
Guardian of Genetic Messenger-RNA-Binding Proteins


{kind=link}
{kind=link}
Abstract
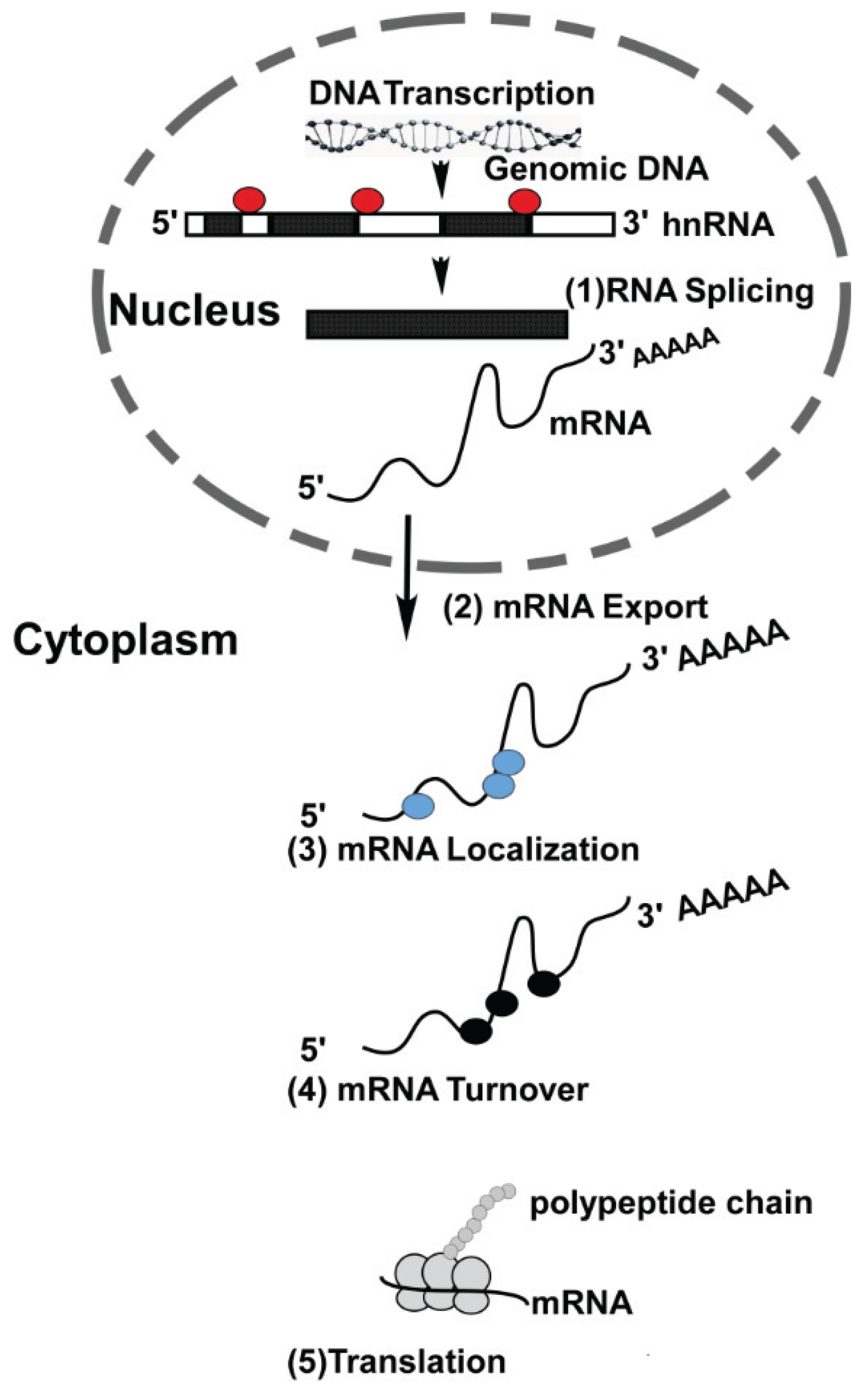
:1. Introduction

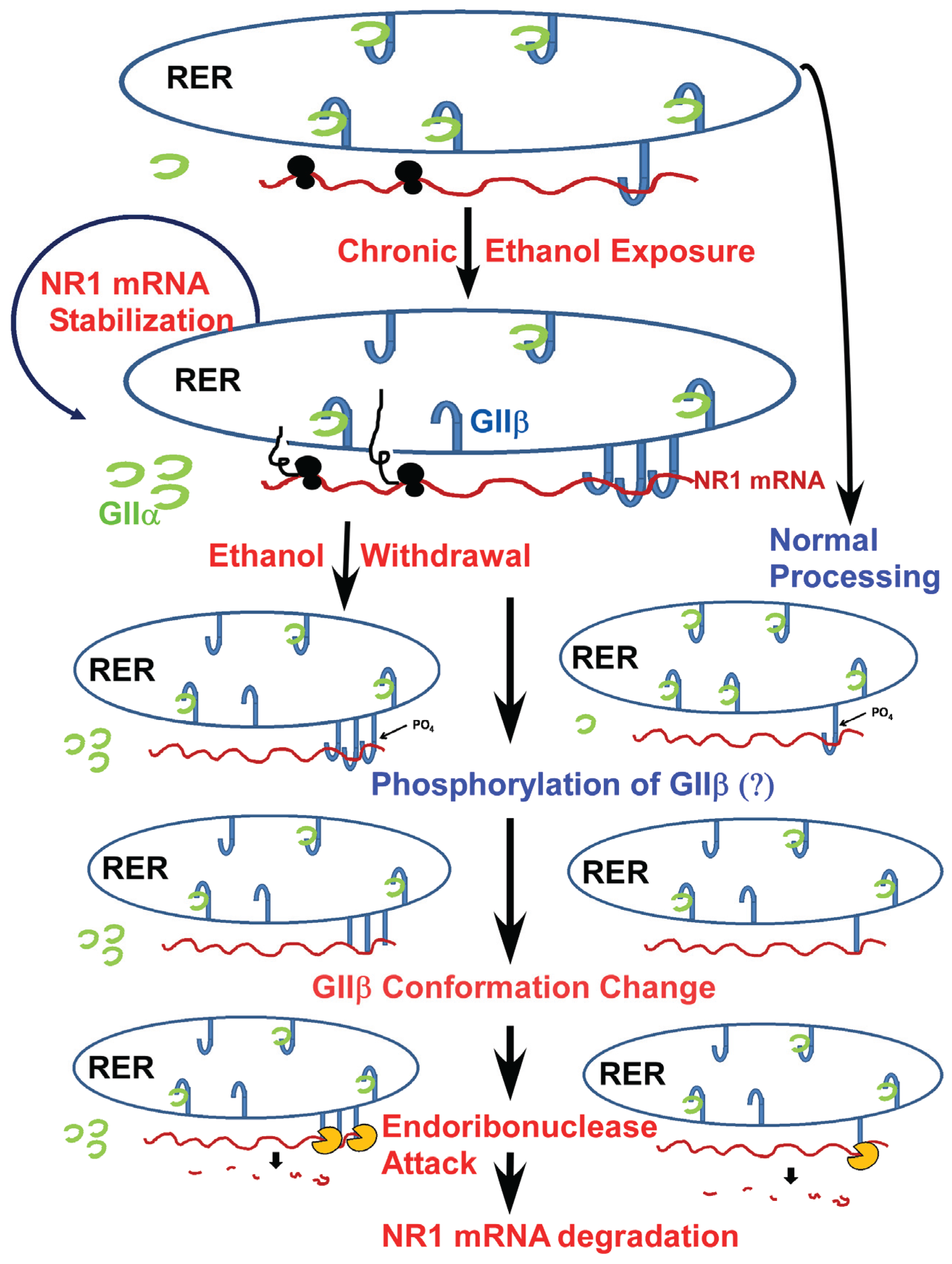
2. RNA-Binding Proteins and Alcohol

3. RNA-Binding Proteins in Neurological Diseases
4. RNA-Binding Proteins and Cancer
5. Approaches to Detection of RBPs
6. Computational Methods
7. Conclusions and Future Directions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Conne, B.; Stutz, A.; Vassalli, J.D. The 3' untranslated region of messenger RNA: A molecular “hotspot” for pathology? Nat. Med. 2000, 6, 637–641. [Google Scholar] [CrossRef] [PubMed]
- Baltz, A.G.; Munschauer, M.; Schwanhäusser, B.; Vasile, A.; Murakawa, Y.; Schueler, M.; Youngs, N.; Penfold-Brown, D.; Drew, K.; Milek, M.; et al. The mRNA-bound proteome and its global occupancy profile on protein-coding transcripts. Mol. Cell 2012, 46, 674–690. [Google Scholar] [CrossRef] [PubMed]
- Castello, A.; Fischer, B.; Eichelbaum, K.; Horos, R.; Beckmann, B.M.; Strein, C.; Davey, N.E.; Humphreys, D.T.; Preiss, T.; Steinmetz, L.M.; et al. Insights into RNA biology from an atlas of mammalian mRNA-binding proteins. Cell 2012, 149, 1393–1406. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, S.; Okano, H. Expression of neural RNA-binding proteins in the postnatal CNS: Implications of their roles in neuronal and glial cell development. J. Neurosci. 1997, 17, 8300–8312. [Google Scholar] [PubMed]
- Jiang, H.; Xu, L.; Wang, Z.; Keene, J.; Gu, Z. Coordinating expression of RNA binding proteins with their mRNA targets. Sci. Rep. 2014. [Google Scholar] [CrossRef] [PubMed]
- Kishore, S.; Luber, S.; Zavolan, M. Deciphering the role of RNA-binding proteins in the post-transcriptional control of gene expression. Brief. Funct. Genomics 2010, 9, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Lunde, B.M.; Moore, C.; Varani, G. RNA-binding proteins: Modular design for efficient function. Nat. Rev. Mol. Cell Biol. 2007, 8, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Ban, T.; Zhu, J.K.; Melcher, K.; Xu, H.E. Structural mechanisms of RNA recognition: Sequence-specific and non-specific RNA-binding proteins and the CAS9-RNA-DNA complex. Cell. Mol. Life Sci. 2015, 72, 1045–1058. [Google Scholar] [CrossRef] [PubMed]
- Gerstberger, S.; Hafner, M.; Tuschl, T. A census of human RNA-binding proteins. Nat. Rev. Genet. 2014, 15, 829–845. [Google Scholar] [CrossRef] [PubMed]
- Neelamraju, Y.; Hashemikhabir, S.; Janga, S.C. The human RBPome: From genes and proteins to human disease. J. Proteom. 2015, 127, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, D.; Kazan, H.; Cook, K.B.; Weirauch, M.T.; Najafabadi, H.S.; Li, X.; Gueroussov, S.; Albu, M.; Zheng, H.; Yang, A.; et al. A compendium of RNA-binding motifs for decoding gene regulation. Nature 2013, 499, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Ciafre, S.A.; Galardi, S. MicroRNAs and RNA-binding proteins: A complex network of interactions and reciprocal regulations in cancer. RNA Biol. 2013, 10, 935–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohr, A.M.; Mott, J.L. Overview of microRNA biology. Semin. Liver Dis. 2015, 35, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Preusse, M.; Marr, C.; Saunders, S.; Maticzka, D.; Lickert, H.; Backofen, R.; Theis, F. SimiRa: A tool to identify coregulation between microRNAs and RNA-binding proteins. RNA Biol. 2015, 12, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Coller, H. Functional interactions between microRNAs and RNA binding proteins. MicroRNA 2012, 1, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence micrornas using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [PubMed]
- Bahrami-Samani, E.; Vo, D.T.; de Araujo, P.R.; Vogel, C.; Smith, A.D.; Penalva, L.O.; Uren, P.J. Computational challenges, tools, and resources for analyzing co- and post-transcriptional events in high throughput. Wiley Interdiscip. Rev. RNA 2015, 6, 291–310. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, N.; Corcoran, D.L.; Nusbaum, J.D.; Reid, D.W.; Georgiev, S.; Hafner, M.; Ascano, M., Jr.; Tuschl, T.; Ohler, U.; Keene, J.D. Integrative regulatory mapping indicates that the RNA-binding protein HuR couples pre-mRNA processing and mRNA stability. Mol. Cell 2011, 43, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Liu, S.; Zhou, H.; Qu, L.H.; Yang, J.H. Starbase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale clip-seq data. Nucleic Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef] [PubMed]
- NIH-NIAAA. Alcohol facts and statistics. Available online: http://pubs.niaaa.nih.gov/publications/AlcoholFacts&Stats/AlcoholFacts&Stats.htm (accessed on 10 December 2015).
- Gukovskaya, A.S.; Mouria, M.; Gukovsky, I.; Reyes, C.N.; Kasho, V.N.; Faller, L.D.; Pandol, S.J. Ethanol metabolism and transcription factor activation in pancreatic acinar cells in rats. Gastroenterology 2002, 122, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Ponomarev, I. Epigenetic control of gene expression in the alcoholic brain. Alcohol Res. Curr. Rev. 2013, 35, 69–76. [Google Scholar]
- Pandey, S.C. The gene transcription factor cyclic AMP-responsive element binding protein: Role in positive and negative affective states of alcohol addiction. Pharmacol. Ther. 2004, 104, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Lonze, B.E.; Ginty, D.D. Function and regulation of CREB family transcription factors in the nervous system. Neuron 2002, 35, 605–623. [Google Scholar] [CrossRef]
- Moonat, S.; Starkman, B.G.; Sakharkar, A.; Pandey, S.C. Neuroscience of alcoholism: Molecular and cellular mechanisms. Cell. Mol. Life Sci. 2010, 67, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Vengeliene, V.; Bilbao, A.; Molander, A.; Spanagel, R. Neuropharmacology of alcohol addiction. Br. J. Pharmacol. 2008, 154, 299–315. [Google Scholar] [CrossRef] [PubMed]
- Ron, D.; Wang, J. The NMDA Receptor and Alcohol Addiction. In Biology of the NMDA Receptor; Taylor & Francis Group, LLC: Boca Raton, FL, USA, 2009. [Google Scholar]
- Traynelis, S.F.; Wollmuth, L.P.; McBain, C.J.; Menniti, F.S.; Vance, K.M.; Ogden, K.K.; Hansen, K.B.; Yuan, H.; Myers, S.J.; Dingledine, R. Glutamate receptor ion channels: Structure, regulation, and function. Pharmacol. Rev. 2010, 62, 405–496. [Google Scholar] [CrossRef] [PubMed]
- Forrest, D.; Yuzaki, M.; Soares, H.D.; Ng, L.; Luk, D.C.; Sheng, M.; Stewart, C.L.; Morgan, J.I.; Connor, J.A.; Curran, T. Targeted disruption of NMDA receptor 1 gene abolishes NMDA response and results in neonatal death. Neuron 1994, 13, 325–338. [Google Scholar] [CrossRef]
- Hollmann, M.; Boulter, J.; Maron, C.; Beasley, L.; Sullivan, J.; Pecht, G.; Heinemann, S. Zinc potentiates agonist-induced currents at certain splice variants of the NMDA receptor. Neuron 1993, 10, 943–954. [Google Scholar] [CrossRef]
- Sanna, E.; Serra, M.; Cossu, A.; Colombo, G.; Follesa, P.; Cuccheddu, T.; Concas, A.; Biggio, G. Chronic ethanol intoxication induces differential effects on GABAA and NMDA receptor function in the rat brain. Alcohol. Clin. Exp. Res. 1993, 17, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.J.; Ticku, M.K. Chronic ethanol treatment upregulates the NMDA receptor function and binding in mammalian cortical neurons. Brain Res. Mol. Brain Res. 1995, 30, 347–356. [Google Scholar] [CrossRef]
- Kumari, M.; Ticku, M.K. Regulation of NMDA receptors by ethanol. In Progress in Drug Research; Birkhäuser: Basel, Switzerland, 2000; Volume 54, pp. 151–189. [Google Scholar]
- Trevisan, L.; Fitzgerald, L.W.; Brose, N.; Gasic, G.P.; Heinemann, S.F.; Duman, R.S.; Nestler, E.J. Chronic ingestion of ethanol up-regulates NMDAR1 receptor subunit immunoreactivity in rat hippocampus. J. Neurochem. 1994, 62, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Ticku, M.K. Ethanol and regulation of the NMDA receptor subunits in fetal cortical neurons. J. Neurochem. 1998, 70, 1467–1473. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Anji, A.; Woods, H., Jr.; Ticku, M.K. The molecular effects of alcohol: Clues to the enigmatic action of alcohol. Ann. N. Y. Acad. Sci. 2003, 993, 82–94, discussion 123–124. [Google Scholar] [CrossRef] [PubMed]
- Hollas, H.; Aukrust, I.; Grimmer, S.; Strand, E.; Flatmark, T.; Vedeler, A. Annexin A2 recognises a specific region in the 3'-UTR of its cognate messenger RNA. Biochim. Biophys. Acta 2006, 1763, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Anji, A.; Kumari, M. A novel RNA binding protein that interacts with NMDA R1 mRNA: Regulation by ethanol. Eur. J. Neurosci. 2006, 23, 2339–2350. [Google Scholar] [CrossRef] [PubMed]
- Anji, A.; Kumari, M. A cis-acting region in the N-methyl-d-aspartate R1 3'-untranslated region interacts with the novel RNA-binding proteins beta subunit of alpha glucosidase II and annexin A2—Effect of chronic ethanol exposure in vivo. Eur. J. Neurosci. 2011, 34, 1200–1211. [Google Scholar] [CrossRef] [PubMed]
- Trombetta, E.S.; Simons, J.F.; Helenius, A. Endoplasmic reticulum glucosidase II is composed of a catalytic subunit, conserved from yeast to mammals, and a tightly bound noncatalytic HDEL-containing subunit. J. Biol. Chem. 1996, 271, 27509–27516. [Google Scholar] [CrossRef] [PubMed]
- Arendt, C.W.; Ostergaard, H.L. Two distinct domains of the beta-subunit of glucosidase II interact with the catalytic alpha-subunit. Glycobiology 2000, 10, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Ciesla, J. Metabolic enzymes that bind RNA: Yet another level of cellular regulatory network? Acta Biochim. Polonica 2006, 53, 11–32. [Google Scholar]
- Sirover, M.A. New nuclear functions of the glycolytic protein, glyceraldehyde-3-phosphate dehydrogenase, in mammalian cells. J. Cell. Biochem. 2005, 95, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yi, X.; Stoffer, J.B.; Bonafe, N.; Gilmore-Hebert, M.; McAlpine, J.; Chambers, S.K. The multifunctional protein glyceraldehyde-3-phosphate dehydrogenase is both regulated and controls colony-stimulating factor-1 messenger RNA stability in ovarian cancer. Mol. Cancer Res. 2008, 6, 1375–1384. [Google Scholar] [CrossRef] [PubMed]
- Scherrer, T.; Mittal, N.; Janga, S.C.; Gerber, A.P. A screen for RNA-binding proteins in yeast indicates dual functions for many enzymes. PLoS ONE 2010, 5, e15499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerke, V.; Moss, S.E. Annexins: From structure to function. Physiol. Rev. 2002, 82, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Filipenko, N.R.; MacLeod, T.J.; Yoon, C.S.; Waisman, D.M. Annexin A2 is a novel RNA-binding protein. J. Biol. Chem. 2004, 279, 8723–8731. [Google Scholar] [CrossRef] [PubMed]
- Merrick, E.L.; Hodgkin, D.; Garnick, D.W.; Horgan, C.M.; Panas, L.; Ryan, M.; Saitz, R.; Blow, F.C. Unhealthy drinking patterns and receipt of preventive medical services by older adults. J. Gen. Intern. Med. 2008, 23, 1741–1748. [Google Scholar] [CrossRef] [PubMed]
- Harper, C.; Matsumoto, I. Ethanol and brain damage. Curr. Opin. Pharmacol. 2005, 5, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Mayfield, J.; Ferguson, L.; Harris, R.A. Neuroimmune signaling: A key component of alcohol abuse. Curr. Opin. Neurobiol. 2013, 23, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, A.; Kirchhoff, F.; Helmchen, F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science 2005, 308, 1314–1318. [Google Scholar] [CrossRef] [PubMed]
- Dheen, S.T.; Kaur, C.; Ling, E.A. Microglial activation and its implications in the brain diseases. Curr. Med. Chem. 2007, 14, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, H.; Higashitsuji, H.; Yokoi, H.; Itoh, K.; Danno, S.; Matsuda, T.; Fujita, J. Cloning and characterization of human CIRP (cold-inducible RNA-binding protein) cDNA and chromosomal assignment of the gene. Gene 1997, 204, 115–120. [Google Scholar] [CrossRef]
- Nishiyama, H.; Itoh, K.; Kaneko, Y.; Kishishita, M.; Yoshida, O.; Fujita, J. A glycine-rich RNA-binding protein mediating cold-inducible suppression of mammalian cell growth. J. Cell Biol. 1997, 137, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Rajayer, S.R.; Jacob, A.; Yang, W.L.; Zhou, M.; Chaung, W.; Wang, P. Cold-inducible RNA-binding protein is an important mediator of alcohol-induced brain inflammation. PLoS ONE 2013, 8, e79430. [Google Scholar] [CrossRef] [PubMed]
- Morf, J.; Rey, G.; Schneider, K.; Stratmann, M.; Fujita, J.; Naef, F.; Schibler, U. Cold-inducible RNA-binding protein modulates circadian gene expression posttranscriptionally. Science 2012, 338, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Crocker, A.; Sehgal, A. Genetic analysis of sleep. Genes Dev. 2010, 24, 1220–1235. [Google Scholar] [CrossRef] [PubMed]
- Olney, J.W. Fetal alcohol syndrome at the cellular level. Addict. Biol. 2004, 9, 137–149, discussion 151. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.M.; Hwang, W.S.; Liew, C.C. Single pass sequencing of a unidirectional human fetal heart cDNA library to discover novel genes of the cardiovascular system. J. Mol. Cell. Cardiol. 1994, 26, 1329–1333. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.K.; Ito, T.; Mitsui, Y.; Sakaki, Y. Fluorescent differential display analysis of gene expression in apoptotic neuroblastoma cells. Gene 1998, 223, 21–31. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, H.; Han, K.; Grabowski, P.J. Region-specific alternative splicing in the nervous system: Implications for regulation by the RNA-binding protein NAPOR. RNA 2002, 8, 671–685. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.K.; Ito, T.; Tsukahara, F.; Hirai, M.; Sakaki, Y. Developmentally-regulated expression of mNapor encoding an apoptosis-induced elav-type RNA binding protein. Gene 1999, 237, 135–142. [Google Scholar] [CrossRef]
- Naha, N.; Lee, H.Y.; Naser, M.I.; Park, T.J.; Kim, S.H.; Kim, M.O. Ethanol inhibited apoptosis-related RNA binding protein, NAPOR-3 gene expression in the prenatal rat brain. Med. Sci. Rev. 2009, 15, BR6–BR12. [Google Scholar]
- Campos, A.R.; Grossman, D.; White, K. Mutant alleles at the locus elav in Drosophila melanogaster lead to nervous system defects. A developmental-genetic analysis. J. Neurogenet. 1985, 2, 197–218. [Google Scholar] [CrossRef] [PubMed]
- Hinman, M.N.; Lou, H. Diverse molecular functions of Hu proteins. Cell. Mol. Life Sci. 2008, 65, 3168–3181. [Google Scholar] [CrossRef] [PubMed]
- Bradley, J.R. TNF-mediated inflammatory disease. J. Pathol. 2008, 214, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Dean, J.L.; Wait, R.; Mahtani, K.R.; Sully, G.; Clark, A.R.; Saklatvala, J. The 3' untranslated region of tumor necrosis factor alpha mRNA is a target of the mRNA-stabilizing factor HuR. Mol. Cell. Biol. 2001, 21, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Kishore, R.; McMullen, M.R.; Cocuzzi, E.; Nagy, L.E. Lipopolysaccharide-mediated signal transduction: Stabilization of TNF-alpha mRNA contributes to increased lipopolysaccharide-stimulated TNF-alpha production by kupffer cells after chronic ethanol feeding. Comp. Hepatol. 2004. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Shyu, A.B. Au-rich elements: Characterization and importance in mRNA degradation. Trends Biochem. Sci. 1995, 20, 465–470. [Google Scholar] [CrossRef]
- Gruber, A.R.; Fallmann, J.; Kratochvill, F.; Kovarik, P.; Hofacker, I.L. Aresite: A database for the comprehensive investigation of AU-rich elements. Nucleic Acids Res. 2011, 39, D66–D69. [Google Scholar] [CrossRef] [PubMed]
- Orman, E.S.; Odena, G.; Bataller, R. Alcoholic liver disease: Pathogenesis, management, and novel targets for therapy. J. Gastroenterol. Hepatol. 2013, 28, 77–84. [Google Scholar] [CrossRef] [PubMed]
- McMullen, M.R.; Cocuzzi, E.; Hatzoglou, M.; Nagy, L.E. Chronic ethanol exposure increases the binding of hur to the TNFalpha 3'-untranslated region in macrophages. J. Biol. Chem. 2003, 278, 38333–38341. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.N.; Lakshman, M.R. Chronic ethanol downregulates Gal-beta-1,4GlcNAc alpha2,6-sialyltransferase and Gal-beta-1,3GlcNAc alpha2,3-sialyltransferase mRNAs in rat liver. Alcohol. Clin. Exp. Res. 1997, 21, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Garige, M.; Gong, M.; Lakshman, M.R. Ethanol destabilizes liver Gal-beta-l,4GlcNAc alpha2,6-sialyltransferase, mRNA by depleting a 3'-untranslated region-specific binding protein. J. Pharmacol. Exp. Ther. 2006, 318, 1076–1082. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Garige, M.; Hirsch, K.; Lakshman, M.R. Liver Gal-beta-1,4GlcNAc alpha2,6-sialyltransferase is down-regulated in human alcoholics: Possible cause for the appearance of asialoconjugates. Metabolism 2007, 56, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Lukong, K.E.; Chang, K.W.; Khandjian, E.W.; Richard, S. RNA-binding proteins in human genetic disease. Trends Genet. 2008, 24, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Hermann, T.; Westhof, E. Rational drug design and high-throughput techniques for RNA targets. Comb. Chem. High Throughput Screen. 2000, 3, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Cheneval, D.; Kastelic, T.; Fuerst, P.; Parker, C.N. A review of methods to monitor the modulation of mRNA stability: A novel approach to drug discovery and therapeutic intervention. J. Biomol. Screen. 2010, 15, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Yeo, G.; Holste, D.; Kreiman, G.; Burge, C.B. Variation in alternative splicing across human tissues. Genome Biol. 2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, C.E.; Bullock, S.L. Subcellular mRNA localization in animal cells and why it matters. Science 2009, 326, 1212–1216. [Google Scholar] [CrossRef] [PubMed]
- Szabo, A.; Dalmau, J.; Manley, G.; Rosenfeld, M.; Wong, E.; Henson, J.; Posner, J.B.; Furneaux, H.M. HuD, a paraneoplastic encephalomyelitis antigen, contains RNA-binding domains and is homologous to elav and sex-lethal. Cell 1991, 67, 325–333. [Google Scholar] [CrossRef]
- Pignolet, B.S.; Gebauer, C.M.; Liblau, R.S. Immunopathogenesis of paraneoplastic neurological syndromes associated with anti-Hu antibodies: A beneficial antitumor immune response going awry. Oncoimmunology 2013, 2, e27384. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Guo, Y.; Chu, H.; Guan, Y.; Bi, J.; Wang, B. Multiple functions of the RNA-binding protein HuR in cancer progression, treatment responses and prognosis. Int. J. Mol. Sci. 2013, 14, 10015–10041. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Mangelsdorf, M.; Liu, J.; Zhu, L.; Wu, J. RNA-binding proteins in neurological diseases. Sci. China Life Sci. 2014, 57, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Darnell, R.B.; Posner, J.B. Paraneoplastic syndromes involving the nervous system. N. Engl. J. Med. 2003, 349, 1543–1554. [Google Scholar] [CrossRef] [PubMed]
- Buckanovich, R.J.; Posner, J.B.; Darnell, R.B. Nova, the paraneoplastic Ri antigen, is homologous to an RNA-binding protein and is specifically expressed in the developing motor system. Neuron 1993, 11, 657–672. [Google Scholar] [CrossRef]
- Ule, J.; Jensen, K.B.; Ruggiu, M.; Mele, A.; Ule, A.; Darnell, R.B. CLIP identifies Nova-regulated RNA networks in the brain. Science 2003, 302, 1212–1215. [Google Scholar] [CrossRef] [PubMed]
- Dredge, B.K.; Darnell, R.B. Nova regulates GABA(A) receptor gamma2 alternative splicing via a distal downstream UCAU-rich intronic splicing enhancer. Mol. Cell. Biol. 2003, 23, 4687–4700. [Google Scholar] [CrossRef] [PubMed]
- Jensen, K.B.; Musunuru, K.; Lewis, H.A.; Burley, S.K.; Darnell, R.B. The tetranucleotide UCAY directs the specific recognition of RNA by the Nova K-homology 3 domain. Proc. Natl. Acad. Sci. USA 2000, 97, 5740–5745. [Google Scholar] [CrossRef] [PubMed]
- Bolognani, F.; Perrone-Bizzozero, N.I. RNA-protein interactions and control of mRNA stability in neurons. J. Neurosci. Res. 2008, 86, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Akamatsu, W.; Fujihara, H.; Mitsuhashi, T.; Yano, M.; Shibata, S.; Hayakawa, Y.; Okano, H.J.; Sakakibara, S.; Takano, H.; Takano, T.; et al. The RNA-binding protein HuD regulates neuronal cell identity and maturation. Proc. Natl. Acad. Sci. USA 2005, 102, 4625–4630. [Google Scholar] [CrossRef] [PubMed]
- Bolognani, F.; Qiu, S.; Tanner, D.C.; Paik, J.; Perrone-Bizzozero, N.I.; Weeber, E.J. Associative and spatial learning and memory deficits in transgenic mice overexpressing the RNA-binding protein HuD. Neurobiol. Learn. Mem. 2007, 87, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Roux, P.P.; Topisirovic, I. Regulation of mRNA translation by signaling pathways. Cold Spring Harb. Perspect. Biol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Costa-Mattioli, M.; Sossin, W.S.; Klann, E.; Sonenberg, N. Translational control of long-lasting synaptic plasticity and memory. Neuron 2009, 61, 10–26. [Google Scholar] [CrossRef] [PubMed]
- Kidd, S.A.; Lachiewicz, A.; Barbouth, D.; Blitz, R.K.; Delahunty, C.; McBrien, D.; Visootsak, J.; Berry-Kravis, E. Fragile X syndrome: A review of associated medical problems. Pediatrics 2014, 134, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.; Joseph, S. Fragile X mental retardation protein: A paradigm for translational control by RNA-binding proteins. Biochimie 2015, 114, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Darnell, J.C.; van Driesche, S.J.; Zhang, C.; Hung, K.Y.; Mele, A.; Fraser, C.E.; Stone, E.F.; Chen, C.; Fak, J.J.; Chi, S.W.; et al. FMRP stalls ribosomal translocation on mRNAs linked to synaptic function and autism. Cell 2011, 146, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Gatto, C.L.; Broadie, K. The fragile X mental retardation protein in circadian rhythmicity and memory consolidation. Mol. Neurobiol. 2009, 39, 107–129. [Google Scholar] [CrossRef] [PubMed]
- Ifrim, M.F.; Williams, K.R.; Bassell, G.J. Single-molecule imaging of PSD-95 mRNA translation in dendrites and its dysregulation in a mouse model of fragile X syndrome. J. Neurosci. 2015, 35, 7116–7130. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Sul, J.Y.; Helmfors, H.; Langel, U.; Kim, J.; Eberwine, J. Dendritic glutamate receptor mRNAs show contingent local hotspot-dependent translational dynamics. Cell Rep. 2013, 5, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.C.; Zukin, R.S. RNA trafficking and local protein synthesis in dendrites: An overview. J. Neurosci. 2006, 26, 7131–7134. [Google Scholar] [CrossRef] [PubMed]
- Bramham, C.R.; Wells, D.G. Dendritic mRNA: Transport, translation and function. Nat. Rev. Neurosci. 2007, 8, 776–789. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.P. Synaptic plasticity: Regulated translation in dendrites. Curr. Biol. 1999, 9, R168–R170. [Google Scholar] [CrossRef]
- Batish, M.; van den Bogaard, P.; Kramer, F.R.; Tyagi, S. Neuronal mRNAs travel singly into dendrites. Proc. Natl. Acad. Sci. USA 2012, 109, 4645–4650. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y.; Hur, J.; Jeong, S. Emerging roles of RNA and RNA-binding protein network in cancer cells. BMB Rep. 2009, 42, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Kechavarzi, B.; Janga, S.C. Dissecting the expression landscape of RNA-binding proteins in human cancers. Genome Biol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Busa, R.; Paronetto, M.P.; Farini, D.; Pierantozzi, E.; Botti, F.; Angelini, D.F.; Attisani, F.; Vespasiani, G.; Sette, C. The RNA-binding protein Sam68 contributes to proliferation and survival of human prostate cancer cells. Oncogene 2007, 26, 4372–4382. [Google Scholar] [CrossRef] [PubMed]
- Lukong, K.E.; Larocque, D.; Tyner, A.L.; Richard, S. Tyrosine phosphorylation of Sam68 by breast tumor kinase regulates intranuclear localization and cell cycle progression. J. Biol. Chem. 2005, 280, 38639–38647. [Google Scholar] [CrossRef] [PubMed]
- Naor, D.; Wallach-Dayan, S.B.; Zahalka, M.A.; Sionov, R.V. Involvement of CD44, a molecule with a thousand faces, in cancer dissemination. Semin. Cancer Biol. 2008, 18, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Alao, J.P. The regulation of cyclin D1 degradation: Roles in cancer development and the potential for therapeutic invention. Mol. Cancer 2007, 6, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Frisone, P.; Pradella, D.; di Matteo, A.; Belloni, E.; Ghigna, C.; Paronetto, M.P. Sam68: Signal transduction and RNA metabolism in human cancer. Biomed. Res. Int. 2015. [Google Scholar] [CrossRef] [PubMed]
- Wurth, L.; Gebauer, F. RNA-binding proteins, multifaceted translational regulators in cancer. Biochim. Biophys. Acta 2015, 1849, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, A.C.; Ruggero, D. Targeting eukaryotic translation initiation factor 4E (eIF4E) in cancer. Clin. Cancer Res. 2010, 16, 4914–4920. [Google Scholar] [CrossRef] [PubMed]
- Budowsky, E.I.; Simukova, N.A.; Turchinsky, M.F.; Boni, I.V.; Skoblov, Y.M. Induced formation of covalent bonds between nucleoprotein components. V. UV or bisulfite induced polynucleotide-protein crosslinkage in bacteriophage MS2. Nucleic Acids Res. 1976, 3, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, J.R. Ultraviolet light-induced crosslinking of mRNA to proteins. Nucleic Acids Res. 1979, 6, 715–732. [Google Scholar] [CrossRef] [PubMed]
- Ule, J.; Jensen, K.; Mele, A.; Darnell, R.B. CLIP: A method for identifying protein-RNA interaction sites in living cells. Methods 2005, 37, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Licatalosi, D.D.; Mele, A.; Fak, J.J.; Ule, J.; Kayikci, M.; Chi, S.W.; Clark, T.A.; Schweitzer, A.C.; Blume, J.E.; Wang, X.; et al. HITS-CLIP yields genome-wide insights into brain alternative RNA processing. Nature 2008, 456, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Cook, K.B.; Kazan, H.; Zuberi, K.; Morris, Q.; Hughes, T.R. RBPDB: A database of RNA-binding specificities. Nucleic Acids Res. 2011, 39, D301–D308. [Google Scholar] [CrossRef] [PubMed]
- Tamburino, A.M.; Ryder, S.P.; Walhout, A.J. A compendium of Caenorhabditis elegans RNA binding proteins predicts extensive regulation at multiple levels. G3 2013, 3, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Hogan, D.J.; Riordan, D.P.; Gerber, A.P.; Herschlag, D.; Brown, P.O. Diverse RNA-binding proteins interact with functionally related sets of RNAs, suggesting an extensive regulatory system. PLoS Biol. 2008, 6, e255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Xiao, G.; Chu, Y.; Zhang, M.Q.; Corey, D.R.; Xie, Y. Design and bioinformatics analysis of genome-wide CLIP experiments. Nucleic Acids Res. 2015, 43, 5263–5274. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Yun, J.; Kim, M.S.; Mendell, J.T.; Xie, Y. Pipe-CLIP: A comprehensive online tool for CLIP-seq data analysis. Genome Biol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Janga, S.C. From specific to global analysis of posttranscriptional regulation in eukaryotes: Posttranscriptional regulatory networks. Brief. Funct. Genomics 2012, 11, 505–521. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anji, A.; Kumari, M. Guardian of Genetic Messenger-RNA-Binding Proteins. Biomolecules 2016, 6, 4. https://doi.org/10.3390/biom6010004
Anji A, Kumari M. Guardian of Genetic Messenger-RNA-Binding Proteins. Biomolecules. 2016; 6(1):4. https://doi.org/10.3390/biom6010004
Chicago/Turabian StyleAnji, Antje, and Meena Kumari. 2016. "Guardian of Genetic Messenger-RNA-Binding Proteins" Biomolecules 6, no. 1: 4. https://doi.org/10.3390/biom6010004
APA StyleAnji, A., & Kumari, M. (2016). Guardian of Genetic Messenger-RNA-Binding Proteins. Biomolecules, 6(1), 4. https://doi.org/10.3390/biom6010004