A Functional Analysis of the Cyclophilin Repertoire in the Protozoan Parasite Trypanosoma Cruzi

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction to Trypanosoma cruzi Infection and Chagas Disease
2. The T. cruzi Cyclophilin Repertoire
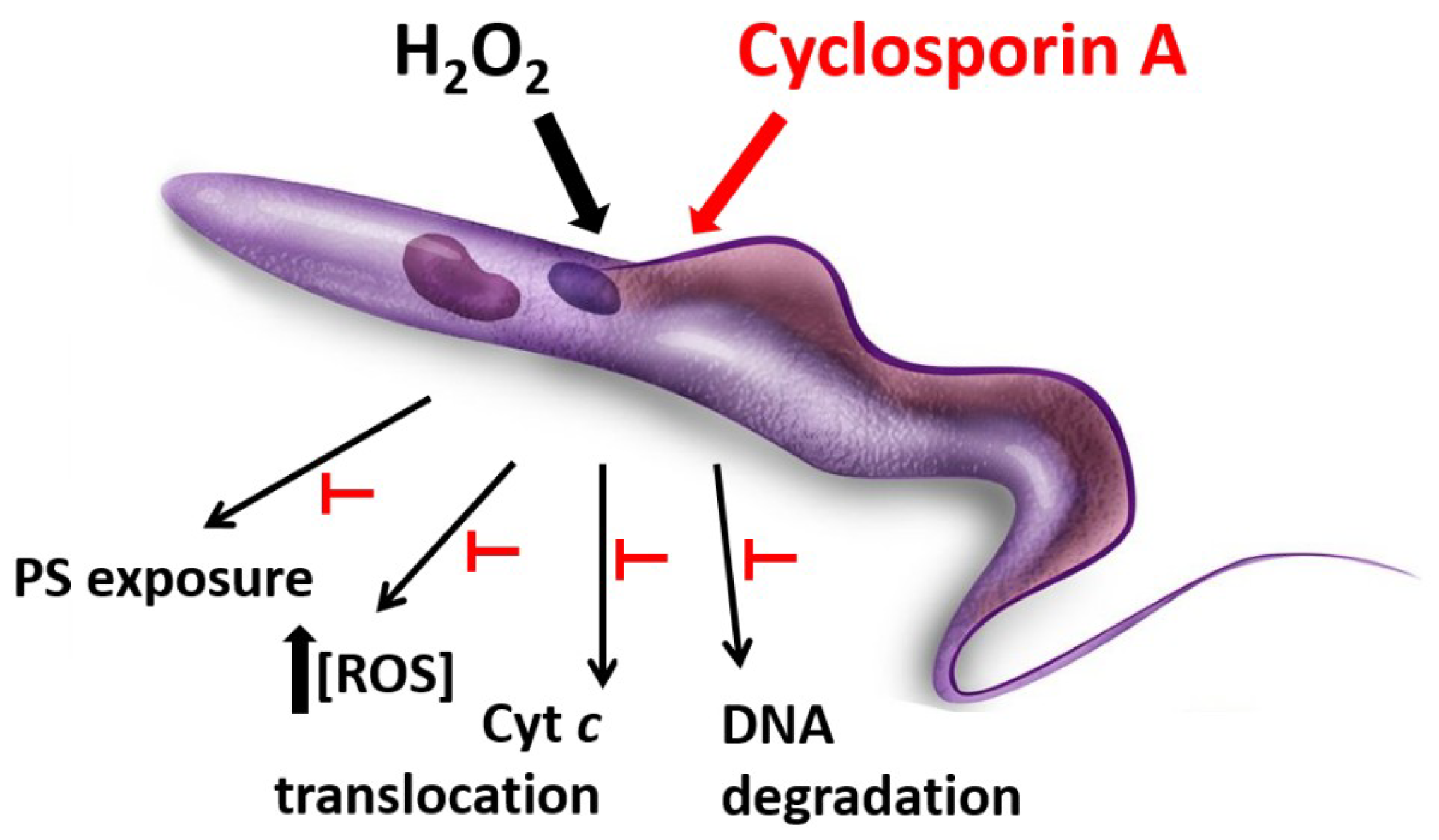
3. Cyclosporin A as Trypanocidal Agent
4. The Effects of Cyclosporin A on T. cruzi Cyclophilins
5. Functional Features of T. cruzi Peptidyl-Prolyl cis-trans Isomerases
6. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. Epidemiology; World Health Organization: Geneva, Switzerland; Available online: http://www.who.int/chagas/epidemiology/en/ (accessed on 1 October 2018).
- Yamagata, Y.; Nakagawa, J. Control of Chagas Disease. Adv. Parasitol. 2006, 61, 129–165. [Google Scholar] [PubMed]
- Bern, C.; Kjos, S.; Yabsley, M.J.; Montgomery, S.P. Trypanosoma cruzi and Chagas’ Disease in the United States. Clin. Microbiol. Rev. 2011, 24, 655–681. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Molina, J.A.; Molina, I. Chagas disease. Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef]
- Picado, A.; Cruz, I.; Redard-Jacot, M.; Schijman, A.G.; Torrico, F.; Sosa-Estani, S.; Katz, Z.; Ndung’u, J.M. The burden of congenital Chagas disease and implementation of molecular diagnostic tools in Latin America. BMJ Glob. Health 2018, 3, e001069. [Google Scholar] [CrossRef] [PubMed]
- Bua, J.; Volta, B.J.; Perrone, A.E.; Scollo, K.; Velázquez, E.B.; Ruiz, A.M.; De Rissio, A.M.; Cardoni, R.L. How to Improve the Early Diagnosis of Trypanosoma cruzi Infection: Relationship between Validated Conventional Diagnosis and Quantitative DNA Amplification in Congenitally Infected Children. PLoS Negl. Trop. Dis. 2013, 7, 12–14. [Google Scholar] [CrossRef] [PubMed]
- Carlier, Y.; Truyens, C. Congenital Chagas disease as an ecological model of interactions between Trypanosoma cruzi parasites, pregnant women, placenta and fetuses. Acta Trop. 2015, 151, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Hsu, R.C.; Burak, J.; Tiwari, S.; Chakraborti, C.; Sander, G.E. Chagas Cardiomyopathy in New Orleans and the Southeastern United States. Ochsner J. 2016, 16, 304–308. [Google Scholar] [PubMed]
- Rodriguez-Guardado, A.; González, M.L.; Rodriguez, M.; Flores-Chavez, M.; Boga, J.A.; Gascon, J. Trypanosoma cruzi infection in a Spanish liver transplant recipient. Clin. Microbiol. Infect. 2015, 21, 687.e1–687.e3. [Google Scholar] [CrossRef] [PubMed]
- Soriano-Arandes, A.; Angheben, A.; Serre-Delcor, N.; Treviño-Maruri, B.; Gómez i Prat, J.; Jackson, Y. Control and management of congenital Chagas disease in Europe and other non-endemic countries: Current policies and practices. Trop. Med. Int. Health 2016, 21, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Kratz, J.M.; Garcia Bournissen, F.; Forsyth, C.J.; Sosa-Estani, S. Clinical and pharmacological profile of benznidazole for treatment of Chagas disease. Expert Rev. Clin. Pharmacol. 2018, 11, 943–957. [Google Scholar] [CrossRef] [PubMed]
- Handschumacher, R.; Harding, M.; Rice, J.; Drugge, R. Specific Cytosolic Bidg Protein for Cyclosporin. Science 1984, 226, 544–547. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Hayano, T.; Suzuki, M. Peptidyl-prolyl cis-trans isomerase is the cyclosporin A-binding protein cyclophilin. Nature 1989, 337, 473–475. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.K.; Swedberg, J.E.; Harvey, P.J.; Kaas, Q.; Craik, D.J. Conformational Flexibility Is a Determinant of Permeability for Cyclosporin. J. Phys. Chem. B 2018, 122, 2261–2276. [Google Scholar] [CrossRef] [PubMed]
- Búa, J.; Åslund, L.; Pereyra, N.; García, G.A.; Bontempi, E.J.; Ruiz, A.M. Characterisation of a cyclophilin isoform in Trypanosoma cruzi. FEMS Microbiol. Lett. 2001, 200, 43–47. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Potenza, M.; Galat, A.; Minning, T.A.; Ruiz, A.M.; Duran, R.; Tarleton, R.L.; Marín, M.; Fichera, L.E.; Búa, J. Analysis of the Trypanosoma cruzi cyclophilin gene family and identification of Cyclosporin A binding proteins. Parasitology 2006, 132, 867–882. [Google Scholar] [CrossRef] [PubMed]
- Bustos, P.L.; Perrone, A.E. Improved Immuno-Detection of a Low-Abundance Cyclophilin Allows the Confirmation of its Expression in a Protozoan Parasite. Immunochem. Immunopathol. 2015, 1, 1–4. [Google Scholar] [CrossRef]
- Kieffers, L.J.; Thalhammere, T.; Handschumacher, E. Isolation and Characterization of a 40-kDa Cyclophilin-related Protein. J. Biol. Chem. 1992, 267, 5503–5507. [Google Scholar]
- Hoffmann, K.; Kakalis, L.T.; Anderson, K.S.; Armitage, I.M.; Handschumacher, R.E. Expression of Human Cyclophilin-40 and the Effect of the His141→Trp Mutation on Catalysis and Cyclosporin A Binding. Eur. J. Biochem. 1995, 229, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, T.; Carrello, A. Cyclophilin 40 (CyP-40), mapping of its hsp90 binding domain and evidence that FKBP52 competes with CyP-40 for hsp90 binding. J. Biol. Chem. 1996, 271, 2961–2965. [Google Scholar] [CrossRef] [PubMed]
- Yau, W.L.; Blisnick, T.; Taly, J.F.; Helmer-Citterich, M.; Schiene-Fischer, C.; Leclercq, O.; Li, J.; Schmidt-Arras, D.; Morales, M.A.; Notredame, C.; et al. Cyclosporin A treatment of Leishmania donovani reveals stage-specific functions of cyclophilins in parasite proliferation and viability. PLoS Negl. Trop. Dis. 2010, 4, e729. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Vijayakumar, S.; Schiene-Fischer, C.; Li, H.; Purkerson, J.M.; Malesevic, M.; Liebscher, J.; Al-Awqati, Q.; Schwartz, G.J. Secreted cyclophilin A, a peptidylprolyl cis-trans isomerase, mediates matrix assembly of Hensin, a protein implicated in epithelial differentiation. J. Biol. Chem. 2009, 284, 6465–6475. [Google Scholar] [CrossRef] [PubMed]
- Pellé, R.; McOdimba, F.; Chuma, F.; Wasawo, D.; Pearson, T.W.; Murphy, N.B. The African trypanosome cyclophilin A homologue contains unusual conserved central and N-terminal domains and is developmentally regulated. Gene 2002, 290, 181–191. [Google Scholar] [CrossRef]
- Aliberti, J.; Valenzuela, J.G.; Carruthers, V.B.; Hieny, S.; Andersen, J.; Charest, H.; Sousa, C.R.; Fairlamb, A.; Ribeiro, J.M.; Sher, A. Molecular mimicry of a CCR5 binding-domain in the microbial activation of dendritic cells. Nat. Immunol. 2003, 4, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, M.M.; Karafova, A.; Kamysz, W.; Schenkman, S.; Pelle, R.; McGwire, B.S. Secreted trypanosome cyclophilin inactivates lytic insect defense peptides and induces parasite calcineurin activation and infectivity. J. Biol. Chem. 2013, 288, 8772–8784. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, J.D.; Jensen, L.J.; Blom, N.; Von Heijne, G.; Brunak, S. Feature-based prediction of non-classical and leaderless protein secretion. Protein Eng. Des. Sel. 2004, 17, 349–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, H.; Schiene-Fischer, C. Functional aspects of extracellular cyclophilins. Biol. Chem. 2014, 395, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Solbach, W.; Forberg, K.; Kammerer, E.; Bogdan, C.; Röllinghoff, M. Suppressive effect of cyclosporin A on the development of Leishmania tropica-induced lesions in genetically susceptible BALB/c mice. J. Immunol. 1986, 137, 702–707. [Google Scholar] [PubMed]
- Chappell, L.H.; Thomson, A.W.; Barker, G.C.; Smith, S.W.G. Dosage, timing, and route of administration of cyclosporin A and nonimmunosuppressive derivatives of dihydrocyclosporin A and cyclosporin C against Schistosoma mansoni in vivo and in vitro. Antimicrob. Agents Chemother. 1987, 31, 1567–1571. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.; Wernli, B.; Franklin, R.M. Roles of peptidyl-prolyl CIS-trans isomerase and calcineurin in the mechanisms of antimalarial action of cyclosporin a, FK506, and rapamycin. Biochem. Pharmacol. 1994, 48, 495–503. [Google Scholar] [CrossRef]
- Galat, A.; Bua, J. Molecular aspects of cyclophilins mediating therapeutic actions of their ligands. Cell. Mol. Life Sci. 2010, 67, 3467–3488. [Google Scholar] [CrossRef] [PubMed]
- Page, A.P.; Kumar, S.; Carlow, C.K.S. Parasite cyclophilins and antiparasite activity of cyclosporin A. Parasitol. Today 1995, 11, 385–388. [Google Scholar] [CrossRef]
- Bell, A.; Monaghan, P.; Page, A.P. Peptidyl-prolyl cis-trans isomerases (immunophilins) and their roles in parasite biochemistry, host-parasite interaction and antiparasitic drug action. Int. J. Parasitol. 2006, 36, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Chappell, L.H.; Wastling, J.M. Cyclosporin A: Antiparasite drug, modulator of the host-parasite relationship and immunosuppressant. Parasitology 1992, 105, S25–S40. [Google Scholar] [CrossRef] [PubMed]
- Bua, J.; Fichera, L.E.; Fuchs, A.G.; Potenza, M.; Dubin, M.; Wenger, R.O.; Moretti, G.; Scabone, C.M.; Ruiz, A.M. Anti-Trypanosoma cruzi effects of cyclosporin A derivatives: Possible role of a P-glycoprotein and parasite cyclophilins. Parasitology 2008, 135, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Garavaglia, P.A.; Laverrière, M.; Cannata, J.J.B.; García, G.A. Putative role of the aldo-keto reductase from Trypanosoma cruzi in benznidazole metabolism. Antimicrob. Agents Chemother. 2016, 60, 2664–2670. [Google Scholar] [CrossRef] [PubMed]
- Luna-Marín, K.P.; Hernández, I.P.; Rueda, C.M.; Zorro, M.M.; Croft, S.L.; Escobar, P. In vitro susceptibility of Trypanosoma cruzi strains from Santander, Colombia, to hexadecylphosphocholine (miltefosine), nifurtimox and benznidazole. Biomédica 2009, 29, 448–455. [Google Scholar]
- McCabe, R.E.; Remington, J.S.; Araujo, F.G. In Vivo and in Vitro Effects of Cyclosporin A on Trypanosoma cruzi. Am. J. Trop. Med. Hyg. 1985, 34, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Bua, J.; Ruiz, A.M.; Potenza, M.; Fichera, L.E. In vitro anti-parasitic activity of Cyclosporin A analogs on Trypanosoma cruzi. Bioorg. Med. Chem. Lett. 2004, 14, 4633–4637. [Google Scholar] [CrossRef] [PubMed]
- Fischer, G.; Bang, H.; Mech, C. Nachweis einer Enzymkatalyse für die cis-trans-Isomerisierung der Peptidbindung in prolinhaltigen Peptiden. Biomed. Biochim. Acta 1984, 43, 1101–1111. [Google Scholar] [PubMed]
- Carraro, R.; Bua, J.; Ruiz, A.; Paulino, M. Modelling and study of cyclosporin A and related compounds in complexes with a Trypanosoma cruzi cyclophilin. J. Mol. Graph. Model. 2007, 26, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Berriman, M.; Fairlamb, A.H. Detailed characterization of a cyclophilin from the human malaria parasite Plasmodium falciparum. Biochem. J. 1998, 334, 437–445. [Google Scholar] [CrossRef] [PubMed]
- High, K.P. The antimicrobial activities of cyclosporine, FK506, and rapamycin. Transplantation 1994, 57, 1689–1700. [Google Scholar] [CrossRef] [PubMed]
- Crompton, M.; Virji, S.; Ward, J.M. Cyclophilin-D binds strongly to complexes of the voltage-dependent anion channel and the adenine nucleotide translocase to form the permeability transition pore. Eur. J. Biochem. 1998, 258, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Baines, C.P.; Kaiser, R.A.; Purcell, N.H.; Blair, N.S.; Osinska, H.; Hambleton, M.A.; Brunskill, E.W.; Sayen, M.R.; Gottlieb, R.A.; Dorn, G.W.; et al. Loss of cyclophilin D reveals a critical role for mitochondrial permeability transition in cell death. Nature 2005, 434, 658–662. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Wang, X.; Deinum, J.; Huang, Z.; Gao, J.; Modjtahedi, N.; Neagu, M.R.; Nilsson, M.; Eriksson, P.S.; Hagberg, H.; et al. Cyclophilin A participates in the nuclear translocation of apoptosis-inducing factor in neurons after cerebral hypoxia-ischemia. J. Exp. Med. 2007, 204, 1741–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustos, P.L.; Perrone, A.E.; Milduberger, N.; Postan, M.; Bua, J. Oxidative stress damage in the protozoan parasite Trypanosoma cruzi is inhibited by Cyclosporin A. Parasitology 2015, 142, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Elmahallawy, E.K.; Jiménez-Aranda, A.; Martínez, A.S.; Rodriguez-Granger, J.; Navarro-Alarcón, M.; Gutiérrez-Fernández, J.; Agil, A. Activity of melatonin against Leishmania infantum promastigotes by mitochondrial dependent pathway. Chem. Biol. Interact. 2014, 220, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Rêgo, J.V.; Duarte, A.P.; Liarte, D.B.; de Carvalho Sousa, F.; Barreto, H.M.; Bua, J.; Romanha, A.J.; Rádis-Baptista, G.; Murta, S.M.F. Molecular characterization of Cyclophilin (TcCyP19) in Trypanosoma cruzi populations susceptible and resistant to benznidazole. Exp. Parasitol. 2015, 148, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, A.; Westermann, B.; Osborn, M.; Nordheim, A. The N-terminal signal peptide of the murine cyclophilin mCyP-S1 is required in vivo for ER localization. Eur. J. Cell. Biol. 1994, 63, 182–191. [Google Scholar] [PubMed]
- Bustos, P.L.; Volta, B.J.; Perrone, A.E.; Milduberger, N.; Bua, J. A homolog of cyclophilin D is expressed in Trypanosoma cruzi and is involved in the oxidative stress–damage response. Cell Death Discov. 2017, 3, 16092. [Google Scholar] [CrossRef] [PubMed]
- Bustos, P.L.; Perrone, A.E.; Milduberger, N.A.; Bua, J. Mitochondrial permeability transition in protozoan parasites: what we learned from Trypanosoma cruzi. Cell Death Dis. 2017, 8, e3057. [Google Scholar] [CrossRef] [PubMed]
- Matrangolo, F.S.V.; Liarte, D.B.; Andrade, L.C.; De Melo, M.F.; Andrade, J.M.; Ferreira, R.F.; Santiago, A.S.; Pirovani, C.P.; Silva-Pereira, R.A.; Murta, S.M.F. Comparative proteomic analysis of antimony-resistant and-susceptible Leishmania braziliensis and Leishmania infantum chagasi lines. Mol. Biochem. Parasitol. 2013, 190, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Das, I.; Datta, R.; Sen, B.; Bhattacharyya, D.; Mandal, C.; Datta, A.K. A single-domain cyclophilin from Leishmania donovani reactivates soluble aggregates of adenosine kinase by isomerase-independent chaperone function. J. Biol. Chem. 2002, 277, 47451–47460. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, D.; Patra, H.; Laskar, A.; Dasgupta, A.; Maiti, N.C.; Datta, A.K. Cyclophilin-mediated reactivation pathway of inactive adenosine kinase aggregates. Arch. Biochem. Biophys. 2013, 537, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Golding, H.; Aliberti, J.; King, L.R.; Manischewitz, J.; Andersen, J.; Valenzuela, J.; Landau, N.R.; Sher, A. Inhibition of HIV-1 infection by a CCR5-binding cyclophilin from Toxoplasma gondii. Blood 2003, 102, 3280–3286. [Google Scholar] [CrossRef] [PubMed]
- Yau, W.L.; Pescher, P.; Macdonald, A.; Hem, S.; Zander, D.; Retzlaff, S.; Blisnick, T.; Rotureau, B.; Rosenqvist, H.; Wiese, M.; et al. The Leishmania donovani chaperone cyclophilin 40 is essential for intracellular infection independent of its stage-specific phosphorylation status. Mol. Microbiol. 2014, 93, 80–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yau, W.L.; Lambertz, U.; Colineau, L.; Pescher, P.; MacDonald, A.; Zander, D.; Retzlaff, S.; Eick, J.; Reiner, N.E.; Clos, J.; et al. Phenotypic Characterization of a Leishmania donovani Cyclophilin 40 Null Mutant. J. Eukaryot. Microbiol. 2016, 63, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.R. Cloning and characterization of a Plasmodium falciparum cyclophilin gene that is stage-specifically expressed. Mol. Biochem. Parasitol. 1995, 73, 111–121. [Google Scholar] [CrossRef]
- Krücken, J.; Greif, G.; Von Samson-Himmelstjerna, G. In silico analysis of the cyclophilin repertoire of apicomplexan parasites. Parasit. Vectors 2009, 2, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Rascher, C.; Pahl, A.; Pecht, A.; Brune, K.; Solbach, W.; Bang, H. Leishmania major parasites express cyclophilin isoforms with an unusual interaction with calcineurin. Biochem. J. 1998, 334, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Dutta, M.; Delhi, P.; Sinha, K.M.; Banerjee, R.; Datta, A.K. Lack of Abundance of Cytoplasmic Cyclosporin A-binding Protein Renders Free-living Leishmania donovani Resistant to Cyclosporin A. J. Biol. Chem. 2001, 276, 19294–19300. [Google Scholar] [CrossRef] [PubMed] [Green Version]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perrone, A.E.; Milduberger, N.; Fuchs, A.G.; Bustos, P.L.; Bua, J. A Functional Analysis of the Cyclophilin Repertoire in the Protozoan Parasite Trypanosoma Cruzi. Biomolecules 2018, 8, 132. https://doi.org/10.3390/biom8040132
Perrone AE, Milduberger N, Fuchs AG, Bustos PL, Bua J. A Functional Analysis of the Cyclophilin Repertoire in the Protozoan Parasite Trypanosoma Cruzi. Biomolecules. 2018; 8(4):132. https://doi.org/10.3390/biom8040132
Chicago/Turabian StylePerrone, Alina E., Natalia Milduberger, Alicia G. Fuchs, Patricia L. Bustos, and Jacqueline Bua. 2018. "A Functional Analysis of the Cyclophilin Repertoire in the Protozoan Parasite Trypanosoma Cruzi" Biomolecules 8, no. 4: 132. https://doi.org/10.3390/biom8040132
APA StylePerrone, A. E., Milduberger, N., Fuchs, A. G., Bustos, P. L., & Bua, J. (2018). A Functional Analysis of the Cyclophilin Repertoire in the Protozoan Parasite Trypanosoma Cruzi. Biomolecules, 8(4), 132. https://doi.org/10.3390/biom8040132