
RNase P-Mediated Sequence-Specific Cleavage of RNA by Engineered External Guide Sequences

Abstract
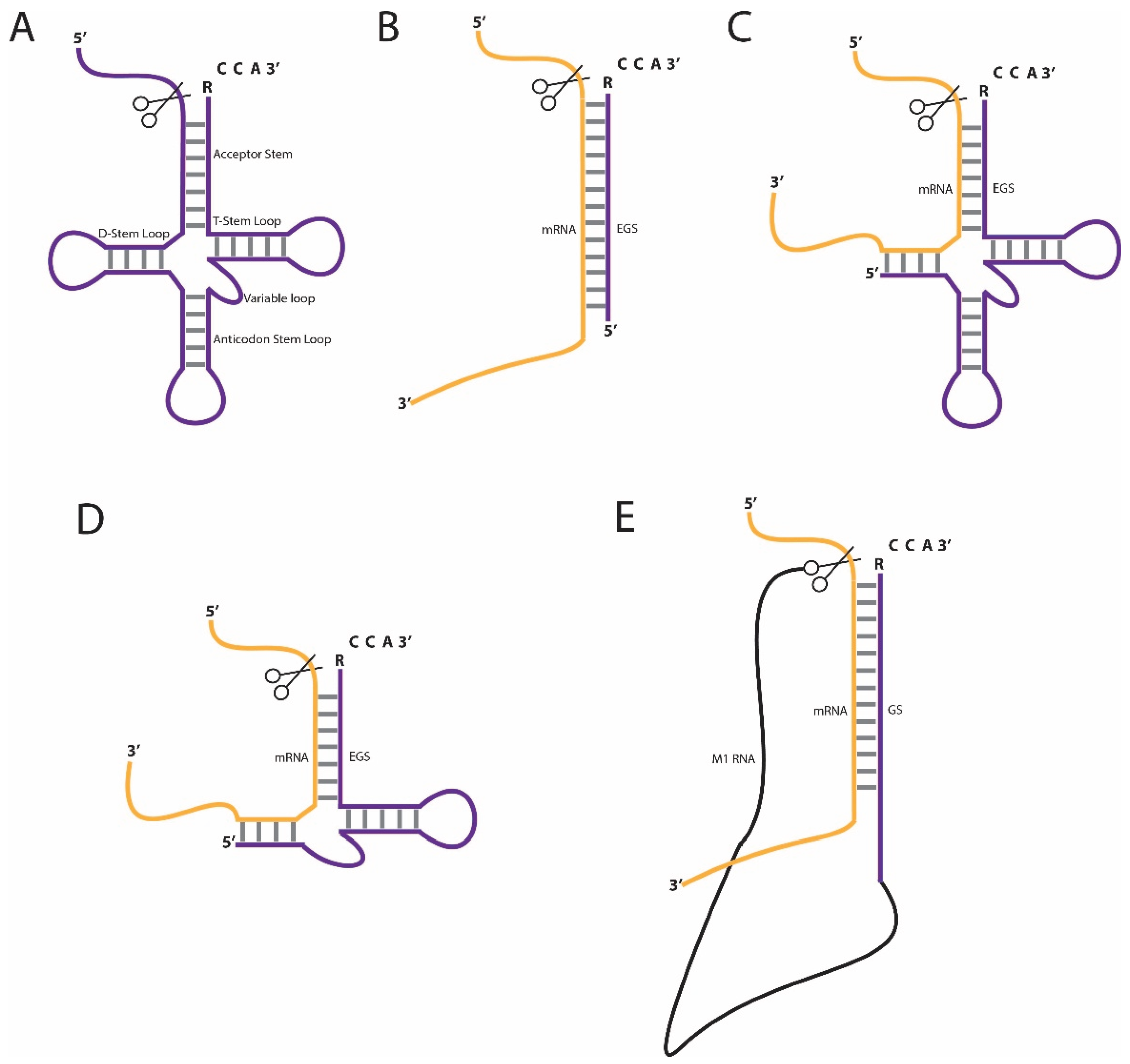
:1. Introduction

2. Approaches in RNase P-Mediated RNA Knockdown
3. Stabilization and Delivery of EGS/M1GS RNAs
3.1. Improved Nuclease Resistance
3.2. EGS and M1GS Delivery
4. Examples of RNase P-Based RNA Knockdown

{kind=link}
| Organism/Infectious Agent | Target Molecule | EGS/M1GS | Effect | System Tested | Reference |
|---|---|---|---|---|---|
| E. coli | bla, cat | EGS | Decreased viability | Cultured E. coli | [35] |
| CPP-PMO EGS | Decreased viability | Cultured E. coli | [28,30] | ||
| LNA/DNA EGS | Decreased viability | Cultured E. coli | [27] | ||
| gyrA, rnpA | EGS | Decreased viability | Cultured E. coli | [36] | |
| gyrA | CPP-PMO EGS | Decreased viability | Cultured E. coli and various other bacteria | [28,29,30] | |
| ftsZ | CPP-PMO EGS | Induced filimentation and ~10-fold decreased viability 1 | Cultured E. coli | [37] | |
| S. aureus | gyrA | CPP-PMO EGS | Accelerated epithelialization and wound closure and smaller eschar formation | Cultured S. aureus; infected murine cutaneous wound model | [38] |
| S. enterica | invB/invC | EGS | Reduced host-invasion | Salmonella invasion in Henle-407 cells | [39] |
| F. tularensis | mglB | EGS | Reduced mRNA levels | mglB expression in E. coli culture | [40] |
| Y. pestis | yscN, YscS | EGS | Reduced mRNA levels | yscN and yscS expression in E. coli culture | [41] |
| P. falciparum | PfGyrA | CPP-PMO EGS | 63%–75% growth inhibition measured in different strains | P. falciparum infected erythrocytes | [31] |
| Influenza virus | NP, PB2 | EGS | 70%–95% reduced expression of the viral M1 protein with the use of two EGSs simultaneously | Mouse C127 cells | [42] |
| Herpes simplex virus 1 | ICP4 | M1GS | 1000-fold reduced virus titer (36 h, MOI 2) | ψCRE cells (NIH 3T3 cells) | [43] |
| TK | Minimized EGS | 80% decrease in mRNA and 75% decrease in protein level | Human 143tk- cells | [44] | |
| Minimized EGS variant, improved by in vitro selection | 96% decrease in mRNA and 95% decrease in protein level | Human 143tk- cells | [45] | ||
| Human cytomegalo-virus | IE1, IE2 | WT M1GS | 500-fold reduced virus titer (5 days, MOI 1) | Human U373MG cells | [46] |
| M1GS—variant G224A, G225A | 3000-fold reduced virus titer (5 days, MOI 1) | Human U373MG cells | [47] | ||
| M1GS—variant U80C, C188U | 10,000-fold reduced virus titer (5 days, MOI 1 | Human U373MG cells | [48] | ||
| M1GS—variant A94G, G194C | 3500-fold reduced virus titer (5 days, MOI 1) | Human U373MG cells | [49] | ||
| AP/PR | Minimized EGS | 500-fold reduced virus titer (4 days, MOI 1) | Human foreskin fibroblasts | [50] | |
| Minimized EGS | 800-fold reduced virus titer (5 days ,MOI 2) | Human U373MG cells | [51] | ||
| Minimized EGS variant C321 | 7000-fold reduced virus titer (5 days, MOI 2) | Human U373MG cells | [52] | ||
| M1GS | 100-fold reduced virus titer (5 days, MOI 3) | Human U373MG cells | [53] | ||
| M1GS | 2000-fold reduced virus titer (5 days) | Human U373MG cells | [54] | ||
| M1GS—variant A81C, G194A | 50,000-fold reduced virus titer (5 days, MOI 1) | Human U251 cells | [55] | ||
| Murine cytomegalo-virus | AP/PR | M1GS; Salmonella SL101 | 2500-fold reduced virus titer (4 days, MOI 1) in macrophages; prolonged survival of mice | Mouse J774 macrophages; mice | [33] |
| Minimized EGS; Salmonella SL201 | 3000-fold reduced virus titer (4 days) in macrophages; prolonged survival of mice | Mouse J774 macrophages; mice | [34] | ||
| Hepatitis B virus | pgRNA, pre-S/L mRNA, S mRNA | Minimized EGS variant C418; Salmonella SL301 | 2000-fold reduced viral DNA level in HepG2.2.15 cells (3 days) and 200,000-fold reduced viral DNA level in mice (5 days) | HepG2.2.15 cells; mice | [56] |
| pgRNA, pre-S/L mRNA, S mRNA | Minimized EGS variant C386; Salmonella SL201 | 6000-fold reduced viral DNA level (4 days) | HepG2.2.15 cells | [57] | |
| Hepatitis C virus | HCV 5'-UTR | M1GS | >1000-fold reduced virus titer (1 day, MOI 1) | Human Huh7.5.1 cells | [58] |
| Human immuno-deficiency virus 1 | LTR and TAT | Minimized EGS | Reduced p24 protein levels | COS cells | [59] |
| TAT region | M1GS—variant G83U, G340A | 150-fold reduced virus titer (12 days) | Human H9 cells | [60] | |
| Humans | CCR5 | Minimized EGS | 50-fold reduced level of HIV p24 (12 days) | Human PM1 cells | [61] |
| BCR-ABL | M1GS | 96% and 97% cell death in respectively, p190 and p210 dependent cells | Ba/F3 cells expressing the p190 and p210 oncogenes | [62] |
4.1. Downregulation of Bacterial RNAs and Infections
4.1.1. Drug Resistance Genes
4.1.2. Essential Genes
4.1.3. Virulence Genes
4.2. Downregulation of Malaria Transcripts and Infections
4.3. Downregulation of Viral RNAs and Infections
4.3.1. Cytomegalovirus
4.3.2. Hepatitis B Virus
4.3.3. Hepatitis C Virus
4.4. Downregulation of Human Genes to Target HIV1
4.5. Downregulation of the BCR-ABL Oncogenic Transcript
4.6. Prophylactic vs. Therapeutic Treatment with EGS/M1GS
5. EGS/M1GS Approaches vs. RNA Interference
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Robertson, H.D.; Altman, S.; Smith, J.D. Purification and properties of a specific Escheria coli ribonuclease which cleaves a tyrosine transfer riboncleic acid precursor. J. Biol. Chem. 1972, 274, 5243–5251. [Google Scholar]
- Hernandez-Cid, A.; Aguirre-Sampieri, S.; Diaz-Vilchis, A.; Torres-Larios, A. Ribonucleases P/MRP and the expanding ribonucleoprotein world. Int. Union Biochem. Mol. Biol. Life 2012, 64, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Guerrier-Takada, C.; Gardiner, K.; Marsh, T.; Pace, N.; Altman, S. The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. Cell 1983, 35, 849–857. [Google Scholar] [CrossRef]
- Pannucci, J.A.; Haas, E.S.; Hall, T.A.; Harris, J.K.; Brown, J.W. RNase P RNAs from some Archaea are catalytically active. Proc. Natl. Acad. Sci. USA 1999, 96, 7803–7808. [Google Scholar] [CrossRef] [PubMed]
- Kikovska, E.; Svärd, S.G.; Kirsebom, L.A. Eukaryotic RNase P RNA mediates cleavage in the absence of protein. Proc. Natl. Acad. Sci. USA 2007, 104, 2062–2067. [Google Scholar] [CrossRef] [PubMed]
- Holzmann, J.; Frank, P.; Löffler, E.; Bennett, K.L.; Gerner, C.; Rossmanith, W. RNase P without RNA: Identification and functional reconstitution of the human mitochondrial tRNA processing enzyme. Cell 2008, 135, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Gobert, A.; Gutmann, B.; Taschner, A.; Gössringer, M.; Holzmann, J.; Hartmann, R.K.; Rossmanith, W.; Giegé, P. A single Arabidopsis organellar protein has RNase P activity. Nat. Struct. Mol. Biol. 2010, 17, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Gutmann, B.; Gobert, A.; Giegé, P. PRORP proteins support RNase P activity in both organelles and the nucleus in Arabidopsis. Genes Dev. 2012, 26, 1022–1027. [Google Scholar] [CrossRef] [PubMed]
- Taschner, A.; Weber, C.; Buzet, A.; Hartmann, R.K.; Hartig, A.; Rossmanith, W. Nuclear RNase P of Trypanosoma brucei: A single protein in place of the multicomponent RNA-protein complex. Cell Rep. 2012, 2, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Sugita, C.; Komura, Y.; Tanaka, K.; Kometani, K.; Satoh, H.; Sugita, M. Molecular characterization of three PRORP proteins in the moss Physcomitrella patens: nuclear PRORP protein is not essential for moss viability. PLoS ONE 2014, 9, e108962. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.; Hartig, A.; Hartmann, R.K.; Rossmanith, W. Playing RNase P evolution: Swapping the RNA catalyst for a protein reveals functional uniformity of highly divergent enzyme forms. PLoS Genet. 2014, 10, e1004506. [Google Scholar] [CrossRef] [PubMed]
- McClain, W.H.; Guerrier-Takada, C.; Altman, S. Model substrates for an RNA enzyme. Science 1987, 238, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Forster, A.C.; Altman, S. External Guide Sequences for an RNA molecule. Science 1990, 249, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Guerrier-Takada, C.; Altman, S. Targeted cleavage of mRNA in vitro by RNase P from Escherichia coli. Proc. Natl. Acad. Sci. USA 1992, 89, 3185–3189. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Hwang, E.S.; Altman, S. Targeted cleavage of mRNA by human RNase P. Proc. Natl. Acad. Sci. USA 1992, 89, 8006–8010. [Google Scholar] [CrossRef] [PubMed]
- Raj, M.L.; Pulukkunat, D.K.; Reckard, J.F.; Thomas, G.; Gopalan, V. Cleavage of bipartite substrates by rice and maize ribonuclease P. Application to degradation of target mRNAs in plants. Plant Physiol. 2001, 125, 1187–1190. [Google Scholar] [CrossRef] [PubMed]
- Rangarajan, S.; Raj, M.L.S.; Hernandez, J.M.; Grotewold, E.; Gopalan, V. RNase P as a tool for disruption of gene expression in maize cells. Biochem. J. 2004, 380, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Guerrier-Takada, C.; Li, Y.; Altman, S. Artificial regulation of gene expression in Escherichia coli by RNase P. Proc. Natl. Acad. Sci. USA 1995, 92, 11115–11119. [Google Scholar] [CrossRef] [PubMed]
- Wegscheid, B.; Hartmann, R.K. The precursor tRNA 3'-CCA interaction with Escherichia coli RNase P RNA is essential for catalysis by RNase P in vivo. RNA 2006, 12, 2135–2148. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Altman, S. Selection of guide sequences that direct efficient cleavage of mRNA by human ribonuclease P. Science 1994, 263, 1269–1273. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Altman, S. Substrate recognition by human RNase P: Identification of small, model substrates for the enzyme. EMBO J. 1995, 14, 159–168. [Google Scholar] [PubMed]
- Ma, M.; Benimetskaya, L.; Lebedeva, I.; Dignam, J.; Takle, G.; Stein, C.A. Intracellular mRNA cleavage induced through activation of RNase P by nuclease-resistant external guide sequences. Nat. Biotechnol. 2000, 18, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Altman, S. Inhibition of viral gene expression by the catalytic RNA subunit of RNase P from Escherichia coli. Genes Dev. 1995, 9, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Altman, S. Cleavage by RNase P of gene N mRNA reduces bacteriophage λ burst size. Nucleic Acids Res. 1996, 24, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.Y.; Jacob-samuel, B.U.I.; Dignam, J.C.; Pace, U.; Goldberg, A.R.; George, S.T. Nuclease-resistanct external guide sequence-induced cleavage of target RNA by human ribonuclease P. Antisense Nucleic Acid Drug Dev. 1998, 8, 415–426. [Google Scholar] [PubMed]
- Hudziak, R.M.; Barofsky, E.; Barofsky, D.F.; Weller, D.L.; Huang, S.B.; Weller, D.D. Resistance of morpholino phosphodiamidate oligomers to enzymatic degradation. Antisense Nucleic Acid Drug Dev. 1996, 6, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Soler Bistué, A.J.C.; Martín, F.A.; Vozza, N.; Ha, H.; Joaquín, J.C.; Zorreguieta, A.; Tolmasky, M.E. Inhibition of AAC(6')-Ib-mediated amikacin resistance by nuclease-resistant external guide sequences in bacteria. Proc. Natl. Acad. Sci. USA 2009, 106, 13230–13235. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Ko, J.; Xiao, G.; Wesolowski, D.; Shan, G.; Geller, B.; Izadjoo, M.; Altman, S. Inactivation of expression of several genes in a variety of bacterial species by EGS technology. Proc. Natl. Acad. Sci. USA 2009, 106, 8163–8168. [Google Scholar] [CrossRef] [PubMed]
- Wesolowski, D.; Alonso, D.; Altman, S. Combined effect of a peptide-morpholino oligonucleotide conjugate and a cell-penetrating peptide as an antibiotic. Proc. Natl. Acad. Sci. USA 2013, 110, 8686–8689. [Google Scholar] [CrossRef] [PubMed]
- Wesolowski, D.; Tae, H.S.; Gandotra, N.; Llopis, P.; Shen, N.; Altman, S. Basic peptide-morpholino oligomer conjugate that is very effective in killing bacteria by gene-specific and nonspecific modes. Proc. Natl. Acad. Sci. USA 2011, 108, 16582–16587. [Google Scholar] [CrossRef] [PubMed]
- Augagneur, Y.; Wesolowski, D.; Tae, H.S.; Altman, S.; Ben Mamoun, C. Gene selective mRNA cleavage inhibits the development of Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2012, 109, 6235–6240. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Li, H.; Vu, G.P.; Gong, H.; Umamoto, S.; Zhou, T.; Lu, S.; Liu, F. Salmonella-mediated delivery of RNase P-based ribozymes for inhibition of viral gene expression and replication in human cells. Proc. Natl. Acad. Sci. USA 2010, 107, 7269–7274. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Gong, H.; Li, H.; Vu, G.P.; Lu, S.; Liu, F. Oral delivery of RNase P ribozymes by Salmonella inhibits viral infection in mice. Proc. Natl. Acad. Sci. USA 2011, 108, 3222–3227. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Gong, H.; Chen, Y.C.; Vu, G.P.; Trang, P.; Zhang, C.Y.; Lu, S.; Liu, F. Effective inhibition of cytomegalovirus infection by external guide sequences in mice. Proc. Natl. Acad. Sci. USA 2012, 109, 13070–13075. [Google Scholar] [CrossRef] [PubMed]
- Guerrier-Takada, C.; Salavati, R.; Altman, S. Phenotypic conversion of drug-resistant bacteria to drug sensitivity. Proc. Natl. Acad. Sci. USA 1997, 94, 8468–8472. [Google Scholar] [CrossRef] [PubMed]
- McKinney, J.; Guerrier-Takada, C.; Wesolowski, D.; Altman, S. Inhibition of Escherichia coli viability by external guide sequences complementary to two essential genes. Proc. Natl. Acad. Sci. USA 2001, 98, 6605–6610. [Google Scholar] [CrossRef] [PubMed]
- Sala, C.D.; Soler-Bistué, A.J.C.; Korprapun, L.; Zorreguieta, A.; Tolmasky, M.E. Inhibition of cell division induced by external guide sequences (EGS Technology) targeting ftsZ. PLoS ONE 2012, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, A.J.; Wesolowski, D.; Gandotra, N.; Stojadinovic, A.; Izadjoo, M.; Altman, S.; Kyriakides, T.R. A peptide-morpholino oligomer conjugate targeting Staphylococcus aureus gyrA mRNA improves healing in an infected mouse cutaneous wound model. Int. J. Pharm. 2013, 453, 651–655. [Google Scholar] [CrossRef] [PubMed]
- McKinney, J.S.; Zhang, H.; Kubori, T.; Galán, J.E.; Altman, S. Disruption of type III secretion in Salmonella enterica serovar Typhimurium by external guide sequences. Nucleic Acids Res. 2004, 32, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Lundblad, E.W.; Izadjoo, M.; Altman, S. Inhibition of expression in Escherichia coli of a virulence regulator MglB of Francisella tularensis using external guide sequence technology. PLoS ONE 2008, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.; Izadjoo, M.; Altman, S. Inhibition of expression of virulence genes of Yersinia pestis in Escherichia coli by external guide sequences and RNase P. RNA 2008, 14, 1656–1662. [Google Scholar] [CrossRef] [PubMed]
- Plehn-Dujowich, D.; Altman, S. Effective inhibition of influenza virus production in cultured cells by external guide sequences and ribonuclease P. Proc. Natl. Acad. Sci. USA 1998, 95, 7327–7332. [Google Scholar] [CrossRef] [PubMed]
- Trang, P.; Kilani, A.; Kim, J.; Liu, F. A ribozyme derived from the catalytic subunit of RNase P from Escherichia coli is highly effective in inhibiting replication of herpes simplex virus 1. J. Mol. Biol. 2000, 301, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Kawa, D.; Wang, J.; Yuan, Y.; Liu, F. Inhibition of viral gene expression by human ribonuclease P. RNA 1998, 4, 1397–1406. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Kim, J.; Kilani, A.F.; Kim, K.; Dunn, W.; Jo, S.; Nepomuceno, E.; Liu, F. In vitro selection of external guide sequences for directing RNase P-mediated inhibition of viral gene expression. J. Biol. Chem. 2002, 277, 30112–30120. [Google Scholar] [CrossRef] [PubMed]
- Trang, P.; Kilani, A.; Lee, J.; Hsu, A.; Liou, K.; Kim, J.; Nassi, A.; Kim, K.; Liu, F. RNase P ribozymes for the studies and treatment of human cytomegalo virus infections. J. Clin. Virol. 2002, 25, S63–S74. [Google Scholar] [CrossRef]
- Trang, P.; Hsu, A.; Zhou, T.; Lee, J.; Kilani, A.F.; Nepomuceno, E.; Liu, F. Engineered RNase P ribozymes inhibit gene expression and growth of cytomegalovirus by increasing rate of cleavage and substrate binding. J. Mol. Biol. 2002, 315, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Lee, J.; Umamoto, S.; Kilani, A.F.; Kim, J.; Trang, P.; Zhou, T.; Liu, F. Engineered RNase P ribozymes are efficient in cleaving a human cytomegalovirus mRNA in vitro and are effective in inhibiting viral gene expression and growth in human cells. J. Biol. Chem. 2003, 278, 37265–37274. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Vu, G.P.; Qian, H.; Chen, Y.C.; Wang, Y.; Reeves, M.; Zen, K.; Liu, F. Engineered RNase P ribozymes effectively inhibit human cytomegalovirus gene expression and replication. Viruses 2014, 6, 2376–2391. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.; Trang, P.; Khan, U.; Zhu, J.; Liu, F. RNase P-mediated inhibition of cytomegalovirus protease expression and viral DNA encapsidation by oligonucleotide external guide sequences. Proc. Natl. Acad. Sci. USA 2001, 98, 14831–14836. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Trang, P.; Kim, K.; Zhou, T.; Umamoto, S.; Liu, F. Effective inhibition of human cytomegalovirus gene expression and growth by intracellular expression of external guide sequence RNA. RNA 2006, 12, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Chen, Y.C.; Gong, H.; Trang, P.; Lu, S.; Liu, F. Ribonuclease P-mediated inhibition of human cytomegalovirus gene expression and replication induced by engineered external guide sequences. RNA Biol. 2012, 9, 1186–1195. [Google Scholar] [CrossRef] [PubMed]
- Trang, P.; Kim, K.; Zhu, J.; Liu, F. Expression of an RNase P ribozyme against the mRNA encoding human cytomegalovirus protease inhibits viral capsid protein processing and growth. J. Mol. Biol. 2003, 328, 1123–1135. [Google Scholar] [CrossRef]
- Kim, K.; Umamoto, S.; Trang, P.; Hai, R.; Liu, F.Y. Intracellular expression of engineered RNase P ribozymes effectively blocks gene expression and replication of human cytomegalovirus. RNA 2004, 10, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Reeves, M.; Ye, J.; Trang, P.; Zhu, L.; Sheng, J.; Wang, Y.; Zen, K.; Wu, J.; Liu, F. RNase P ribozymes inhibit the replication of human cytomegalovirus by targeting essential viral capsid proteins. Viruses 2015, 7, 3345–3360. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Chen, Y.C.; Gong, H.; Zeng, W.; Vu, G.P.; Trang, P.; Lu, S.; Wu, J.; Liu, F. Inhibition of hepatitis B virus gene expression and replication by ribonuclease P. Mol. Ther. 2013, 21, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Vu, G.P.; Gong, H.; Xia, C.; Chen, Y.C.; Liu, F.; Wu, J.; Lu, S. Engineered external guide sequences are highly effective in inhibiting gene expression and replication of hepatitis B virus in cultured cells. PLoS ONE 2013, 8, e65268. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Li, X.; Mao, X.; Huang, Z.; Zhang, C.; Zhang, W.; Wu, J.; Li, G. Inhibition of hepatitis C virus by an M1GS ribozyme derived from the catalytic RNA subunit of Escherichia coli RNase P. Virol. J. 2014. [Google Scholar] [CrossRef] [PubMed]
- Barnor, J.S.; Endo, Y.; Habu, Y.; Miyano-Kurosaki, N.; Kitano, M.; Yamamoto, H.; Takaku, H. Effective inhibition of HIV-1 replication in cultured cells by external guide sequences and ribonuclease P. Bioorg. Med. Chem. Lett. 2004, 14, 4941–4944. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Chen, Y.C.; Bai, Y.; Trang, P.; Vu, G.P.; Lu, S.; Wu, J.; Liu, F. Effective inhibition of human immunodeficiency virus 1 replication by engineered RNase P ribozyme. PLoS ONE 2012, 7, e51855. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Vu, G.P.; Bai, Y.; Chen, Y.C.; Trang, P.; Lu, S.; Xiao, G.; Liu, F. RNase P-associated external guide sequence effectively reduces the expression of human CC-chemokine receptor 5 and inhibits the infection of human immunodeficiency virus 1. BioMed Res. Int. 2013. [Google Scholar] [CrossRef] [PubMed]
- Cobaleda, C.; Sánchez-García, I. In vivo inhibition by a site-specific catalytic RNA subunit of RNase P designed against the BCR-ABL oncogenic products: A novel approach for cancer treatment. Blood 2000, 95, 731–737. [Google Scholar] [PubMed]
- Tolmasky, M.E. Aminoglycoside-modifying enzymes: Characteristics, localization, and dissemination. In Enzyme-Mediated Resistance to Antibiotics; Bonomo, R., Tolmask, A., Eds.; ASM Press: Washington, DC, USA, 2007; pp. 35–52. [Google Scholar]
- Soler Bistué, A.J.C.; Ha, H.; Sarno, R.; Don, M.; Zorreguieta, A.; Tolmasky, M.E. External guide sequences targeting the AAC(6ʹ)-Ib mRNA induce inhibition of amikacin resistance. Antimicrob. Agents Chemother. 2007, 51, 1918–1925. [Google Scholar] [CrossRef] [PubMed]
- Eichelberg, K.; Ginocchio, C.C.; Galan, J.E. Molecular and functional characterization of the Salmonella typhimurium invasion genes invB and invC: Homology of invC to the F0F1 ATPase family of proteins. J. Bacteriol. 1994, 176, 4501–4510. [Google Scholar] [PubMed]
- Bronstein, P.A.; Miao, E.A. InvB is a type III secretion chaperone specific for SspA. J. Bacteriol. 2000, 182, 6638–6644. [Google Scholar] [CrossRef] [PubMed]
- De Luca, N.A.; McCarthy, A.M.; Schaffer, P.A. Isolation and characterization of deletion mutants of herpes simplex virus type 1 in the gene encoding immediate-early regulatory protein ICP4. J. Virol. 1985, 56, 558–570. [Google Scholar]
- Mocarski, E.S.; Shenk, T.; Pass, R. Cytomegalovirus. In Fields Virology; Knipe, D., Howley, P., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007; pp. 2701–2772. [Google Scholar]
- Trang, P.; Lee, M.; Nepomuceno, E.; Kim, J.; Zhu, H.; Liu, F. Effective inhibition of human cytomegalovirus gene expression and replication by a ribozyme derived from the catalytic RNA subunit of RNase P from Escherichia coli. Proc. Natl. Acad. Sci. USA 2000, 97, 5812–5817. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Li, H.; Li, Y.; Cui, Y.; Zhou, Q.; Yang, G.; Zhou, T. Effective inhibition of human cytomegalovirus gene expression by DNA-based external guide sequences. Acta Biochim. Biophys. Sin. 2009, 41, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, H.; Li, Y.; Zeng, Z.; Li, S.; Zhang, X.; Zou, Y.; Zhou, T. Effective inhibition of HCMV UL49 gene expression and viral replication by oligonucleotide external guide sequences and RNase P. Virol. J. 2010. [Google Scholar] [CrossRef] [PubMed]
- Seegar, C.; Zoulim, F.; Mason, W. Hepadnaviruses. In Fields Virology; Knipe, D., Howley, P., Eds.; Lippincott Willam and Wilkins: Philadelphia, PA, USA, 2007; pp. 2978–3029. [Google Scholar]
- Guidotti, L.G.; Chisari, F.V. Immunobiology and pathogeneis of viral hepatitis. Annu. Rev. Pathol. 2006, 1, 23–61. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Grisé, H. Cellular and molecular biology of HCV infection and hepatitis. Clin. Sci. 2006, 117, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Freed, E.; Martin, M. HIVs and their replication. In Fields Virology; Knipe, D., Howley, P., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007; pp. 2108–2185. [Google Scholar]
- Kurtizkes, D.; Walker, B. HIV-1: Pathogenesis, clinical manifestations, and treatment. In Fields Virology; Knipe, D., Howley, P., Eds.; Lippincott Willam and Wilkins: Philadelphia, PA, USA, 2007; pp. 2188–2214. [Google Scholar]
- Hermans, A.; Heisterkamp, N.; von Linden, M.; van Baal, S.; Meijer, D.; van der Plas, D.; Wiedemann, L.M.; Groffen, J.; Bootsma, D.; Grosveld, G. Unique fusion of BCR and C-ABL genes in Philadelphia chromosome positive acute lymphoblastic leukemia. Cell 1987, 51, 33–40. [Google Scholar] [CrossRef]
- Ben-Neriah, Y.; Daley, G.Q.; Mes-Masson, A.M.; Witte, O.N.; Baltimore, D. The chronic myelogenous leukemia-specific P210 protein is the product of the bcr/abl hyrbrid gene. Science 1986, 233, 212–214. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-García, I.; Grütz, G. Tumorigenic activity of the BCR-ABL oncogenes is mediated by BCL2. Proc. Natl. Acad. Sci. USA 1995, 92, 5287–5291. [Google Scholar] [CrossRef] [PubMed]
- Wittrup, A.; Lieberman, J. Knocking down disease: A progress report on siRNA therapeutics. Nat. Rev. Genet. 2015, 16, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Altman, S. Inhibition of the expression of the human RNase P protein subunits Rpp21, Rpp25, Rpp29 by external guide sequences (EGSs) and siRNA. J. Mol. Biol. 2004, 342, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, D.L.; Shashikant, C.; Hayday, A.C. A comparative analysis of RNA targeting strategies in the thymosin beta 4 gene. J. Mol. Biol. 2004, 342, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Derksen, M.; Mertens, V.; Pruijn, G.J.M. RNase P-Mediated Sequence-Specific Cleavage of RNA by Engineered External Guide Sequences. Biomolecules 2015, 5, 3029-3050. https://doi.org/10.3390/biom5043029
Derksen M, Mertens V, Pruijn GJM. RNase P-Mediated Sequence-Specific Cleavage of RNA by Engineered External Guide Sequences. Biomolecules. 2015; 5(4):3029-3050. https://doi.org/10.3390/biom5043029
Chicago/Turabian StyleDerksen, Merel, Vicky Mertens, and Ger J.M. Pruijn. 2015. "RNase P-Mediated Sequence-Specific Cleavage of RNA by Engineered External Guide Sequences" Biomolecules 5, no. 4: 3029-3050. https://doi.org/10.3390/biom5043029
APA StyleDerksen, M., Mertens, V., & Pruijn, G. J. M. (2015). RNase P-Mediated Sequence-Specific Cleavage of RNA by Engineered External Guide Sequences. Biomolecules, 5(4), 3029-3050. https://doi.org/10.3390/biom5043029