
CPSF30 at the Interface of Alternative Polyadenylation and Cellular Signaling in Plants

Abstract
:1. Introduction

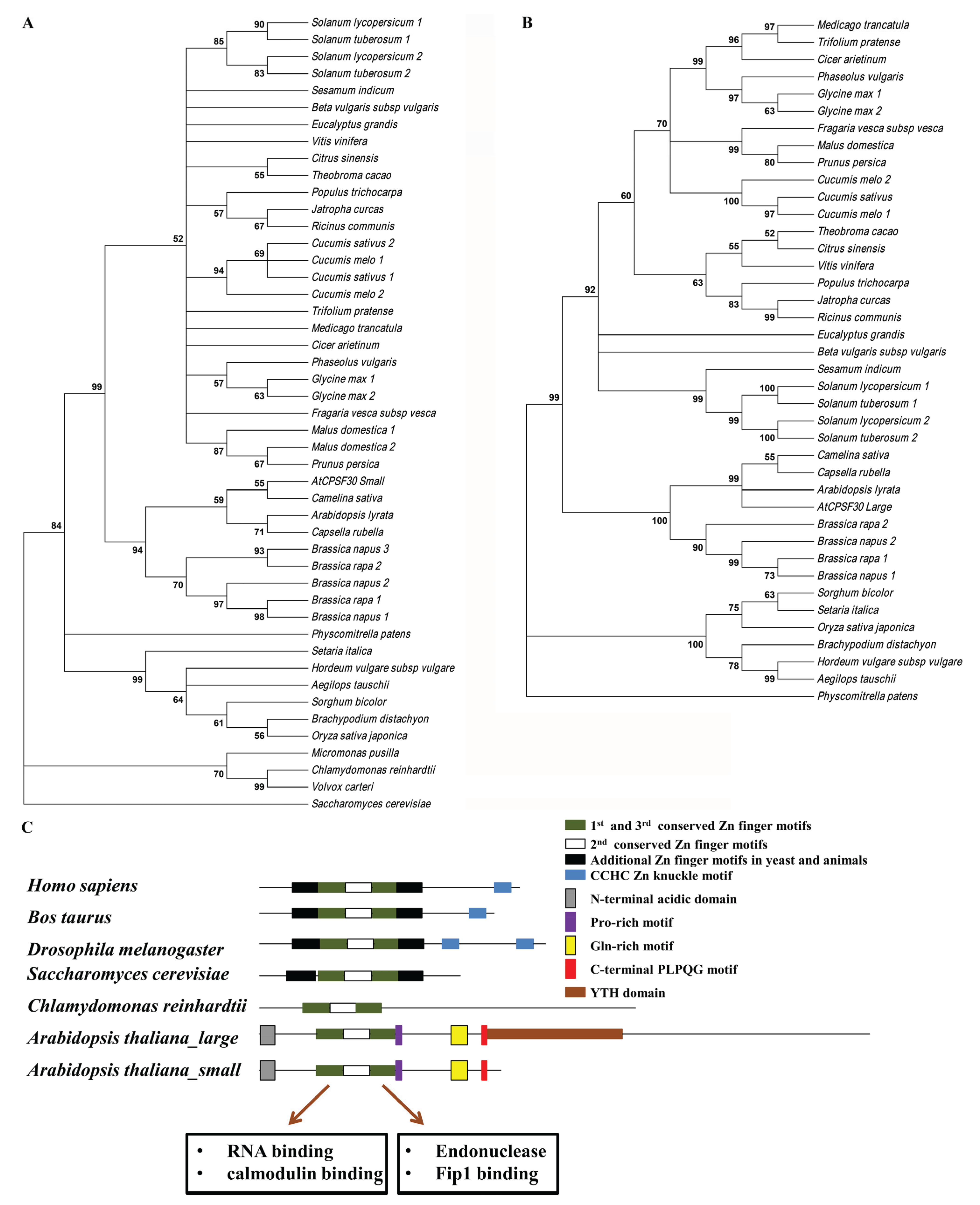
2. CPSF30 in Plant is a Key Polyadenylation Factor with Unique Biochemical Features

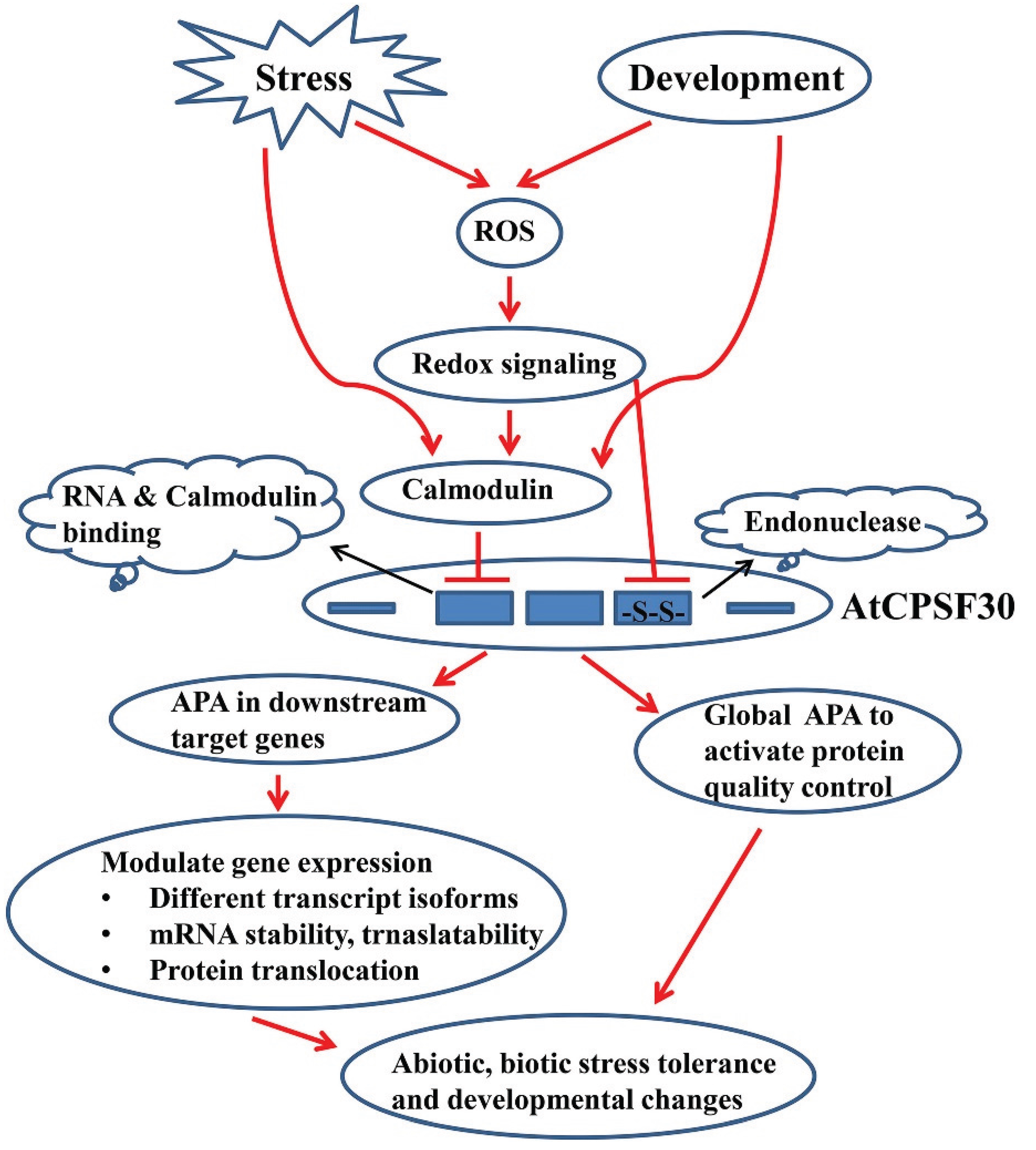
3. CPSF30, Cellular Signaling, and Plant Growth and Development

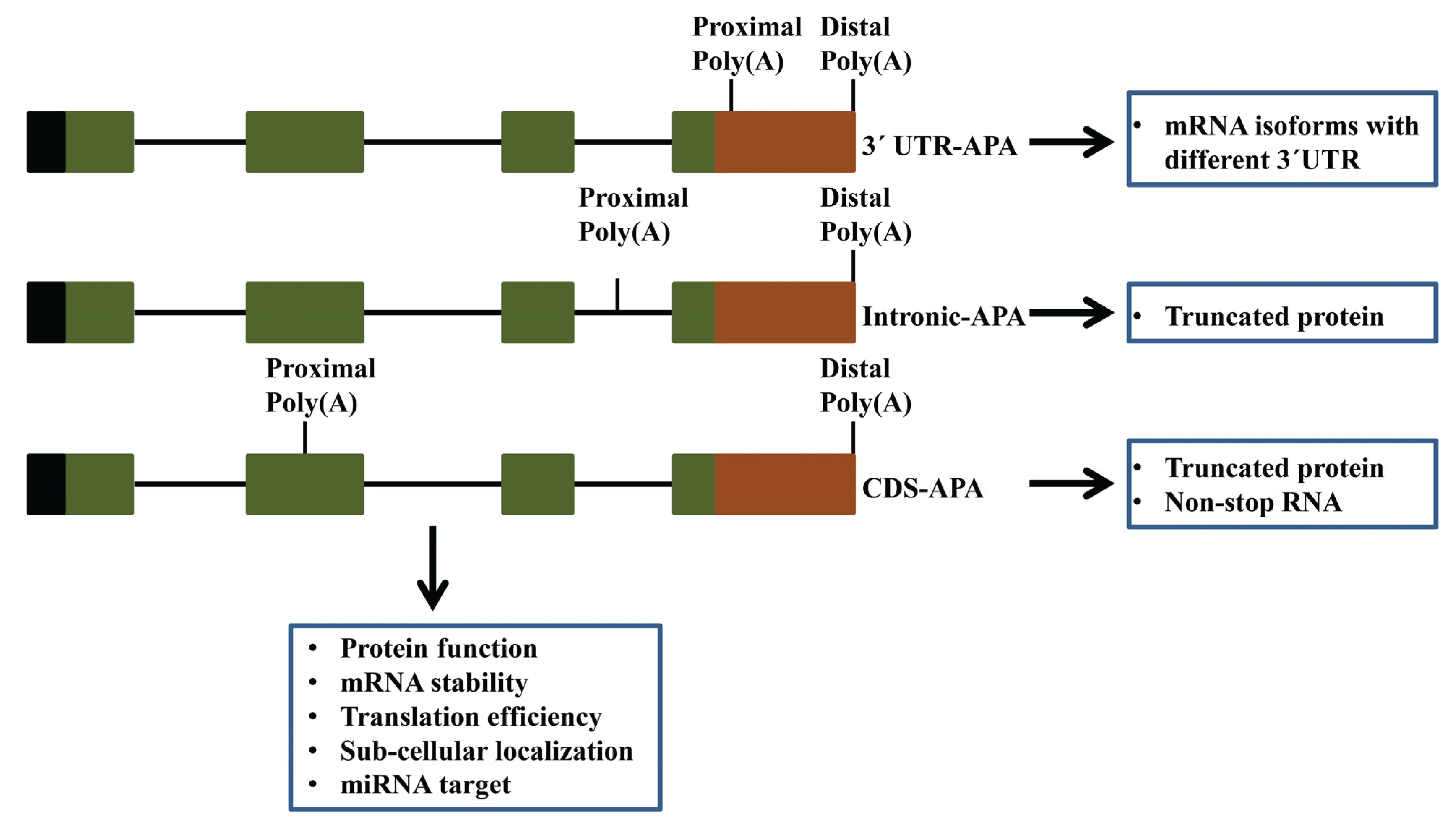
4. The Scope of AtCPSF30-Mediated APA
5. Conclusions and Future Directions
Acknowledgments
Author Contributions
Appendix

{kind=link}
{kind=link}
{kind=link}
| Species | Genbank accessions used for the phylogeny for small polypeptide | Genbank accessions used for the phylogeny for large polypeptide |
|---|---|---|
| Camelina sativa | XP_010460838 | XP_010478453 |
| Brassica napus | CDX90251, CDX98205, CDX77591 | CDX77591, CDX98205 |
| Brassica rapa | XP_009102697, XP_009109585 | XP_009109585, XP_009102697 |
| Populus trichocarpa | XP_006377637 | XP_002300333 |
| Arabidopsis lyrata | XP_002893618 | XP_002893618 |
| Capsella rubella | XP_006306994 | XP_006306994 |
| Cucumis sativus | XP_004156192, XP_004141524 | XP_004141524 |
| Cucumis melo | XP_008459517, XP_008445183 | XP_008459517, XP_008445183 |
| Citrus sinensis | XP_006468290 | XP_006468290 |
| Vitis vinifera | XP_002281594 | XP_002281594 |
| Malus domestica | NP_001280880, XP_008372260 | NP_001280880 |
| Theobroma cacao | XP_007041140 | XP_007041140 |
| Medicago trancatula | KEH37048 | KEH37048 |
| Cicer arietinum | XP_004486563 | XP_004486563 |
| Beta vulgaris subsp vulgaris | XP_010687042 | XP_010687042 |
| Solanum lycopersicum | XP_004231555, XP_004233145 | XP_004231555, XP_004233145 |
| Solanum tuberosum | XP_006359103, XP_006352991 | XP_006359103, XP_006352991 |
| Trifolium pratense | Predicted from RNAseq | Predicted from RNAseq |
| Fragaria vesca subsp vesca | XP_004295608 | XP_004295608 |
| Hordeum vulgare subsp vulgare | BAJ96745 | BAJ96745 |
| Sorghum bicolor | XP_002437445 | XP_002437445 |
| Oryza sativa japonica | NP_001058359 | NP_001058359 |
| Brachypodium distachyon | XP_003563404 | XP_003563404 |
| Phaseolus vulgaris | XP_007147504 | XP_007147504 |
| Glycine max | XP_003546247, XP_003534764 | XP_003546247, XP_003534764 |
| Jatropha curcas | KDP34942 | KDP34942 |
| Ricinus communis | XP_002523201 | XP_002523201 |
| Prunus persica | XP_007214175 | XP_007214175 |
| Eucalyptus grandis | XP_010056977 | XP_010056977 |
| Setaria italica | XP_004966206 | XP_004966206 |
| Sesamum indicum | XP_011085214 | XP_011085214 |
| Aegilops tauschii | EMT09537 | EMT09537 |
| Physcomitrella patens | XP_001753463 | XP_001753463 |
| Chlamydomonas reinhardtii | g18261.t1 (from Phytozome) | |
| Volvox carteri | XP_002947795 | |
| Micromonas pusilla | XP_003056614 | |
| Saccharomyces cerevisiae_EC1118 | CAY87059 |
Conflicts of Interest
References
- Gruber, A.R.; Martin, G.; Keller, W.; Zavolan, M. Means to an end: Mechanisms of alternative polyadenylation of messenger RNA precursors. Wiley Interdiscip. Rev. RNA 2014, 5, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Neve, J.; Furger, A. Alternative polyadenylation: Less than meets the eye? Biochem. Soc. Trans. 2014, 42, 1190–1195. [Google Scholar] [CrossRef] [PubMed]
- Elkon, R.; Ugalde, A.P.; Agami, R. Alternative cleavage and polyadenylation: Extent, regulation and function. Nat. Rev. Genet. 2013, 14, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Manley, J.L. Alternative cleavage and polyadenylation: The long and short of it. Trends Biochem. Sci. 2013, 38, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Bentley, D.L. Rules of engagement: Co-transcriptional recruitment of pre-mRNA processing factors. Curr. Opin. Cell Biol. 2005, 17, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Erickson, B.; Luo, W.; Seward, D.; Graber, J.H.; Pollock, D.D.; Megee, P.C.; Bentley, D.L. Gene-specific RNA polymerase II phosphorylation and the CTD code. Nat. Struct. Mol. Biol. 2010, 17, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Lemay, J.-F.; Lemieux, C.; St-Andre, O.; Bachand, F. Crossing the borders poly(A)-binding proteins working on both sides of the fence. RNA Biol. 2010, 7, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Mapendano, C.K.; Lykke-Andersen, S.; Kjems, J.; Bertrand, E.; Jensen, T.H. Crosstalk between mRNA 3' end processing and transcription initiation. Mol. Cell 2010, 40, 410–422. [Google Scholar] [CrossRef] [PubMed]
- Ozsolak, F.; Kapranov, P.; Foissac, S.; Kim, S.W.; Fishilevich, E.; Monaghan, A.P.; John, B.; Milos, P.M. Comprehensive polyadenylation site maps in yeast and human reveal pervasive alternative polyadenylation. Cell 2010, 143, 1018–1029. [Google Scholar] [CrossRef] [PubMed]
- Derti, A.; Garrett-Engele, P.; MacIsaac, K.D.; Stevens, R.C.; Sriram, S.; Chen, R.H.; Rohl, C.A.; Johnson, J.M.; Babak, T. A quantitative atlas of polyadenylation in five mammals. Genome Res. 2012, 22, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Hu, J.; Zhang, H.B.; Lutz, C.S. A large-scale analysis of mRNA polyadenylation of human and mouse genes. Nucleic Acids Res. 2005, 33, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.H.; Liu, M.; Downie, B.; Liang, C.; Ji, G.L.; Li, Q.Q.; Hunt, A.G. Genome-wide landscape of polyadenylation in Arabidopsis provides evidence for extensive alternative polyadenylation. Proc. Natl. Acad. Sci. USA 2011, 108, 12533–12538. [Google Scholar] [CrossRef] [PubMed]
- Shepard, P.J.; Choi, E.A.; Lu, J.T.; Flanagan, L.A.; Hertel, K.J.; Shi, Y.S. Complex and dynamic landscape of RNA polyadenylation revealed by PAS-Seq. RNA 2011, 17, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y. Alternative polyadenylation: New insights from global analyses. RNA 2012, 18, 2105–2117. [Google Scholar] [CrossRef] [PubMed]
- Boutet, S.C.; Cheung, T.H.; Quach, N.L.; Liu, L.; Prescott, S.L.; Edalati, A.; Iori, K.; Rando, T.A. Alternative polyadenylation mediates microrna regulation of muscle stem cell function. Cell Stem Cell 2012, 10, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Kreth, S.; Limbeck, E.; Hinske, L.C.; Schutz, S.V.; Thon, N.; Hoefig, K.; Egensperger, R.; Kreth, F.W. In human glioblastomas transcript elongation by alternative polyadenylation and mirna targeting is a potent mechanism of mgmt silencing. Acta Neuropathol. 2013, 125, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.; McKenna, M.C.; Rollins, D.A.; Song, M.; Nuriel, T.; Gross, S.S.; Xu, G.Q.; Glatt, C.E. Anxiety-associated alternative polyadenylation of the serotonin transporter mRNA confers translational regulation by hnrnpk. Proc. Natl. Acad. Sci. USA 2013, 110, 11624–11629. [Google Scholar] [CrossRef] [PubMed]
- Tranter, M.; Helsley, R.N.; Paulding, W.R.; McGuinness, M.; Brokamp, C.; Haar, L.; Liu, Y.; Ren, X.P.; Jones, W.K. Coordinated post-transcriptional regulation of Hsp70.3 gene expression by microrna and alternative polyadenylation. J. Biol. Chem. 2011, 286, 29828–29837. [Google Scholar] [CrossRef] [PubMed]
- Pinto, P.A.B.; Henriques, T.; Freitas, M.O.; Martins, T.; Domingues, R.G.; Wyrzykowska, P.S.; Coelho, P.A.; Carmo, A.M.; Sunkel, C.E.; Proudfoot, N.J.; et al. RNA polymerase II kinetics in polo polyadenylation signal selection. EMBO J. 2011, 30, 2431–2444. [Google Scholar] [CrossRef] [PubMed]
- Akman, B.H.; Can, T.; Erson-Bensan, A.E. Estrogen-induced upregulation and 3'-UTR shortening of CDC6. Nucleic Acids Res. 2012, 40, 10679–10688. [Google Scholar] [CrossRef] [PubMed]
- Miura, P.; Shenker, S.; Andreu-Agullo, C.; Westholm, J.O.; Lai, E.C. Widespread and extensive lengthening of 3' UTRs in the mammalian brain. Genome Res. 2013, 23, 812–825. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L.; DeCerbo, J.N.; Carmichael, G.G. Alu element-mediated gene silencing. EMBO J. 2008, 27, 1694–1705. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Brockman, J.M.; Dass, B.; Hutchins, L.N.; Singh, P.; McCarrey, J.R.; MacDonald, C.C.; Graber, J.H. Systematic variation in mRNA 3'-processing signals during mouse spermatogenesis. Nucleic Acids Res. 2007, 35, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Lee, J.Y.; Pan, Z.; Jiang, B.; Tian, B. Progressive lengthening of 3' untranslated regions of mRNAs by alternative polyadenylation during mouse embryonic development. Proc. Natl. Acad. Sci. USA 2009, 106, 7028–7033. [Google Scholar] [CrossRef] [PubMed]
- Mayr, C.; Bartel, D.P. Widespread shortening of 3' UTRs by alternative cleavage and polyadenylation activates oncogenes in cancer cells. Cell 2009, 138, 673–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orkin, S.H.; Cheng, T.C.; Antonarakis, S.E.; Kazazian, H.H. Thalassemia due to a mutation in the cleavage-polyadenylation signal of the human beta-globin gene. EMBO J. 1985, 4, 453–456. [Google Scholar] [PubMed]
- Gehring, N.H.; Frede, U.; Neu-Yilik, G.; Hundsdoerfer, P.; Vetter, B.; Hentze, M.W.; Kulozik, A.E. Increased efficiency of mRNA 3' end formation: A new genetic mechanism contributing to hereditary thrombophilia. Nat. Genet. 2001, 28, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Higgs, D.R.; Goodbourn, S.E.Y.; Lamb, J.; Clegg, J.B.; Weatherall, D.J.; Proudfoot, N.J. α-Thalassemia caused by a polyadenylation signal mutation. Nature 1983, 306, 398–400. [Google Scholar] [CrossRef] [PubMed]
- Brais, B.; Bouchard, J.P.; Xie, Y.G.; Rochefort, D.L.; Chretien, N.; Tome, F.M.S.; Lafreniere, R.G.; Rommens, J.M.; Uyama, E.; Nohira, O.; et al. Short GCG expansions in the PABP2 gene cause oculopharyngeal muscular dystrophy. Nat. Genet. 1998, 18, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.L.; Brunkow, M.E.; Ramsdell, F.; O’Briant, K.C.; Zhu, Q.; Fuleihan, R.L.; Shigeoka, A.O.; Ochs, H.D.; Chance, P.F. A rare polyadenylation signal mutation of the FOXP3 gene (AAUAAA → AAUGAA) leads to the IPEX syndrome. Immunogenetics 2001, 53, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.L.G.; Bristol, L.A.; Jin, L.; Dykes-Hoberg, M.; Crawford, T.; Clawson, L.; Rothstein, J.D. Aberrant RNA processing in a neurodegenerative disease: The cause for absent EAAT2 a glutamate transporter, in amyotrophic lateral sclerosis. Neuron 1998, 20, 589–602. [Google Scholar] [CrossRef]
- Lackford, B.; Yao, C.G.; Charles, G.M.; Weng, L.J.; Zheng, X.F.; Choi, E.A.; Xie, X.H.; Wan, J.; Xing, Y.; Freudenberg, J.M.; et al. Fip1 regulates mRNA alternative polyadenylation to promote stem cell self-renewal. EMBO J. 2014, 33, 878–889. [Google Scholar] [CrossRef] [PubMed]
- Simpson, G.G.; Dijkwel, P.P.; Quesada, V.; Henderson, I.; Dean, C. Fy is an RNA 3' end-processing factor that interacts with FCA to control the Arabidopsis floral transition. Cell 2003, 113, 777–787. [Google Scholar] [CrossRef]
- Liu, F.; Marquardt, S.; Lister, C.; Swiezewski, S.; Dean, C. Targeted 3' processing of antisense transcripts triggers Arabidopsis FLC chromatin silencing. Science 2010, 327, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Hornyik, C.; Terzi, L.C.; Simpson, G.G. The spen family protein FPA controls alternative cleavage and polyadenylation of RNA. Dev. Cell 2010, 18, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Duc, C.; Sherstnev, A.; Cole, C.; Barton, G.J.; Simpson, G.G. Transcription termination and chimeric RNA formation controlled by Arabidopsis thaliana FPA. PLoS Genet. 2013. [Google Scholar] [CrossRef]
- Xu, R.Q.; Ye, X.F.; Li, Q.S.Q. AtCPSF73-II gene encoding an Arabidopsis homolog of CPSF 73 kDa subunit is critical for early embryo development. Gene 2004, 324, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.Q.; Zhao, H.W.; Dinkins, R.D.; Cheng, X.W.; Carberry, G.; Li, Q.Q. The 73 kD subunit of the cleavage and polyadenylation specificity factor (CPSF) complex affects reproductive development in Arabidopsis. Plant Mol. Biol. 2006, 61, 799–815. [Google Scholar] [CrossRef] [PubMed]
- Giranton, J.L.; Ariza, M.J.; Dumas, C.; Cock, J.M.; Gaude, T. The S locus receptor kinase gene encodes a soluble glycoprotein corresponding to the SKR extracellular domain in Brassica oleracea. Plant J. 1995, 8, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Tantikanjana, T.; Nasrallah, M.E.; Stein, J.C.; Chen, C.H.; Nasrallah, J.B. An alternative transcript of the S locus glycoprotein gene in a class II pollen-recessive self-incompatibility haplotype of Brassica oleracea encodes a membrane-anchored protein. Plant Cell 1993, 5, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Zhu, X.; Gakiere, B.; Levanony, H.; Kahana, A.; Galili, G. The bifunctional LKR/SDH locus of plants also encodes a highly active monofunctional lysine-ketoglutarate reductase using a polyadenylation signal located within an intron. Plant Physiol. 2002, 130, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Moody, M.A.; Nassuth, A. Grape contains 4 ICE genes whose expression includes alternative polyadenylation, leading to transcripts encoding at least 7 different ICE proteins. Environ. Exp. Bot. 2014, 106, 70–78. [Google Scholar] [CrossRef]
- Wu, X.; Gaffney, B.; Hunt, A.G.; Li, Q.Q. Genome-wide determination of poly(A) sites in medicago truncatula: Evolutionary conservation of alternative poly(A) site choice. BMC Genomics 2014. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Venu, R.C.; Nobuta, K.; Wu, X.; Notibala, V.; Demirci, C.; Meyers, B.C.; Wang, G.L.; Ji, G.; Li, Q.Q. Transcriptome dynamics through alternative polyadenylation in developmental and environmental responses in plants revealed by deep sequencing. Genome Res. 2011, 21, 1478–1486. [Google Scholar] [CrossRef] [PubMed]
- Sherstnev, A.; Duc, C.; Cole, C.; Zacharaki, V.; Hornyik, C.; Ozsolak, F.; Milos, P.M.; Barton, G.J.; Simpson, G.G. Direct sequencing of Arabidopsis thaliana RNA reveals patterns of cleavage and polyadenylation. Nat. Struct. Mol. Biol. 2012, 19, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Mandel, C.R.; Bai, Y.; Tong, L. Protein factors in pre-mRNA 3'-end processing. Cell Mol. Life Sci. 2008, 65, 1099–1122. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; di Giammartino, D.C.; Taylor, D.; Sarkeshik, A.; Rice, W.J.; Yates, J.R.; Frank, J.; Manley, J.L. Molecular architecture of the human pre-mRNA 3' processing complex. Mol. Cell 2009, 33, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Hunt, A.G.; Xing, D.; Li, Q.Q. Plant polyadenylation factors: Conservation and variety in the polyadenylation complex in plants. BMC Genomics 2012. [Google Scholar] [CrossRef] [PubMed]
- Nemeroff, M.E.; Barabino, S.M.L.; Li, Y.Z.; Keller, W.; Krug, R.M. Influenza virus NS1 protein interacts with the cellular 30 kDa subunit of CPSF and inhibits 3' end formation of cellular pre-mRNAs. Mol. Cell 1998, 1, 991–1000. [Google Scholar] [CrossRef]
- Chan, S.L.; Huppertz, I.; Yao, C.; Weng, L.; Moresco, J.J.; Yates, J.R., III; Ule, J.; Manley, J.L.; Shi, Y. CPSF30 and WDR33 directly bind to aauaaa in mammalian mRNA 3' processing. Genes Dev. 2014, 28, 2370–2380. [Google Scholar] [CrossRef] [PubMed]
- Delaney, K.J.; Xu, R.Q.; Zhang, J.X.; Li, Q.Q.; Yun, K.Y.; Falcone, D.L.; Hunt, A.G. Calmodulin interacts with and regulates the RNA-binding activity of an Arabidopsis polyadenylation factor subunit. Plant Physiol. 2006, 140, 1507–1521. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, Z.; Gomez, A.; Hon, G.C.; Yue, Y.; Han, D.; Fu, Y.; Parisien, M.; Dai, Q.; Jia, G.; et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature 2014, 505, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.Y.; Tolias, P.P. Drosophila clipper/CPSF 30k is a post-transcriptionally regulated nuclear protein that binds RNA containing GC clusters. Nucleic Acids Res. 1998, 26, 1597–1604. [Google Scholar] [CrossRef] [PubMed]
- Addepalli, B.; Hunt, A.G. A novel endonuclease activity associated with the Arabidopsis ortholog of the 30-kDa subunit of cleavage and polyadenylation specificity factor. Nucleic Acids Res. 2007, 35, 4453–4463. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.; Tolias, P.P. Cleavage of RNA hairpins mediated by a developmentally regulated CCCH zinc finger protein. Mol. Cell Biol. 1996, 16, 6661–6667. [Google Scholar] [PubMed]
- Tacahashi, Y.; Helmling, S.; Moore, C.L. Functional dissection of the zinc finger and flanking domains of the YTH1 cleavage/polyadenylation factor. Nucleic Acids Res. 2003, 31, 1744–1752. [Google Scholar] [CrossRef] [PubMed]
- Addepalli, B.; Hunt, A.G. Redox and heavy metal effects on the biochemical activities of an Arabidopsis polyadenylation factor subunit. Arch. Biochem. Biophys. 2008, 473, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Addepalli, B.; Limbach, P.A.; Hunt, A.G. A disulfide linkage in a CCCH zinc finger motif of an Arabidopsis CPSF30 ortholog. FEBS Lett. 2010, 584, 4408–4412. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.A.; Hunt, A.G. The Arabidopsis ortholog of the 77 kDa subunit of the cleavage stimulatory factor (AtCSTF-77) involved in mRNA polyadenylation is an RNA-binding protein. FEBS Lett. 2010, 584, 1449–1454. [Google Scholar] [CrossRef] [PubMed]
- Forbes, K.P.; Addepalli, B.; Hunt, A.G. An Arabidopsis Fip1 homolog interacts with RNA and provides conceptual links with a number of other polyadenylation factor subunits. J. Biol. Chem. 2006, 281, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Hunt, A.G.; Xu, R.Q.; Addepalli, B.; Rao, S.; Forbes, K.P.; Meeks, L.R.; Xing, D.H.; Mo, M.; Zhao, H.W.; Bandyopadhyay, A.; et al. Arabidopsis mRNA polyadenylation machinery: Comprehensive analysis of protein-protein interactions and gene expression profiling. BMC Genomics 2008. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Dinkins, R.D.; Hunt, A.G. Distinctive interactions of the Arabidopsis homolog of the 30 kD subunit of the cleavage and polyadenylation specificity factor (AtCPSF30) with other polyadenylation factor subunits. BMC Cell Biol. 2009. [Google Scholar] [CrossRef] [PubMed]
- Twu, K.Y.; Noah, D.L.; Rao, P.; Kuo, R.L.; Krug, R.M. The CPSF30 binding site on the NS1A protein of influenza a virus is a potential antiviral target. J. Virol. 2006, 80, 3957–3965. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.B.; Guo, W.; Li, M.; Shi, D.B.; Tian, Y.; Li, Z.L.; Wang, J.S.; Fu, L.Y.; Xiao, X.S.; Liu, Q.Q.; et al. Upregulation of cleavage and polyadenylation specific factor 4 in lung adenocarcinoma and its critical role for cancer cell survival and proliferation. PLoS ONE 2013, 8, e82728. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.B.; Qin, L.J.; Wang, S.S.; Li, M.; Shi, D.B.; Tian, Y.; Wang, J.S.; Fu, L.Y.; Li, Z.L.; Guo, W.; et al. CPSF4 activates telomerase reverse transcriptase and predicts poor prognosis in human lung adenocarcinomas. Mol. Oncol. 2014, 8, 704–716. [Google Scholar] [CrossRef] [PubMed]
- Sarwat, M.; Ahmad, P.; Nabi, G.; Hu, X. Ca2+ signals: The versatile decoders of environmental cues. Crit. Rev. Biotechnol. 2013, 33, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Xu, R.Q.; Merrill, C.; Hong, L.W.; von Lanken, C.; Hunt, A.G.; Li, Q.S.Q. Integration of developmental and environmental signals via a polyadenylation factor in Arabidopsis. PLoS ONE 2014, 9, e115779. [Google Scholar] [CrossRef] [PubMed]
- Bruggeman, Q.; Garmier, M.; de Bont, L.; Soubigou-Taconnat, L.; Mazubert, C.; Benhamed, M.; Raynaud, C.; Bergounioux, C.; Delarue, M. The polyadenylation factor subunit cleavage and polyadenylation specificity factor30: A key factor of programmed cell death and a regulator of immunity in Arabidopsis. Plant Physiol. 2014, 165, 732–746. [Google Scholar] [CrossRef] [PubMed]
- Coll, N.S.; Epple, P.; Dangl, J.L. Programmed cell death in the plant immune system. Cell Death Differ. 2011, 18, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Gapper, C.; Dolan, L. Control of plant development by reactive oxygen species. Plant Physiol. 2006, 141, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Vranova, E.; Atichartpongkul, S.; Villarroel, R.; van Montagu, M.; Inze, D.; van Camp, W. Comprehensive analysis of gene expression in nicotiana tabacum leaves acclimated to oxidative stress. Proc. Natl. Acad. Sci. USA 2002, 99, 10870–10875. [Google Scholar] [CrossRef] [PubMed]
- Evans, N.H.; McAinsh, M.R.; Hetherington, A.M.; Knight, M.R. Ros perception in Arabidopsis thaliana: The ozone-induced calcium response. Plant J. 2005, 41, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Price, A.H.; Taylor, A.; Ripley, S.J.; Griffiths, A.; Trewavas, A.J.; Knight, M.R. Oxidative signals in tobacco increase cytosolic calcium. Plant Cell 1994, 6, 1301–1310. [Google Scholar] [CrossRef] [PubMed]
- Rentel, M.C.; Lecourieux, D.; Ouaked, F.; Usher, S.L.; Petersen, L.; Okamoto, H.; Knight, H.; Peck, S.C.; Grierson, C.S.; Hirt, H.; et al. Oxi1 kinase is necessary for oxidative burst-mediated signalling in Arabidopsis. Nature 2004, 427, 858–861. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Poovaiah, B.W. Hydrogen peroxide homeostasis: Activation of plant catalase by calcium/calmodulin. Proc. Natl. Acad. Sci. USA 2002, 99, 4097–4102. [Google Scholar] [CrossRef] [PubMed]
- Barford, D. The role of cysteine residues as redox-sensitive regulatory switches. Curr. Opin. Struct. Biol. 2004, 14, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.E.; Wu, X.H.; Liu, M.; Gaffney, B.; Ji, G.L.; Li, Q.S.Q.; Hunt, A.G. Genome-wide control of polyadenylation site choice by CPSF30 in Arabidopsis. Plant Cell 2012, 24, 4376–4388. [Google Scholar] [CrossRef] [PubMed]
- Loke, J.C.; Stahlberg, E.A.; Strenski, D.G.; Haas, B.J.; Wood, P.C.; Li, Q.Q. Compilation of mRNA polyadenylation signals in Arabidopsis revealed a new signal element and potential secondary structures. Plant Physiol. 2005, 138, 1457–1468. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Addepalli, B.; Yun, K.Y.; Hunt, A.G.; Xu, R.; Rao, S.; Li, Q.Q.; Falcone, D.L. A polyadenylation factor subunit implicated in regulating oxidative signaling in Arabidopsis thaliana. PLoS ONE 2008, 3, e2410. [Google Scholar] [CrossRef] [PubMed]
- Howell, S.H. Endoplasmic reticulum stress responses in plants. Annu. Rev. Plant Biol. 2013, 64, 477–499. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-X.; Howell, S.H. Endoplasmic reticulum protein quality control and its relationship to environmental stress responses in plants. Plant Cell 2010, 22, 2930–2942. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, J. Endoplasmic reticulum-mediated protein quality control in Arabidopsis. Front. Plant Sci. 2014. [Google Scholar] [CrossRef] [PubMed]
- Fanata, W.I.D.; Lee, S.Y.; Lee, K.O. The unfolded protein response in plants: A fundamental adaptive cellular response to internal and external stresses. J. Proteomics 2013, 93, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Clancy, M.J.; Shambaugh, M.E.; Timpte, C.S.; Bokar, J.A. Induction of sporulation in saccharomyces cerevisiae leads to the formation of N6-methyladenosine in mRNA: A potential mechanism for the activity of the IME4 gene. Nucleic Acids Res. 2002, 30, 4509–4518. [Google Scholar] [CrossRef] [PubMed]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6a RNA methylomes revealed by M6A-Seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Hongay, C.F.; Orr-Weaver, T.L. Drosophila inducer of meiosis 4 (IME4) is required for notch signaling during oogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 14855–14860. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Fu, Y.; He, C. Reversible RNA adenosine methylation in biological regulation. Trends Genet. 2013, 29, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.G.; et al. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.-Z.; MacQueen, A.; Zheng, G.; Duan, H.; Dore, L.C.; Lu, Z.; Liu, J.; Chen, K.; Jia, G.; Bergelson, J.; et al. Unique features of the M6A methylome in Arabidopsis thaliana. Nat. Commun. 2014. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.; Agarwala, S.D.; Mumbach, M.R.; Jovanovic, M.; Mertins, P.; Shishkin, A.; Tabach, Y.; Mikkelsen, T.S.; Satija, R.; Ruvkun, G.; et al. High-resolution mapping reveals a conserved, widespread, dynamic mRNA methylation program in yeast meiosis. Cell 2013, 155, 1409–1421. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fedorcsak, P.; Huang, C.M.; Li, C.J.; Vågbø, C.B.; Shi, Y.; Wang, W.L.; Song, S.H.; et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Li, H.; Bodi, Z.; Button, J.; Vespa, L.; Herzog, M.; Fray, R.G. MTA is an Arabidopsis messenger RNA adenosine methylase and interacts with a homolog of a sex-specific splicing factor. Plant Cell 2008, 20, 1278–1288. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.; Gruber, A.R.; Keller, W.; Zavolan, M. Genome-wide analysis of pre-mRNA 3' end processing reveals a decisive role of human cleavage factor I in the regulation of 3' UTR length. Cell Rep. 2012, 1, 753–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, T.; Wada, T.; Yamaguchi, Y.; Shimizu, A.; Handa, H. Knock-down of 25 kDa subunit of cleavage factor Im in hela cells alters alternative polyadenylation within 3'-UTRs. Nucleic Acids Res. 2006, 34, 6264–6271. [Google Scholar] [CrossRef] [PubMed]
- Masamha, C.P.; Xia, Z.; Yang, J.; Albrecht, T.R.; Li, M.; Shyu, A.B.; Li, W.; Wagner, E.J. CFIm25 links alternative polyadenylation to glioblastoma tumour suppression. Nature 2014, 510, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Jenal, M.; Elkon, R.; Loayza-Puch, F.; van Haaften, G.; Kühn, U.; Menzies, F.M.; Oude Vrielink, J.A.; Bos, A.J.; Drost, J.; Rooijers, K.; et al. The poly(A)-binding protein nuclear 1 suppresses alternative cleavage and polyadenylation sites. Cell 2012, 149, 538–553. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Biesinger, J.; Wan, J.; Weng, L.; Xing, Y.; Xie, X.; Shi, Y. Transcriptome-wide analyses of CSTF64-RNA interactions in global regulation of mRNA alternative polyadenylation. Proc. Natl. Acad. Sci. USA 2012, 109, 18773–18778. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Choi, E.A.; Weng, L.; Xie, X.; Wan, J.; Xing, Y.; Moresco, J.J.; Tu, P.G.; Yates, J.R.; Shi, Y. Overlapping and distinct functions of CSTF64 and CSTF64τ in mammalian mRNA 3' processing. RNA 2013, 19, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chakrabarti, M.; Hunt, A.G. CPSF30 at the Interface of Alternative Polyadenylation and Cellular Signaling in Plants. Biomolecules 2015, 5, 1151-1168. https://doi.org/10.3390/biom5021151
Chakrabarti M, Hunt AG. CPSF30 at the Interface of Alternative Polyadenylation and Cellular Signaling in Plants. Biomolecules. 2015; 5(2):1151-1168. https://doi.org/10.3390/biom5021151
Chicago/Turabian StyleChakrabarti, Manohar, and Arthur G. Hunt. 2015. "CPSF30 at the Interface of Alternative Polyadenylation and Cellular Signaling in Plants" Biomolecules 5, no. 2: 1151-1168. https://doi.org/10.3390/biom5021151
APA StyleChakrabarti, M., & Hunt, A. G. (2015). CPSF30 at the Interface of Alternative Polyadenylation and Cellular Signaling in Plants. Biomolecules, 5(2), 1151-1168. https://doi.org/10.3390/biom5021151