
CD69 Signaling in Eosinophils Induces IL-10 Production and Apoptosis via the Erk1/2 and JNK Pathways, Respectively
 ,
,  , , , , , , and
, , , , , , and 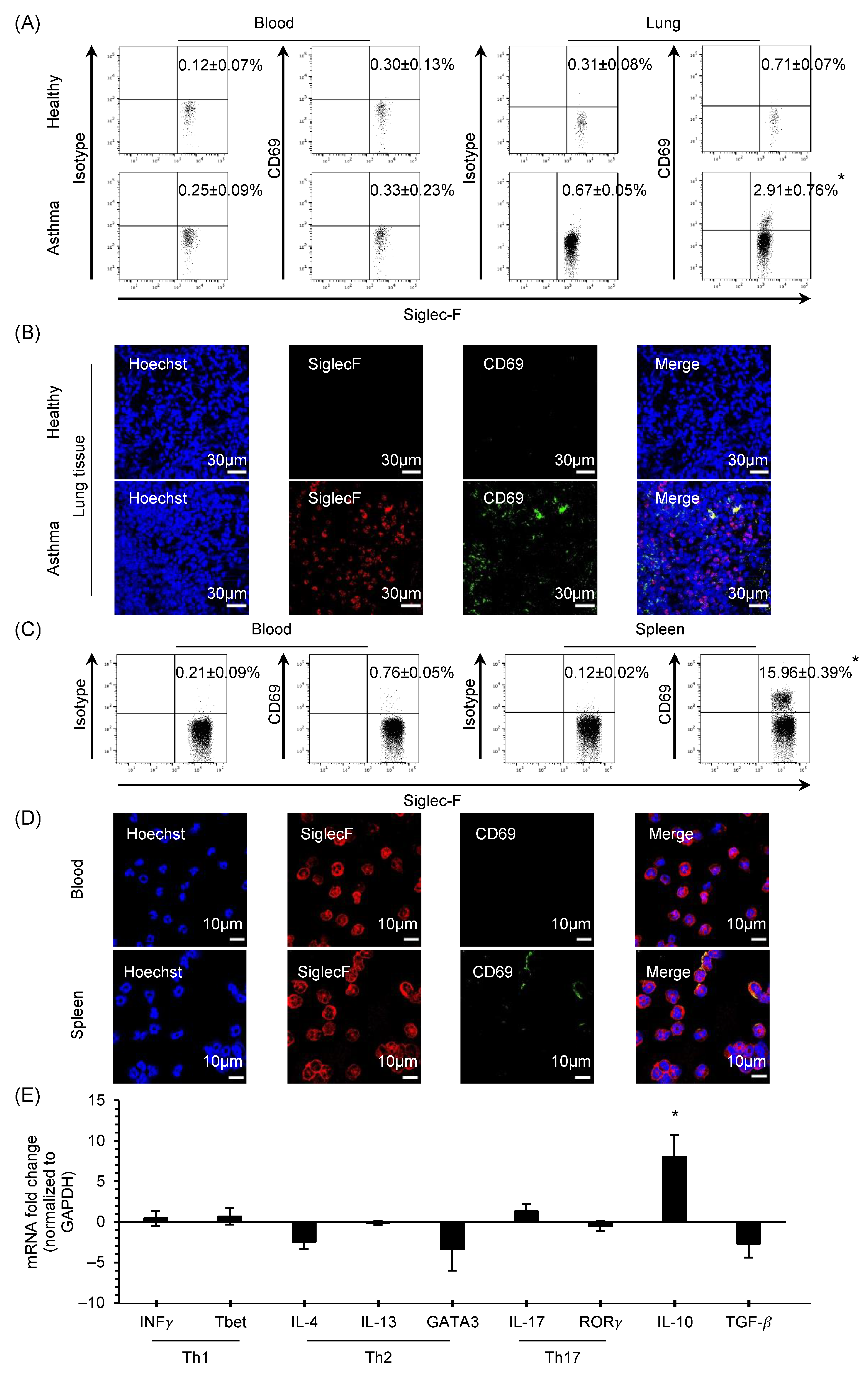{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Animals
2.2. OVA-Induced Asthma Model
2.3. Eosinophil Purification
2.4. CD69 Crosslinking on Eosinophils
2.5. Functional Assay of Eosinophils
2.6. Singal Transduction
2.7. Statistical Analysis
3. Results
3.1. CD69 Expression and Characteristics of CD69+ Eosinophils
3.2. Cytokine Profiling by CD69 Crosslinking on Eosinophils
3.3. Phosphorylation of Erk1/2 and JNK by CD69 Crosslinking on Eosinophils
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Novosad, J.; Krčmová, I.; Souček, O.; Drahošová, M.; Sedlák, V.; Kulířová, M.; Králíčková, P. Subsets of Eosinophils in Asthma, a Challenge for Precise Treatment. Int. J. Mol. Sci. 2023, 24, 5716. [Google Scholar] [CrossRef]
- Frøssing, L.; Von Bülow, A.; Porsbjerg, C. Bronchiectasis in severe asthma is associated with eosinophilic airway inflammation and activation. J. Allergy Clin. Immunol. Glob. 2023, 2, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Al-Shaikhly, T.; Murphy, R.C.; Parker, A.; Lai, Y.; Altman, M.C.; Larmore, M.; Altemeier, W.A.; Frevert, C.W.; Debley, J.S.; Piliponsky, A.M.; et al. Location of eosinophils in the airway wall is critical for specific features of airway hyperresponsiveness and T2 inflammation in asthma. Eur. Respir. J. 2022, 60, 2101865. [Google Scholar] [CrossRef]
- Jesenak, M.; Diamant, Z.; Simon, D.; Tufvesson, E.; Seys, S.F.; Mukherjee, M.; Lacy, P.; Vijverberg, S.; Slisz, T.; Sediva, A.; et al. Eosinophils—From cradle to grave. Allergy 2023, 78, 3077–3102. [Google Scholar] [CrossRef]
- Rothenberg, M.E.; Hogan, S.P. The eosinophil. Annu. Rev. Immunol. 2006, 24, 147–174. [Google Scholar] [CrossRef] [PubMed]
- Kanda, A.; Yun, Y.; Bui, D.V.; Nguyen, L.M.; Kobayashi, Y.; Suzuki, K.; Mitani, A.; Sawada, S.; Hamada, S.; Asako, M.; et al. The multiple functions and subpopulations of eosinophils in tissues under steady-state and pathological conditions. Allergol. Int. 2020, 70, 9–18. [Google Scholar] [CrossRef]
- Rosenberg, H.F.; Dyer, K.D.; Foster, P.S. Eosinophils: Changing perspectives in health and disease. Nat. Rev. Immunol. 2013, 13, 9–22. [Google Scholar] [CrossRef]
- Metcalfe, D.D.; Pawankar, R.; Ackerman, S.J.; Akin, C.; Clayton, F.; Falcone, F.H.; Gleich, G.J.; Irani, A.M.; Johansson, M.W.; Klion, A.D.; et al. Biomarkers of the involvement of mast cells, basophils and eosinophils in asthma and allergic diseases. World Allergy Organ. J. 2016, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Cibrian, D.; Sanchez-Madrid, F. CD69: From activation marker to metabolic gatekeeper. Eur. J. Immunol. 2017, 47, 946–953. [Google Scholar] [CrossRef]
- Yun, Y.; Kanda, A.; Kobayashi, Y.; Van Bui, D.; Suzuki, K.; Sawada, S.; Baba, K.; Yagi, M.; Asako, M.; Okazaki, H.; et al. Increased CD69 expression on activated eosinophils in eosinophilic chronic rhinosinusitis correlates with clinical findings. Allergol. Int. 2020, 69, 232–238. [Google Scholar] [CrossRef]
- Martín, P.; Gómez, M.; Lamana, A.; Matesanz Marín, A.; Cortés, J.R.; Ramírez-Huesca, M.; Barreiro, O.; López-Romero, P.; Gutiérrez-Vázquez, C.; de la Fuente, H.; et al. The leukocyte activation antigen CD69 limits allergic asthma and skin contact hypersensitivity. J. Allergy Clin. Immunol. 2010, 126, 355–365.e3. [Google Scholar] [CrossRef]
- Kobayashi, T.; Iijima, K.; Matsumoto, K.; Lama, J.K.; Kita, H. Lung-resident CD69+ST2+ TH2 cells mediate long-term type 2 memory to inhaled antigen in mice. J. Allergy Clin. Immunol. 2023, 152, 167–181.e6. [Google Scholar] [CrossRef]
- Sasano, H.; Harada, N.; Harada, S.; Takeshige, T.; Sandhu, Y.; Tanabe, Y.; Ishimori, A.; Matsuno, K.; Nagaoka, T.; Ito, J.; et al. Pretreatment circulating MAIT cells, neutrophils, and periostin predicted the real-world response after 1-year mepolizumab treatment in asthmatics. Allergol. Int. 2023, 73, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ochkur, S.I.; McGarry, M.P.; O’Neill, K.R.; Colbert, D.C.; Cormier, S.A.; Lee, N.A.; Shen, H.H.; Lee, J.J. CD69 expression on eosinophils is a marker of activation in the lung following allergen provocation. J. Allergy Clin. Immunol. 2004, 113, S188. [Google Scholar] [CrossRef]
- Chu, H.H.; Kobayashi, Y.; Bui, D.V.; Yun, Y.; Nguyen, L.M.; Mitani, A.; Suzuki, K.; Asako, M.; Kanda, A.; Iwai, H. CCL4 Regulates Eosinophil Activation in Eosinophilic Airway Inflammation. Int. J. Mol. Sci. 2022, 23, 16149. [Google Scholar] [CrossRef] [PubMed]
- Testi, R.; Phillips, J.H.; Lanier, L.L. T cell activation via Leu-23 (CD69). J. Immunol. 1989, 143, 1123–1128. [Google Scholar] [CrossRef] [PubMed]
- Moretta, A.; Poggi, A.; Pende, D.; Tripodi, G.; Orengo, A.M.; Pella, N.; Augugliaro, R.; Bottino, C.; Ciccone, E.; Moretta, L. CD69-mediated pathway of lymphocyte activation: Anti-CD69 monoclonal antibodies trigger the cytolytic activity of different lymphoid effector cells with the exception of cytolytic T lymphocytes expressing T cell receptor alpha/beta. J. Exp. Med. 1991, 174, 1393–1398. [Google Scholar] [CrossRef]
- Martin, P.; Gomez, M.; Lamana, A.; Cruz-Adalia, A.; Ramirez-Huesca, M.; Ursa, M.A.; Yanez-Mo, M.; Sanchez-Madrid, F. CD69 association with Jak3/Stat5 proteins regulates Th17 cell differentiation. Mol. Cell. Biol. 2010, 30, 4877–4889. [Google Scholar] [CrossRef]
- Zingoni, A.; Palmieri, G.; Morrone, S.; Carretero, M.; Lopez-Botel, M.; Piccoli, M.; Frati, L.; Santoni, A. CD69-triggered ERK activation and functions are negatively regulated by CD94 / NKG2-A inhibitory receptor. Eur. J. Immunol. 2000, 30, 644–651. [Google Scholar] [CrossRef]
- Yu, L.; Yang, F.; Zhang, F.; Guo, D.; Li, L.; Wang, X.; Liang, T.; Wang, J.; Cai, Z.; Jin, H. CD69 enhances immunosuppressive function of regulatory T-cells and attenuates colitis by prompting IL-10 production. Cell Death Dis. 2018, 9, 905. [Google Scholar] [CrossRef]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, W.; Rutz, S.; Crellin, N.K.; Valdez, P.A.; Hymowitz, S.G. Regulation and functions of the IL-10 family of cytokines in inflammation and disease. Annu. Rev. Immunol. 2011, 29, 71–109. [Google Scholar] [CrossRef] [PubMed]
- Boonpiyathad, T.; Satitsuksanoa, P.; Akdis, M.; Akdis, C.A. Il-10 producing T and B cells in allergy. Semin. Immunol. 2019, 44, 101326. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; O’Garra, A. The regulation of IL-10 production by immune cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; Christensen, J.R.; Veldhoen, M.; Murphy, T.L.; Murphy, K.M.; O’Garra, A. Interleukin-10 production by Th1 cells requires interleukin-12-induced STAT4 transcription factor and ERK MAP kinase activation by high antigen dose. Immunity 2009, 31, 209–219. [Google Scholar] [CrossRef]
- Woerly, G.; Roger, N.; Loiseau, S.; Dombrowicz, D.; Capron, A.; Capron, M. Expression of CD28 and CD86 by human eosinophils and role in the secretion of type 1 cytokines (interleukin 2 and interferon gamma): Inhibition by immunoglobulin a complexes. J. Exp. Med. 1999, 190, 487–495. [Google Scholar] [CrossRef]
- Walsh, G.M.; Williamson, M.L.; Symon, F.A.; Willars, G.B.; Wardlaw, A.J. Ligation of CD69 induces apoptosis and cell death in human eosinophils cultured with granulocyte-macrophage colony-stimulating factor. Blood 1996, 87, 2815–2821. [Google Scholar] [CrossRef]
- Ickrath, P.; Kleinsasser, N.; Ding, X.; Ginzkey, C.; Beyersdorf, N.; Hagen, R.; Kerkau, T.; Hackenberg, S. Accumulation of CD69+ tissueresident memory T cells in the nasal polyps of patients with chronic rhinosinusitis. Int. J. Mol. Med. 2018, 42, 1116–1124. [Google Scholar] [CrossRef]
- Urasaki, T.; Takasaki, J.; Nagasawa, T.; Ninomiya, H. Pivotal role of 5-lipoxygenase in the activation of human eosinophils: Platelet-activating factor and interleukin-5 induce CD69 on eosinophils through the 5-lipoxygenase pathway. J. Leukoc. Biol. 2001, 69, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Gebreselassie, N.G.; Gagliardo, L.F.; Ruyechan, M.C.; Lee, N.A.; Lee, J.J.; Appleton, J.A. Eosinophil-derived IL-10 supports chronic nematode infection. J. Immunol. 2014, 193, 4178–4187. [Google Scholar] [CrossRef]
- Nakajima, H.; Gleich, G.J.; Kita, H. Constitutive production of IL-4 and IL-10 and stimulated production of IL-8 by normal peripheral blood eosinophils. J. Immunol. 1996, 156, 4859–4866. [Google Scholar] [CrossRef]
- Foerster, M.; Haefner, D.; Kroegel, C. Bcl-2-mediated regulation of CD69-induced apoptosis of human eosinophils: Identification and characterization of a novel receptor-induced mechanism and relationship to CD95-transduced signalling. Scand. J. Immunol. 2002, 56, 417–428. [Google Scholar] [CrossRef]
- Gavioli, R.; Risso, A.; Smilovich, D.; Baldissarro, I.; Capra, M.C.; Bargellesi, A.; Cosulich, M.E. CD69 molecule in human neutrophils: Its expression and role in signal-transducing mechanisms. Cell. Immunol. 1992, 142, 186–196. [Google Scholar] [CrossRef]
- de la Fuente, H.; Cruz-Adalia, A.; Martinez Del Hoyo, G.; Cibrian-Vera, D.; Bonay, P.; Perez-Hernandez, D.; Vazquez, J.; Navarro, P.; Gutierrez-Gallego, R.; Ramirez-Huesca, M.; et al. The leukocyte activation receptor CD69 controls T cell differentiation through its interaction with galectin-1. Mol. Cell. Biol. 2014, 34, 2479–2487. [Google Scholar] [CrossRef]
- Hayashizaki, K.; Kimura, M.Y.; Tokoyoda, K.; Hosokawa, H.; Shinoda, K.; Hirahara, K.; Ichikawa, T.; Onodera, A.; Hanazawa, A.; Iwamura, C.; et al. Myosin light chains 9 and 12 are functional ligands for CD69 that regulate airway inflammation. Sci. Immunol. 2016, 1, eaaf9154. [Google Scholar] [CrossRef] [PubMed]
- van der Leij, J.; van den Berg, A.; Blokzijl, T.; Harms, G.; van Goor, H.; Zwiers, P.; van Weeghel, R.; Poppema, S.; Visser, L. Dimeric galectin-1 induces IL-10 production in T-lymphocytes: An important tool in the regulation of the immune response. J. Pathol. 2004, 204, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.Y.; Hayashizaki, K.; Tokoyoda, K.; Takamura, S.; Motohashi, S.; Nakayama, T. Crucial role for CD69 in allergic inflammatory responses: CD69-Myl9 system in the pathogenesis of airway inflammation. Immunol. Rev. 2017, 278, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, M.; Kimura, M.Y.; Ito, T.; Hayashizaki, K.; Endo, Y.; Wang, Y.; Yagi, R.; Nakagawa, T.; Kato, N.; Matsubara, H.; et al. Myosin Light Chain 9/12 Regulates the Pathogenesis of Inflammatory Bowel Disease. Front. Immunol. 2020, 11, 594297. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bui, D.V.; Nguyen, L.M.; Kanda, A.; Chu, H.H.; Thi Le, N.K.; Yun, Y.; Kobayashi, Y.; Suzuki, K.; Mitani, A.; Shimamura, A.; et al. CD69 Signaling in Eosinophils Induces IL-10 Production and Apoptosis via the Erk1/2 and JNK Pathways, Respectively. Biomolecules 2024, 14, 360. https://doi.org/10.3390/biom14030360
Bui DV, Nguyen LM, Kanda A, Chu HH, Thi Le NK, Yun Y, Kobayashi Y, Suzuki K, Mitani A, Shimamura A, et al. CD69 Signaling in Eosinophils Induces IL-10 Production and Apoptosis via the Erk1/2 and JNK Pathways, Respectively. Biomolecules. 2024; 14(3):360. https://doi.org/10.3390/biom14030360
Chicago/Turabian StyleBui, Dan Van, Linh Manh Nguyen, Akira Kanda, Hanh Hong Chu, Nhi Kieu Thi Le, Yasutaka Yun, Yoshiki Kobayashi, Kensuke Suzuki, Akitoshi Mitani, Akihiro Shimamura, and et al. 2024. "CD69 Signaling in Eosinophils Induces IL-10 Production and Apoptosis via the Erk1/2 and JNK Pathways, Respectively" Biomolecules 14, no. 3: 360. https://doi.org/10.3390/biom14030360
APA StyleBui, D. V., Nguyen, L. M., Kanda, A., Chu, H. H., Thi Le, N. K., Yun, Y., Kobayashi, Y., Suzuki, K., Mitani, A., Shimamura, A., Fukui, K., Sawada, S., Dombrowicz, D., & Iwai, H. (2024). CD69 Signaling in Eosinophils Induces IL-10 Production and Apoptosis via the Erk1/2 and JNK Pathways, Respectively. Biomolecules, 14(3), 360. https://doi.org/10.3390/biom14030360