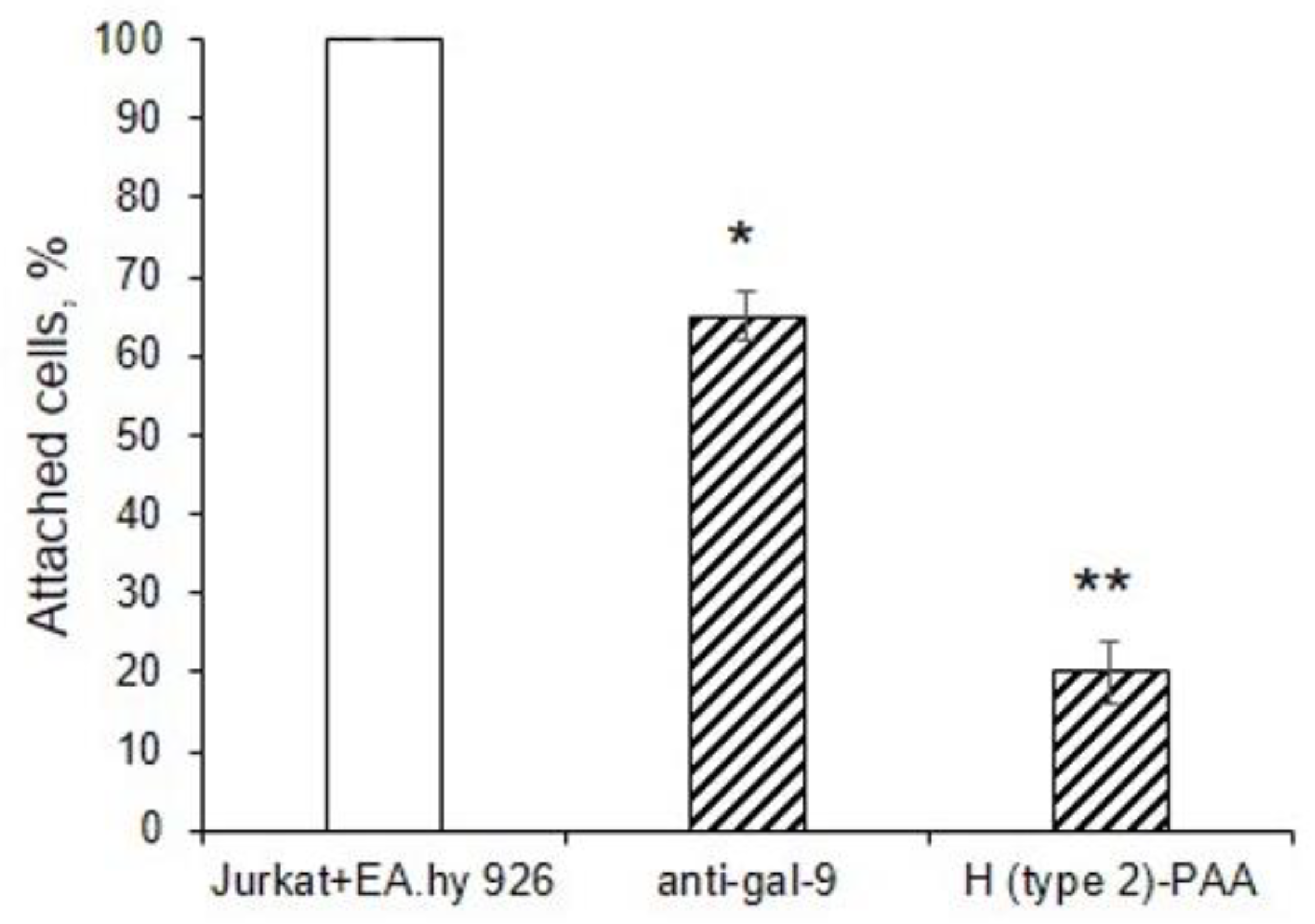
3.4. Gal-9 Promotes Adhesion of Jurkat Cells to EA.hy 926 Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Langer, H.F.; Chavakis, T. Leukocyte–endothelial interactions in inflammation. J. Cell. Mol. Med. 2009, 13, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- DeGrendele, H.C.; Estess, P.; Picker, L.J.; Siegelmanand, M.H. CD44 and its ligand hyaluronate mediate rolling under physiologic flow: A novel lymphocyte-endothelial cell primary adhesion pathway. J. Exp. Med. 1996, 183, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.I.; Kerfoot, S.M.; Heit, B.; Liu, L.; Andonegui, G.; Ruffell, B.; Johnson, P.; Kubes, P. Role of CD44 and hyaluronan in neutrophil recruitment. J. Immunol. 2004, 173, 7594–7601. [Google Scholar] [CrossRef] [Green Version]
- Johnson, P.; Ruffell, B. CD44 and its role in inflammation and inflammatory diseases. Inflamm. Allergy Drug Targets 2009, 8, 208–220. [Google Scholar] [CrossRef]
- Rosemblatt, M.; Bono, M.R. Functional consequences of immune cell adhesion to endothelial cells. Curr. Pharm. Des. 2004, 10, 109–120. [Google Scholar] [CrossRef]
- Filippi, M.-D. Neutrophil transendothelial migration: Updates and new perspectives. Blood 2019, 133, 2149–2158. [Google Scholar] [CrossRef] [PubMed]
- Elola, M.T.; Wolfenstein-Todel, C.; Troncoso, M.F.; Vasta, G.R.; Rabinovich, G.A. Galectins: Matricellular glycan-binding proteins linking cell adhesion, migration, and survival. Cell. Mol. Life Sci. 2007, 64, 1679–1700. [Google Scholar] [CrossRef]
- Rao, S.P.; Wang, Z.; Zuberi, R.I.; Sikora, L.; Bahaie, N.S.; Zuraw, B.L.; Liu, F.-T.; Sriramarao, P. Galectin-3 Functions as an adhesion molecule to support eosinophil rolling and adhesion under conditions of flow. J. Immunol. 2007, 179, 7800–7807. [Google Scholar] [CrossRef]
- Thijssen, V.L. Galectins in endothelial cell biology and angiogenesis: The basics. Biomolecules 2021, 11, 1386. [Google Scholar] [CrossRef]
- Colomb, F.; Wang, W.; Simpson, D.; Zafar, M.; Beynon, R.; Rhodes, J.M.; Yu, L.-G. Galectin-3 interacts with the cell-surface glycoprotein CD146 (MCAM, MUC18) and induces secretion of metastasispromoting cytokines from vascular endothelial cells. J. Biol. Chem. 2017, 292, 8381–8389. [Google Scholar] [CrossRef] [Green Version]
- Cueni, L.N.; Detmar, M. Galectin-8 interacts with podoplanin and modulates lymphatic endothelial cell functions. Exp. Cell Res. 2009, 315, 1715–1723. [Google Scholar] [CrossRef] [Green Version]
- Fernández, M.M.; Ferragut, F.; Delgado, V.M.; Bracalente, C.; Bravo, A.I.; Cagnoni, A.J.; Nuñez, M.; Morosi, L.G.; Quinta, H.R.; Espelt, M.V.; et al. Glycosylation-dependent binding of galectin-8 to activated leukocyte cell adhesion molecule (ALCAM/CD166) promotes its surface segregation on breast cancer cells. Biochim. Biophys. Acta 2016, 1860, 2255–2268. [Google Scholar] [CrossRef]
- O’Donnell, J.; Mille-Baker, B.; Laffan, M. Human umbilical vein endothelial cells differ from other endothelial cells in failing to express ABO blood group antigens. J. Vasc. Res. 2000, 37, 540–547. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, J.; Laffan, M.A. The relationship between ABO histo-blood group, factor VIII and von Willebrand factor. Transfusion Med. 2001, 11, 343–351. [Google Scholar] [CrossRef]
- Oriol, R.; Candelier, J.J.; Mollicone, R. Molecular genetics of H. Vox Sang 2000, 78 (Suppl. S2), 105–108. [Google Scholar] [PubMed]
- Stowell, S.R.; Arthur, C.M.; Dias-Baruffi, M.; Rodrigues, L.; Gourdine, J.-P.; Heimburg-Molinaro, J.; Ju, T.; Molinaro, R.J.; Rivera-Marrero, C.; Xia, B.; et al. Innate immune lectins kill bacteria expressing blood group antigen. Nat. Med. 2010, 16, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.C.; Ho, A.D.; Kamili, N.A.; Wang, J.; Murdock, K.L.; Cummings, R.D.; Arthur, C.M.; Stowell, S.R. Full-length galectin-3 is required for high affinity microbial interactions and antimicrobial activity. Front. Microbiol 2021, 12, 731026. [Google Scholar] [CrossRef]
- Baranska, P.; Jerczynska, H.; Pawlowska, Z.; Koziokiewicz, W.; Cierniewski, C.S. Expression of integrins and adhesive properties of human endothelial cell line EA.hy 926. Cancer Gen. Prot. 2005, 2, 265–270. [Google Scholar]
- Data of Consortium for Functional Glycomics. Available online: https://www.functionalglycomics.org (accessed on 11 July 2012).
- Korchagina, E.; Tuzikov, A.; Formanovsky, A.; Popova, I.; Henry, S.; Bovin, N. Toward creating cell membrane glyco-landscapes with glycan lipid constructs. Carb. Res. 2012, 356, 238–246. [Google Scholar] [CrossRef]
- Gabius, H.-J. Influence of type of linkage and spacer on the interaction of β-galactoside-binding proteins with immobilized affinity ligands. Anal. Biochem. 1990, 189, 91–94. [Google Scholar] [CrossRef]
- André, S.; Sansone, F.; Kaltner, H.; Casnati, A.; Kopitz, J.; Gabius, H.-J.; Ungaro, R. Calix[n]arene-based glycoclusters: Bioactivity of thiourea-linked galactose/lactose moieties as inhibitors of binding of medically relevant lectins to a glycoprotein and cell-surface glycoconjugates and selectivity among human adhesion/growth-regulatory galectins. ChemBioChem 2008, 9, 1649–1661. [Google Scholar] [CrossRef] [PubMed]
- Schwarzmann, G.; Hoffmann-Bleihauer, P.; Schubert, J.; Sandhoff, K.; Marsh, D. Incorporation of ganglioside analogues into fibroblast cell membranes. A spin-label study. Biochemistry 1983, 22, 5041–5048. [Google Scholar] [CrossRef]
- Oliver, C.; Blake, D.; Henry, S. In vivo neutralization of anti-A and successful transfusion of A antigen incompatible red cells in an animal model. Transfusion 2011, 51, 2664–2675. [Google Scholar] [CrossRef]
- Rapoport, E.M.; André, S.; Kurmyshkina, O.V.; Pochechueva, T.V.; Severov, V.V.; Pazynina, G.V.; Gabius, H.-J.; Bovin, N.V. Galectin-loaded cells as a platform for the profiling of lectin specificity by fluorescent neoglycoconjugates: A case study on galectins-1 and -3 and the impact of assay setting. Glycobiology 2008, 18, 315–324. [Google Scholar] [CrossRef] [Green Version]
- do Canto, A.; Robalo, J.; Santos, B.; Carvalho, A.; Ramalho, J.; Loura, L. Diphenylhexatriene membrane probes DPH and TMA-DPH: A comparative molecular dynamics simulation study. Biochim. Biophys. Acta 2016, 1858, 2647–2661. [Google Scholar] [CrossRef] [PubMed]
- Vokhmyanina, O.A.; Rapoport, E.M.; André, S.; Severov, V.V.; Ryzhov, I.M.; Pazynina, G.V.; Korchagina, E.J.; Gabius, H.-J.; Bovin, N.V. Comparative study of the glycan specificities of cell-bound human tandem repeat-type galectins -4, -8 and -9. Glycobiology 2012, 22, 1207–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapoport, E.M.; Bovin, N.V. Specificity of human galectins on cell surface. Biochemistry 2015, 80, 846–856. [Google Scholar] [CrossRef]
- Zarbock, A.; Ley, K.; McEver, R.P.; Hidalgo, A. Leukocyte ligands for endothelial selectins: Specialized glycoconjugates that mediate rolling and signaling under flow. Blood 2011, 118, 6743–6751. [Google Scholar] [CrossRef] [Green Version]
- Eastlund, T. The histo-blood group ABO system and tissue transplantation. Transfusion 1998, 38, 975–988. [Google Scholar] [CrossRef]
- Kumar, S.; Frank, M.; Schwartz-Albiez, R. Understanding the specificity of human galectin-8C domain interactions with its glycan ligands based on molecular dynamics simulations. PLoS ONE 2013, 8, e59761. [Google Scholar] [CrossRef] [Green Version]
- Bum-Erdene, K.; Leffler, H.; Nilsson, U.J.; Blanchard, H. Structural characterization of human galectin-4 C-terminal domain: Elucidating the molecular basis for recognition of glycosphingolipids, sulfated saccharides and blood group antigens. FEBS J. 2015, 282, 3348–3367. [Google Scholar] [CrossRef] [PubMed]
- Bum-Erdene, K.; Leffler, H.; Nilsson, U.J.; Blanchard, H. Structural characterisation of human galectin-4 N-terminal carbohydrate recognition domain in complex with glycerol, lactose, 3′-sulfo-lactose, and 2′-fucosyllactose. Sci. Rep. 2016, 6, 20289. [Google Scholar] [CrossRef] [Green Version]
- Hirabayashi, J.; Hashidate, T.; Arata, Y.; Nishi, N.; Nakamura, T.; Hirashima, M.; Urashima, T.; Oka, T.; Futai, M.; Muller, W.E.G.; et al. Oligosaccharide specificity of galectins: A search by frontal affinity chromatography. Biochim. Biophys. Acta 2002, 1572, 232–254. [Google Scholar] [CrossRef]
- Chabot, S.; Kashio, Y.; Seki, M.; Shirato, Y.; Nakamura, K.; Nishi, N.; Nakamura, T.; Matsumoto, R.; Hirashima, M. Regulation of galectin-9 expression and release in Jurkat T cell line cells. Glycobiology 2002, 12, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahfeld, P.; Withers, S.G. Toward universal donor blood: Enzymatic conversion of A and B to O type. J. Biol. Chem. 2020, 295, 325–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, S.M.; Tuzikov, A.B.; Bovin, N.V. Kode Technology Illustrated Technical Manual, 1st ed.; Kode Biotech: Auckland, New Zeland, 2023; pp. 1–80. Available online: https://hdl.handle.net/2292/62953 (accessed on 21 February 2023).
- Lahm, H.; André, S.; Hoeflich, A.; Kaltner, H.; Siebert, H.C.; Sordat, B.; von der Lieth, C.W.; Wolf, E.; Gabius, H.J. Tumor galectinology: Insights into the complex network of a family of endogenous lectins. Glycoconj. J. 2004, 20, 227–238. [Google Scholar] [CrossRef]
- Lahm, H.; André, S.; Hoeflich, A.; Fischer, J.R.; Sordat, B.; Kaltner, H.; Wolf, E.; Gabius, H.J. Comprehensive galectin fingerprinting in a panel of 61 human tumor cell lines by RT-PCR and its implications for diagnostic and therapeutic procedures. Cancer Res. Clin. Oncol. 2001, 127, 375–386. [Google Scholar] [CrossRef]
- Rapoport, E.M.; Matveeva, V.K.; Vokhmyanina, O.A.; Belyanchikov, I.M.; Gabius, H.J.; Bovin, N.V. Localization of galectins within glycocalyx. Biochemistry 2018, 83, 727–737. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rapoport, E.M.; Ryzhov, I.M.; Slivka, E.V.; Korchagina, E.Y.; Popova, I.S.; Khaidukov, S.V.; André, S.; Kaltner, H.; Gabius, H.-J.; Henry, S.; et al. Galectin-9 as a Potential Modulator of Lymphocyte Adhesion to Endothelium via Binding to Blood Group H Glycan. Biomolecules 2023, 13, 1166. https://doi.org/10.3390/biom13081166
Rapoport EM, Ryzhov IM, Slivka EV, Korchagina EY, Popova IS, Khaidukov SV, André S, Kaltner H, Gabius H-J, Henry S, et al. Galectin-9 as a Potential Modulator of Lymphocyte Adhesion to Endothelium via Binding to Blood Group H Glycan. Biomolecules. 2023; 13(8):1166. https://doi.org/10.3390/biom13081166
Chicago/Turabian StyleRapoport, Eugenia M., Ivan M. Ryzhov, Ekaterina V. Slivka, Elena Yu. Korchagina, Inna S. Popova, Sergey V. Khaidukov, Sabine André, Herbert Kaltner, Hans-J. Gabius, Stephen Henry, and et al. 2023. "Galectin-9 as a Potential Modulator of Lymphocyte Adhesion to Endothelium via Binding to Blood Group H Glycan" Biomolecules 13, no. 8: 1166. https://doi.org/10.3390/biom13081166
APA StyleRapoport, E. M., Ryzhov, I. M., Slivka, E. V., Korchagina, E. Y., Popova, I. S., Khaidukov, S. V., André, S., Kaltner, H., Gabius, H. -J., Henry, S., & Bovin, N. V. (2023). Galectin-9 as a Potential Modulator of Lymphocyte Adhesion to Endothelium via Binding to Blood Group H Glycan. Biomolecules, 13(8), 1166. https://doi.org/10.3390/biom13081166