Regulation of Cellular Metabolism through Phase Separation of Enzymes

Abstract
:1. Regulation of Metabolite Flux
2. Metabolism-Related Enzyme Condensates in Yeast and Other Organisms
3. Phase Separation as a Means to Acutely Regulate Enzymatic Activity
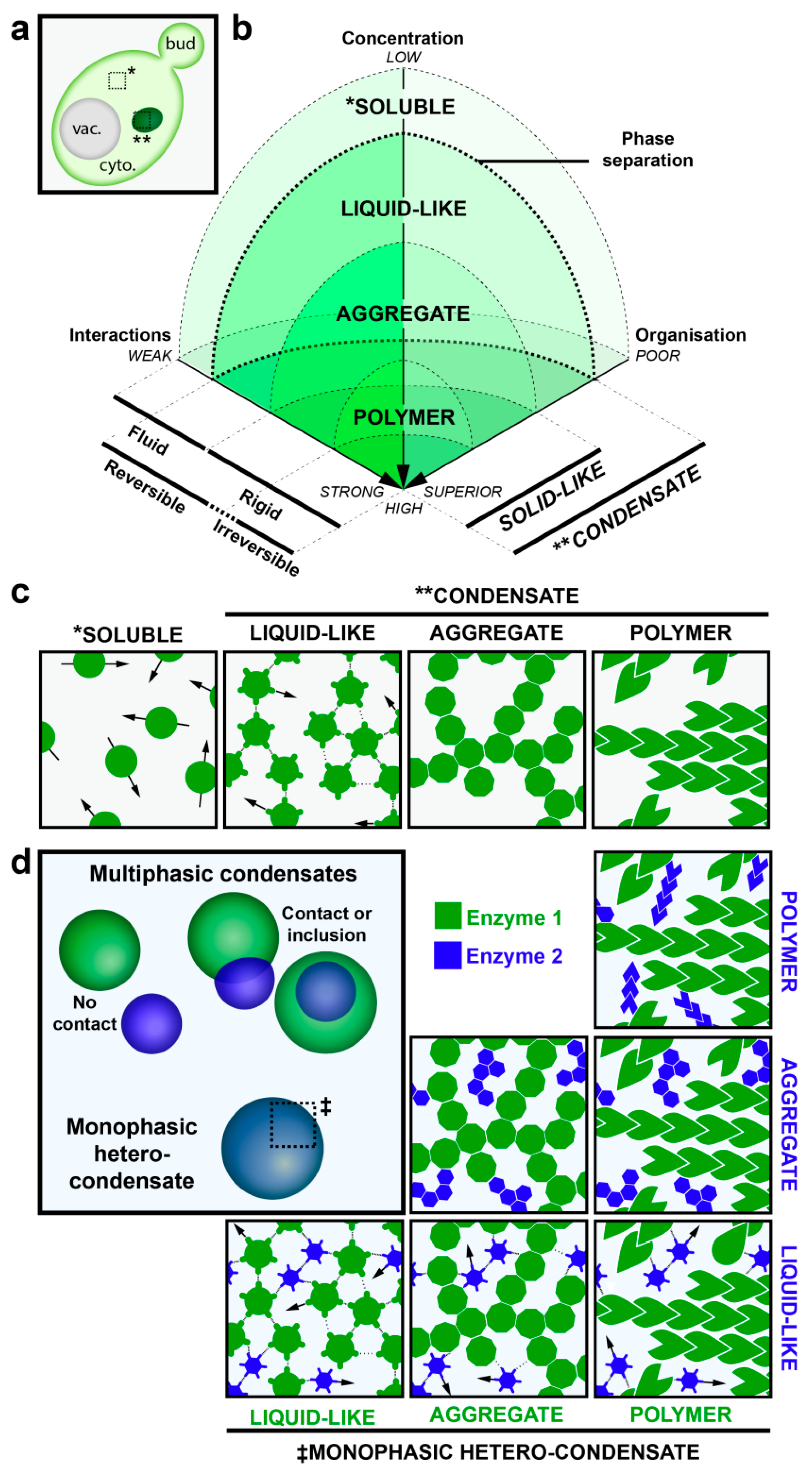
- Liquid Droplets: The phase separation continuum is illustrated in Figure 1. Condensates referred to as liquid droplets are composed of one or more factors that self-associate through numerous, weak interactions. Consequently, the factors within these condensates are highly mobile and can be rapidly exchanged with the soluble phase as assessed by fluorescence recovery after photobleaching. RNA- and protein-containing P-granules are the prototypic example of this type of condensate. The proteins that form liquid droplets typically contain domains of low sequence complexity known as inherently disordered regions.
- Amyloid-like aggregates: Further concentration of these proteins, combined with partial unfolding of the inherently disordered regions, can lead to rigidification of the condensate into what can be referred to as an aggregate. Relative to liquid droplets, proteins in aggregates are held together by stronger β-strand-like contacts that form amyloid fiber-like structures. In amyloid-fiber-like condensates/aggregates, proteins are much less mobile and typically present only a partially organized structure (Figure 1). Cdc19 condensates, described above, have been proposed to be reversible amyloid-like aggregates [11], while other amyloid-fiber-like aggregates appear to be more stable and even engaged in irreversible, potentially pathogenic aggregates that can only be cleared through degradative pathways [45,46,47].
- Polymers: Electron microscopy (EM) studies initiated in the 1970s revealed that many metabolic enzymes form filaments and/or helices [20,29,30,36,37,39,40,41,48]. These are typically formed upon exposure to an extrinsic stimulus, suggesting that enzyme polymerization represents an underappreciated mechanism by which cells regulate enzymatic activity and metabolic homeostasis. Polymers thus potentially represent another type of rigid protein condensate found in cells. Unlike aggregates, polymers would present a well-defined structure that can be readily disassembled in vivo. Indeed, rapid assembly/disassembly kinetics would endow upon this type of condensate the potential to acutely regulate enzymatic activity (Figure 1). We highlight now a selection of metabolic enzymes that appear to be regulated through phase separation into polymers.
3.1. Examples of Phase Separation in Carbohydrate Metabolism
3.2. Examples of Phase Separation in Nucleotide Metabolism
3.3. Examples of Phase Separation in Fatty Acid Metabolism
3.4. Examples of Phase Separation in Amino Acid Metabolism
4. Metabolism-Related Signaling Enzymes Also Polymerize/Coalesce into Foci
5. Outlook
Funding
Conflicts of Interest
References
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O.; Posener, K.; Negelein, E. On the metabolism of carcinoma cells. Biochem. Z. 1924, 152, 309–344. [Google Scholar] [CrossRef]
- Israelsen, W.J.; Vander Heiden, M.G. Pyruvate kinase: Function, regulation and role in cancer. Semin. Cell Dev. Biol. 2015, 43, 43–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, M.; Sato, T.; Nomura, M.; Sakamoto, Y.; Inoue, Y.; Tanaka, R.; Ito, S.; Kurosawa, K.; Yamaguchi, K.; Sugiura, Y.; et al. PKM1 confers metabolic advantages and promotes cell-autonomous tumor cell growth. Cancer Cell 2018, 33, 355–367 e357. [Google Scholar] [CrossRef] [PubMed]
- Boles, E.; Schulte, F.; Miosga, T.; Freidel, K.; Schluter, E.; Zimmermann, F.K.; Hollenberg, C.P.; Heinisch, J.J. Characterization of a glucose-repressed pyruvate kinase (Pyk2p) in Saccharomyces cerevisiae that is catalytically insensitive to fructose-1,6-bisphosphate. J. Bacteriol. 1997, 179, 2987–2993. [Google Scholar] [CrossRef] [PubMed]
- Kochanowski, K.; Volkmer, B.; Gerosa, L.; van Rijsewijk, B.R.; Schmidt, A.; Heinemann, M. Functioning of a metabolic flux sensor in Escherichia coli. Proc. Natl. Acad. Sci. USA 2013, 110, 1130–1135. [Google Scholar] [CrossRef] [PubMed]
- Portela, P.; Howell, S.; Moreno, S.; Rossi, S. In vivo and in vitro phosphorylation of two Isoforms of yeast pyruvate kinase by protein kinase A. J. Biol. Chem. 2002, 277, 30477–30487. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.P.; Ludwig, C.; Picotti, P.; Kogadeeva, M.; Aebersold, R.; Sauer, U. Regulation of yeast central metabolism by enzyme phosphorylation. Mol. Syst. Biol. 2012, 8, 623. [Google Scholar] [CrossRef]
- Soulard, A.; Cremonesi, A.; Moes, S.; Schutz, F.; Jeno, P.; Hall, M.N. The rapamycin-sensitive phosphoproteome reveals that TOR controls protein kinase A toward some but not all substrates. Mol. Biol. Cell 2010, 21, 3475–3486. [Google Scholar] [CrossRef]
- Xu, Y.F.; Zhao, X.; Glass, D.S.; Absalan, F.; Perlman, D.H.; Broach, J.R.; Rabinowitz, J.D. Regulation of yeast pyruvate kinase by ultrasensitive allostery independent of phosphorylation. Mol. Cell 2012, 48, 52–62. [Google Scholar] [CrossRef]
- Saad, S.; Cereghetti, G.; Feng, Y.; Picotti, P.; Peter, M.; Dechant, R. Reversible protein aggregation is a protective mechanism to ensure cell cycle restart after stress. Nat. Cell Biol. 2017, 19, 1202–1213. [Google Scholar] [CrossRef] [PubMed]
- Srere, P.A. Complexes of sequential metabolic enzymes. Annu. Rev. Biochem. 1987, 56, 89–124. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, J.D.; Zhao, A.; Ellington, A.D.; Marcotte, E.M. Dynamic reorganization of metabolic enzymes into intracellular bodies. Annu. Rev. Cell Dev. Biol. 2012, 28, 89–111. [Google Scholar] [CrossRef] [PubMed]
- Hyman, A.A.; Simons, K. Cell biology. Beyond oil and water—phase transitions in cells. Science 2012, 337, 1047–1049. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.Z.; Gitai, Z. Beyond the cytoskeleton: mesoscale assemblies and their function in spatial organization. Curr. Opin. Microbiol. 2013, 16, 177–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prouteau, M.; Desfosses, A.; Sieben, C.; Bourgoint, C.; Lydia Mozaffari, N.; Demurtas, D.; Mitra, A.K.; Guichard, P.; Manley, S.; Loewith, R. TORC1 organized in inhibited domains (TOROIDs) regulate TORC1 activity. Nature 2017, 550, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Narayanaswamy, R.; Levy, M.; Tsechansky, M.; Stovall, G.M.; O’Connell, J.D.; Mirrielees, J.; Ellington, A.D.; Marcotte, E.M. Widespread reorganization of metabolic enzymes into reversible assemblies upon nutrient starvation. Proc. Natl. Acad. Sci. USA 2009, 106, 10147–10152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noree, C.; Sato, B.K.; Broyer, R.M.; Wilhelm, J.E. Identification of novel filament-forming proteins in Saccharomyces cerevisiae and Drosophila melanogaster. J. Cell Biol. 2010, 190, 541–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Q.J.; Kassim, H.; Huang, Y.; Li, H.; Zhang, J.; Li, G.; Wang, P.Y.; Yan, J.; Ye, F.; Liu, J.L. filamentation of metabolic enzymes in Saccharomyces cerevisiae. J. Genet. Genom./Yi Chuan Xue Bao 2016, 43, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Foe, L.G.; Trujillo, J.L. Quaternary structure of pig liver phosphofructokinase. J. Biol. Chem. 1980, 255, 10537–10541. [Google Scholar]
- Webb, B.A.; Dosey, A.M.; Wittmann, T.; Kollman, J.M.; Barber, D.L. The glycolytic enzyme phosphofructokinase-1 assembles into filaments. J. Cell Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Fuller, G.G.; Han, T.; Yao, Y.; Alessi, A.F.; Freeberg, M.A.; Roach, N.P.; Moresco, J.J.; Karnovsky, A.; Baba, M.; et al. Glycolytic enzymes coalesce in G bodies under hypoxic stress. Cell Rep. 2017, 20, 895–908. [Google Scholar] [CrossRef] [PubMed]
- Ingerson-Mahar, M.; Briegel, A.; Werner, J.N.; Jensen, G.J.; Gitai, Z. The metabolic enzyme CTP synthase forms cytoskeletal filaments. Nat. Cell Biol. 2010, 12, 739–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.L. Intracellular compartmentation of CTP synthase in Drosophila. J. Genet. Genom./Yi Chuan Xue Bao 2010, 37, 281–296. [Google Scholar] [CrossRef]
- Lowe, N.; Rees, J.S.; Roote, J.; Ryder, E.; Armean, I.M.; Johnson, G.; Drummond, E.; Spriggs, H.; Drummond, J.; Magbanua, J.P.; et al. Analysis of the expression patterns, subcellular localisations and interaction partners of Drosophila proteins using a pigP protein trap library. Development 2014, 141, 3994–4005. [Google Scholar] [CrossRef] [Green Version]
- Habrian, C.; Chandrasekhara, A.; Shahrvini, B.; Hua, B.; Lee, J.; Jesinghaus, R.; Barry, R.; Gitai, Z.; Kollman, J.; Baldwin, E.P. Inhibition of Escherichia coli CTP Synthetase by NADH and Other Nicotinamides and Their Mutual Interactions with CTP and GTP. Biochemistry 2016, 55, 5554–5565. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Gam, J.; French, J.B.; Zhao, H.; An, S.; Benkovic, S.J. Mapping protein-protein proximity in the purinosome. J. Biol. Chem. 2012, 287, 36201–36207. [Google Scholar] [CrossRef]
- An, S.; Kumar, R.; Sheets, E.D.; Benkovic, S.J. Reversible Compartmentalization of de Novo Purine Biosynthetic Complexes in Living Cells. Science 2008, 320, 103–106. [Google Scholar] [CrossRef]
- Gregolin, C.; Ryder, E.; Kleinschmidt, A.K.; Warner, R.C.; Lane, M.D. Molecular characteristics of liver acetyl CoA carboxylase. Proc. Natl. Acad. Sci. USA 1966, 56, 148–155. [Google Scholar] [CrossRef]
- Gregolin, C.; Ryder, E.; Warner, R.C.; Kleinschmidt, A.K.; Lane, M.D. Liver acetyl CoA carboxylase: The dissociation-reassociation process and its relation to catalytic activity. Proc. Natl. Acad. Sci. USA 1966, 56, 1751–1758. [Google Scholar] [CrossRef]
- Suresh, H.G.; da Silveira Dos Santos, A.X.; Kukulski, W.; Tyedmers, J.; Riezman, H.; Bukau, B.; Mogk, A. Prolonged starvation drives reversible sequestration of lipid biosynthetic enzymes and organelle reorganization in Saccharomyces cerevisiae. Mol. Biol. Cell 2015, 26, 1601–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meredith, M.J.; Lane, M.D. Acetyl-CoA carboxylase. Evidence for polymeric filament to protomer transition in the intact avian liver cell. J. Biol. Chem. 1978, 253, 3381–3383. [Google Scholar]
- Hunkeler, M.; Hagmann, A.; Stuttfeld, E.; Chami, M.; Guri, Y.; Stahlberg, H.; Maier, T. Structural basis for regulation of human acetyl-CoA carboxylase. Nature 2018, 558, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.W.; Moon, Y.A.; Park, S.W.; Cheng, D.; Kwon, H.J.; Horton, J.D. Induced polymerization of mammalian acetyl-CoA carboxylase by MIG12 provides a tertiary level of regulation of fatty acid synthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 1926–1931. [Google Scholar] [CrossRef] [PubMed]
- Beaty, N.B.; Lane, M.D. The polymerization of acetyl-CoA carboxylase. J. Biol. Chem. 1983, 258, 13051–13055. [Google Scholar] [PubMed]
- Eisenberg, H.; Tomkins, G.M. Molecular weight of the subunits, oligomeric and associated forms of bovine liver glutamate dehydrogenase. J. Mol. Biol. 1968, 31, 37–49. [Google Scholar] [CrossRef]
- Cohen, R.J.; Benedek, G.B. The functional relationship between the polymerization and catalytic activity of beef liver glutamate dehydrogenase. III. Analysis of Thusius’ critique. J. Mol. Biol. 1979, 129, 37–44. [Google Scholar] [CrossRef]
- Petrovska, I.; Nuske, E.; Munder, M.C.; Kulasegaran, G.; Malinovska, L.; Kroschwald, S.; Richter, D.; Fahmy, K.; Gibson, K.; Verbavatz, J.M.; et al. Filament formation by metabolic enzymes is a specific adaptation to an advanced state of cellular starvation. eLife 2014. [Google Scholar] [CrossRef]
- Josephs, R.; Borisy, G. Self-assembly of glutamic dehydrogenase into ordered superstructures: Multichain tubes formed by association of single molecules. J. Mol. Biol. 1972, 65, 127–155. [Google Scholar] [CrossRef]
- Frey, T.G.; Eisenberg, D.; Eiserling, F.A. Glutamine synthetase forms three- and seven-stranded helical cables. Proc. Natl. Acad. Sci. USA 1975, 72, 3402–3406. [Google Scholar] [CrossRef]
- Olsen, B.R.; Svenneby, G.; Kvamme, E.; Tveit, B.; Eskeland, T. Formation and ultrastructure of enzymically active polymers of pig renal glutaminase. J. Mol. Biol. 1970, 52, 239–245. [Google Scholar] [CrossRef]
- Alberti, S. Phase separation in biology. Curr. Biol. 2017, 27, R1097–R1102. [Google Scholar] [CrossRef] [PubMed]
- Brangwynne, C.P.; Tompa, P.; Pappu, R.V. Polymer physics of intracellular phase transitions. Nat. Phys. 2015, 11, 899–904. [Google Scholar] [CrossRef]
- Franzmann, T.M.; Jahnel, M.; Pozniakovsky, A.; Mahamid, J.; Holehouse, A.S.; Nuske, E.; Richter, D.; Baumeister, W.; Grill, S.W.; Pappu, R.V.; et al. Phase separation of a yeast prion protein promotes cellular fitness. Science 2018, 359. [Google Scholar] [CrossRef] [PubMed]
- Aguzzi, A.; Lakkaraju, A.K. Cell Biology of Prions and Prionoids: A Status Report. Trends Cell Biol. 2016, 26, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Lee, H.O.; Jawerth, L.; Maharana, S.; Jahnel, M.; Hein, M.Y.; Stoynov, S.; Mahamid, J.; Saha, S.; Franzmann, T.M.; et al. A Liquid-to-Solid Phase Transition of the ALS Protein FUS Accelerated by Disease Mutation. Cell 2015, 162, 1066–1077. [Google Scholar] [CrossRef] [PubMed]
- Peskett, T.R.; Rau, F.; O’Driscoll, J.; Patani, R.; Lowe, A.R.; Saibil, H.R. A Liquid to Solid Phase Transition Underlying Pathological Huntingtin Exon1 Aggregation. Mol. Cell 2018, 70, 588–601. [Google Scholar] [CrossRef]
- Kleinschmidt, A.K.; Moss, J.; Lane, D.M. Acetyl coenzyme A carboxylase: filamentous nature of the animal enzymes. Science 1969, 166, 1276–1278. [Google Scholar] [CrossRef]
- Trujillo, J.L.; Deal, W.C., Jr. Pig liver phosphofructokinase: asymmetry properties, proof of rapid association—dissociation equilibria, and effect of temperature and protein concentration on the equilibria. Biochemistry 1977, 16, 3098–3104. [Google Scholar] [CrossRef]
- Barry, R.M.; Bitbol, A.F.; Lorestani, A.; Charles, E.J.; Habrian, C.H.; Hansen, J.M.; Li, H.J.; Baldwin, E.P.; Wingreen, N.S.; Kollman, J.M.; et al. Large-scale filament formation inhibits the activity of CTP synthetase. eLife 2014, 3, e03638. [Google Scholar] [CrossRef] [Green Version]
- Lynch, E.M.; Hicks, D.R.; Shepherd, M.; Endrizzi, J.A.; Maker, A.; Hansen, J.M.; Barry, R.M.; Gitai, Z.; Baldwin, E.P.; Kollman, J.M. Human CTP synthase filament structure reveals the active enzyme conformation. Nat. Struct. Mol. Biol. 2017, 24, 507–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azzam, G.; Liu, J.L. Only one isoform of Drosophila melanogaster CTP synthase forms the cytoophidium. PLoS Genet. 2013, 9, e1003256. [Google Scholar] [CrossRef] [PubMed]
- He, Y.X.; Gui, L.; Liu, Y.Z.; Du, Y.; Zhou, Y.; Li, P.; Zhou, C.Z. Crystal structure of Saccharomyces cerevisiae glutamine synthetase Gln1 suggests a nanotube-like supramolecular assembly. Proteins 2009, 76, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Ereno-Orbea, J.; Majtan, T.; Oyenarte, I.; Kraus, J.P.; Martinez-Cruz, L.A. Structural basis of regulation and oligomerization of human cystathionine β-synthase, the central enzyme of transsulfuration. Proc. Natl. Acad. Sci. USA 2013, 110, E3790–3799. [Google Scholar] [CrossRef] [PubMed]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR signaling in growth and metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Wippich, F.; Bodenmiller, B.; Trajkovska, M.G.; Wanka, S.; Aebersold, R.; Pelkmans, L. Dual specificity kinase DYRK3 couples stress granule condensation/dissolution to mTORC1 signaling. Cell 2013, 152, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Thedieck, K.; Holzwarth, B.; Prentzell, M.T.; Boehlke, C.; Klasener, K.; Ruf, S.; Sonntag, A.G.; Maerz, L.; Grellscheid, S.N.; Kremmer, E.; et al. Inhibition of mTORC1 by astrin and stress granules prevents apoptosis in cancer cells. Cell 2013, 154, 859–874. [Google Scholar] [CrossRef] [PubMed]
- Takahara, T.; Maeda, T. Transient sequestration of TORC1 into stress granules during heat stress. Mol. Cell 2012, 47, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Hughes Hallett, J.E.; Luo, X.; Capaldi, A.P. Snf1/AMPK promotes the formation of Kog1/Raptor-bodies to increase the activation threshold of TORC1 in budding yeast. eLife 2015, 4, e09181. [Google Scholar] [CrossRef] [PubMed]
- Kira, S.; Kumano, Y.; Ukai, H.; Takeda, E.; Matsuura, A.; Noda, T. Dynamic relocation of the TORC1-Gtr1/2-Ego1/2/3 complex is regulated by Gtr1 and Gtr2. Mol. Biol. Cell 2016, 27, 382–396. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.A.; Forouhar, F.; Szu, F.E.; Seetharaman, J.; Tong, L.; Barber, D.L. Structures of human phosphofructokinase-1 and atomic basis of cancer-associated mutations. Nature 2015, 523, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aylett, C.H.; Sauer, E.; Imseng, S.; Boehringer, D.; Hall, M.N.; Ban, N.; Maier, T. Architecture of human mTOR complex 1. Science 2016, 351, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Seisdedos, H.; Empereur-Mot, C.; Elad, N.; Levy, E.D. Proteins evolve on the edge of supramolecular self-assembly. Nature 2017, 548, 244–247. [Google Scholar] [PubMed]
- Pedley, A.M.; Benkovic, S.J. A New View into the Regulation of Purine Metabolism: The Purinosome. Trends Biochem. Sci. 2017, 42, 141–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Metabolic Functions | Enzyme Name | Protomer | Condensates | Yeast | References | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| B | Y | D | M | Gene Name | Stress Triggering Condensation | |||||
| Carbohydrate metabolism | Glycogen debranching enzyme | ☑ | Gdb1 | SP | [19] | |||||
| ATP-dependent 6-phosphofructokinase | Tetramer | ☑ | ☑ | Pfk1/2 | SP | [19,20,21] | ||||
| Pyruvate kinase | Tetramer | ☑ | ☑ | Cdc19 | SP, GS, HS | [11,17] | ||||
| Enolase | Dimer | ☑ | Eno2 | HY | [22] | |||||
| Fructose bisphosphate aldolase | ☑ | Fba1 | HY | [22] | ||||||
| Alcool deshydrogenase | Tetramer | ☑ | Adh2 | SP | [17] | |||||
| UTP-glucose-1-phosphate uridylyltransferase | Dimer to octamer | ☑ | Ugp1 | SP | [17] | |||||
| Nucleotide metabolism | Pyrimidine synthesis | CTP synthase | Dimer and tetramer | ☑ | ☑ | ☑ | ☑ | Ura7/8 | SP, GS | [18,23,24,25,26] |
| Trifunctional CAD enzyme | - | ☑ | Ura2 | SP | [17] | |||||
| Adenylosuccinate synthetase | Dimer | ☑ | Ade12 | SP | [17] | |||||
| De novo purine synthesis | PPAT enzyme | Tetramer | ☑ | ☑ | Ade4 | SP | [17,27] | |||
| Trifunctional GART enzyme | Dimer | ☑ | ☑ | ☑ | Ade5/7 | SP, PS | [17,25,27,28] | |||
| Bifunctional PAICS enzyme | Octamer | ☑ | ☑ | ☑ | Ade2 | - | [25,27] | |||
| FGAMS enzyme | Monomer | ☑ | ☑ | Ade6 | - | [17,27,28] | ||||
| ADSL enzyme | Tetramer | ☑ | Ade13 | - | [27,28] | |||||
| Bifunctional ATIC enzyme | Dimer | ☑ | ☑ | Ade17 | SP, PS | [17,27,28] | ||||
| Fatty acids and Sterol metabolism | Acetyl-CoA carboxylase | Dimer and tetramer | ☑ | ☑ | Acc1/2 | SP, GS | [19,29,30,31,32,33,34,35] | |||
| Fatty acid synthase complex | - | ☑ | Fas1/2 | SP, GS | [17,31] | |||||
| Sterol 3-beta-glucosyltransferase | - | ☑ | Ugt51 | SP | [17] | |||||
| Amino acids metabolism | Asparagine synthetase | Dimer | ☑ | Asn1/2 | SP | [19] | ||||
| Glutamate synthetase | Hexamer | ☑ | Glt1 | SP | [18,19,36,37,38] | |||||
| Glutamate dehydrogenase | Hexamer | ☑ | ☑ | Gdh2 | SP | [19,39] | ||||
| Glutamine synthetase | Decamer | ☑ | ☑ | Gln1 | GS + apH | [17,19,38,40] | ||||
| Glutaminase | Tetramer | ☑ | - | - | [41] | |||||
| Cystathionine beta-synthase | Tetramer | ☑ | Cys4 | SP | [17] | |||||
| Metabolism regulator | Target of Rapamycin Complex 1 | Dimer of heterotetramer | ☑ | TORC1 | SP, GS | [16] | ||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prouteau, M.; Loewith, R. Regulation of Cellular Metabolism through Phase Separation of Enzymes. Biomolecules 2018, 8, 160. https://doi.org/10.3390/biom8040160
Prouteau M, Loewith R. Regulation of Cellular Metabolism through Phase Separation of Enzymes. Biomolecules. 2018; 8(4):160. https://doi.org/10.3390/biom8040160
Chicago/Turabian StyleProuteau, Manoël, and Robbie Loewith. 2018. "Regulation of Cellular Metabolism through Phase Separation of Enzymes" Biomolecules 8, no. 4: 160. https://doi.org/10.3390/biom8040160
APA StyleProuteau, M., & Loewith, R. (2018). Regulation of Cellular Metabolism through Phase Separation of Enzymes. Biomolecules, 8(4), 160. https://doi.org/10.3390/biom8040160