Effect of Coenzyme Q10 Supplementation on Testosterone


Abstract
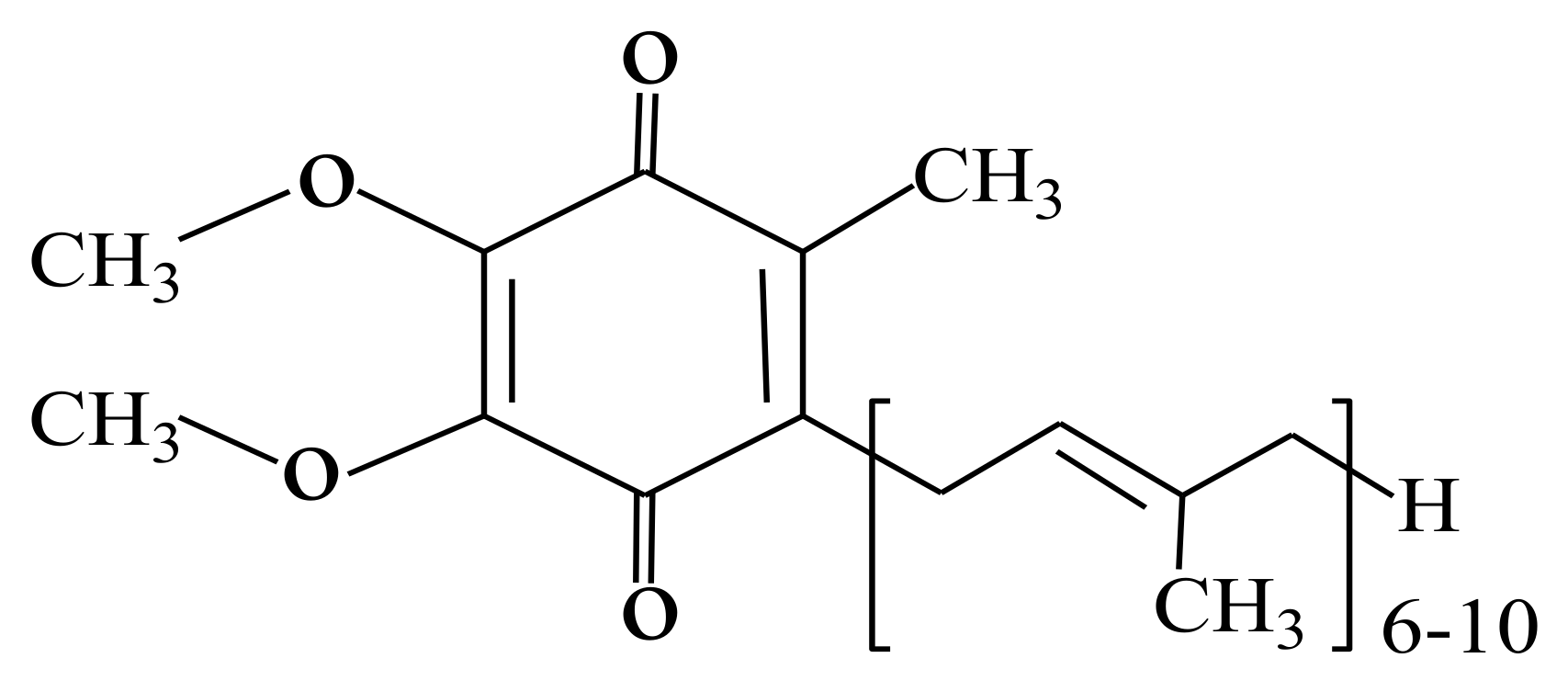
:1. Introduction
2. Effect of Coenzyme Q10 Supplementation on Testosterone
3. Mechanistic Studies
4. Conclusions and Future Perspectives
Funding
Acknowledgments
Conflicts of Interest
References
- Sood, B.; Keenaghan, M. Coenzyme Q10; StatPearls Publishing: Treasure Island, FL, USA, 2018. [Google Scholar]
- Battino, M.; Ferri, E.; Gorini, A.; Villa, R.F.; Rodriguez Huertas, J.F.; Fiorella, P.; Genova, M.L.; Lenaz, G.; Marchetti, M. Natural distribution and occurrence of coenzyme Q homologues. Membr. Biochem. 1990, 9, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Cluis, C.P.; Burja, A.M.; Martin, V.J. Current prospects for the production of coenzyme Q10 in microbes. Trends Biotechnol. 2007, 25, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Varela-Lopez, A.; Giampieri, F.; Battino, M.; Quiles, J.L. Coenzyme Q and its role in the dietary therapy against aging. Molecules 2016, 21, 373. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.; Loiselle, D.; Power, A.; Hickey, A.J. Mitochondrial inefficiencies and anoxic ATP hydrolysis capacities in diabetic rat heart. Am. J. Physiol. Cell Physiol. 2014, 307, C499–C507. [Google Scholar] [CrossRef] [PubMed]
- Merlo Pich, M.; Castagnoli, A.; Biondi, A.; Bernacchia, A.; Tazzari, P.L.; D’Aurelio, M.; Parenti Castelli, G.; Formiggini, G.; Conte, R.; Bovina, C.; et al. Ubiquinol and a coenzyme Q reducing system protect platelet mitochondrial function of transfusional buffy coats from oxidative stress. Free Radic. Res. 2002, 36, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Pepe, S.; Marasco, S.F.; Haas, S.J.; Sheeran, F.L.; Krum, H.; Rosenfeldt, F.L. Coenzyme Q10 in cardiovascular disease. Mitochondrion 2007, 7, S154–S167. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Fonarow, G.C.; Butler, J.; Ezekowitz, J.A.; Felker, G.M. Coenzyme Q10 and heart failure: A state-of-the-art review. Circ. Heart Fail. 2016, 9, e002639. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Yang, K.L.; Zeng, L.T.; Wu, X.H.; Huang, H.Y. Effectiveness of coenzyme Q10 supplementation for type 2 diabetes mellitus: A systematic review and meta-analysis. Int. J. Endocrinol. 2018, 2018, 6484839. [Google Scholar] [CrossRef]
- Tarry-Adkins, J.L.; Fernandez-Twinn, D.S.; Madsen, R.; Chen, J.H.; Carpenter, A.; Hargreaves, I.P.; McConnell, J.M.; Ozanne, S.E. Coenzyme Q10 prevents insulin signaling dysregulation and inflammation prior to development of insulin resistance in male offspring of a rat model of poor maternal nutrition and accelerated postnatal growth. Endocrinology 2015, 156, 3528–3537. [Google Scholar] [CrossRef]
- Hernandez-Camacho, J.D.; Bernier, M.; Lopez-Lluch, G.; Navas, P. Coenzyme Q10 supplementation in aging and disease. Front. Physiol. 2018, 9, 44. [Google Scholar] [CrossRef]
- Mischley, L.K.; Lau, R.C.; Bennett, R.D. Role of diet and nutritional supplements in Parkinson’s disease progression. Oxid. Med. Cell. Longev. 2017, 2017, 6405278. [Google Scholar] [CrossRef] [PubMed]
- Yoritaka, A.; Kawajiri, S.; Yamamoto, Y.; Nakahara, T.; Ando, M.; Hashimoto, K.; Nagase, M.; Saito, Y.; Hattori, N. Randomized, double-blind, placebo-controlled pilot trial of reduced coenzyme Q10 for Parkinson’s disease. Parkinsonism Relat. Disord. 2015, 21, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Shannon, K.M.; Fraint, A. Therapeutic advances in Huntington’s disease. Mov. Disord. 2015, 30, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.T.; Huang, Y.C.; Cheng, S.B.; Huang, Y.T.; Lin, P.T. Effects of coenzyme Q10 supplementation on antioxidant capacity and inflammation in hepatocellular carcinoma patients after surgery: A randomized, placebo-controlled trial. Nutr. J. 2016, 15, 85. [Google Scholar] [CrossRef]
- Liu, H.T.; Cheng, S.B.; Huang, Y.C.; Huang, Y.T.; Lin, P.T. Coenzyme Q10 and oxidative stress: Inflammation status in hepatocellular carcinoma patients after surgery. Nutrients 2017, 9, 29. [Google Scholar] [CrossRef]
- Jain, S.; Garg, T.; Kushwah, V.; Thanki, K.; Agrawal, A.K.; Dora, C.P. α-Tocopherol as functional excipient for resveratrol and coenzyme Q10-loaded SNEDDS for improved bioavailability and prophylaxis of breast cancer. J. Drug Target. 2017, 25, 554–565. [Google Scholar] [CrossRef]
- Gvozdjakova, A.; Kucharska, J.; Dubravicky, J.; Mojto, V.; Singh, R.B. Coenzyme Q10, α-tocopherol, and oxidative stress could be important metabolic biomarkers of male infertility. Dis. Markers 2015, 2015, 827941. [Google Scholar] [CrossRef]
- Hathcock, J.N.; Shao, A. Risk assessment for coenzyme Q10 (ubiquinone). Regul. Toxicol. Pharmacol. 2006, 45, 282–288. [Google Scholar] [CrossRef]
- McGarry, A.; McDermott, M.; Kieburtz, K.; de Blieck, E.A.; Beal, F.; Marder, K.; Ross, C.; Shoulson, I.; Gilbert, P.; Mallonee, W.M.; et al. A randomized, double-blind, placebo-controlled trial of coenzyme Q10 in Huntington disease. Neurology 2017, 88, 152–159. [Google Scholar] [CrossRef]
- Banihani, S.A. Ginger and testosterone. Biomolecules 2018, 8, 119. [Google Scholar] [CrossRef]
- Mooradian, A.D.; Morley, J.E.; Korenman, S.G. Biological actions of androgens. Endocr. Rev. 1987, 8, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Khaki, A.; Khaki, A.A.; Hajhosseini, L.; Golzar, F.S.; Ainehchi, N. The anti-oxidant effects of ginger and cinnamon on spermatogenesis dys-function of diabetes rats. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Paternostro, G. Longevity and testosterone. Nature 1994, 368, 408. [Google Scholar] [CrossRef] [PubMed]
- Audet-Walsh, E.; Yee, T.; Tam, I.S.; Giguere, V. Inverse regulation of DHT synthesis enzymes 5α-reductase types 1 and 2 by the androgen receptor in prostate cancer. Endocrinology 2017, 158, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Dehghan, F.; Muniandy, S.; Yusof, A.; Salleh, N. Testosterone reduces knee passive range of motion and expression of relaxin receptor isoforms via 5α-dihydrotestosterone and androgen receptor binding. Int. J. Mol. Sci. 2014, 15, 4619–4634. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.M.; Akhtar, S.; Sellers, D.J.; Muraleedharan, V.; Channer, K.S.; Jones, T.H. Testosterone differentially regulates targets of lipid and glucose metabolism in liver, muscle and adipose tissues of the testicular feminised mouse. Endocrine 2016, 54, 504–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermeulen, A.; Kaufman, J.M.; Goemaere, S.; van Pottelberg, I. Estradiol in elderly men. Aging Male 2002, 5, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Travison, T.G.; Morley, J.E.; Araujo, A.B.; O’Donnell, A.B.; McKinlay, J.B. The relationship between libido and testosterone levels in aging men. J. Clin. Endocrinol. Metab. 2006, 91, 2509–2513. [Google Scholar] [CrossRef]
- Moffat, S.D. Effects of testosterone on cognitive and brain aging in elderly men. Ann. N. Y. Acad. Sci. 2005, 1055, 80–92. [Google Scholar] [CrossRef]
- Bassil, N.; Alkaade, S.; Morley, J.E. The benefits and risks of testosterone replacement therapy: A review. Ther. Clin. Risk Manag. 2009, 5, 427–448. [Google Scholar]
- Karakas, M.; Schafer, S.; Appelbaum, S.; Ojeda, F.; Kuulasmaa, K.; Bruckmann, B.; Berisha, F.; Schulte-Steinberg, B.; Jousilahti, P.; Blankenberg, S.; et al. Testosterone levels and type 2 diabetes—No correlation with age, differential predictive value in men and women. Biomolecules 2018, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Tuck, S.P.; Francis, R.M. Testosterone, bone and osteoporosis. Front. Horm Res. 2009, 37, 123–132. [Google Scholar] [PubMed]
- Yarrow, J.F.; Phillips, E.G.; Conover, C.F.; Bassett, T.E.; Chen, C.; Teurlings, T.; Vasconez, A.; Alerte, J.; Prock, H.; Jiron, J.M.; et al. Testosterone plus finasteride prevents bone loss without prostate growth in a rodent spinal cord injury model. J. Neurotrauma 2017, 34, 2972–2981. [Google Scholar] [CrossRef] [PubMed]
- El-Migdadi, F.; Banihani, I.; Banihani, S.A. Clinico-hormonal correlation of oligospermic patients in the below sea level environment (Jordan Valley). Neuro Endocrinol. Lett. 2005, 26, 13–18. [Google Scholar] [PubMed]
- Kelly, D.M.; Jones, T.H. Testosterone: A metabolic hormone in health and disease. J. Endocrinol. 2013, 217, R25–R45. [Google Scholar] [CrossRef] [PubMed]
- Hoenjet, K.M.; Dagnelie, P.C.; Delaere, K.P.; Wijckmans, N.E.; Zambon, J.V.; Oosterhof, G.O. Effect of a nutritional supplement containing vitamin E, selenium, vitamin C and coenzyme Q10 on serum PSA in patients with hormonally untreated carcinoma of the prostate: A randomised placebo-controlled study. Eur. Urol. 2005, 47, 433–439, discussion 439–440. [Google Scholar] [CrossRef]
- Safarinejad, M.R. Efficacy of coenzyme Q10 on semen parameters, sperm function and reproductive hormones in infertile men. J. Urol. 2009, 182, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Safarinejad, M.R. The effect of coenzyme Q10 supplementation on partner pregnancy rate in infertile men with idiopathic oligoasthenoteratozoospermia: An open-label prospective study. Int. Urol. Nephrol. 2012, 44, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.F.; Xing, Y.; Wu, C.Y.; Liu, R.Z.; Wang, X.Y.; Xing, J.P. Tamoxifen combined with coenzyme Q10 for idiopathic oligoasthenospermia. Zhonghua Nan Ke Xue 2011, 17, 615–618. (In Chinese) [Google Scholar]
- Izadi, A.; Ebrahimi, S.; Shirzai, S.; Taghizadeh, S.; Parizad, M.; Farzadi, L.; Pourghassem Gargari, B. Hormonal and metabolic effects of coenzyme Q10 and/or vitamin E in patients with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2018. [Google Scholar] [CrossRef]
- Souri, Z.; Bidmeshki-Pour, A.; Karimi, I. Effect of coenzyme Q10 supplementation on p53 tumor suppressor gene expression in mouse model of andropause. Biharean Biol. 2017, 11, 53–56. [Google Scholar]
- Souri, Z.; Bidmeshki-Pour, A.; Karimi, I. Influence of coenzyme Q10 supplementation on skin follicle characteristics in gonadectomized mice. Biharean Biol. 2016, 10, 12–15. [Google Scholar]
- Vazir, N.; Adibmoradi, M.; Tajik, P.; Rezaian, M.; Babapour, V.; Baghcheghi, Y. A morphometrical study of testis and histometrical study of seminiferous tubules in coenzyme Q10 fed ostriches. J. Vet. Res. 2013, 68, 349–357. [Google Scholar]
- Fouad, A.A.; Al-Sultan, A.I.; Yacoubi, M.T. Coenzyme Q10 counteracts testicular injury induced by sodium arsenite in rats. Eur. J. Pharmacol. 2011, 655, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Ghanbarzadeh, S.; Garjani, A.; Ziaee, M.; Khorrami, A. Effects of l-carnitine and coenzyme Q10 on impaired spermatogenesis caused by isoproterenol in male rats. Drug Res. (Stuttg.) 2014, 64, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, N.S.; Arafa, M.H.; Atteia, H.H. Coenzyme Q10 and fish oil synergistically alleviate aluminum chloride-induced suppression of testicular steroidogenesis and antioxidant defense. Free Radic. Res. 2015, 49, 1319–1334. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.; Fournier, M.A.; Maltais, R.; Kenmogne, L.C.; Poirier, D. In vitro and in vivo evaluation of a 3β-androsterone derivative as inhibitor of 17β-hydroxysteroid dehydrogenase type 3. J. Steroid Biochem. Mol. Biol. 2014, 141, 44–51. [Google Scholar] [CrossRef]
- Griswold, M.D. The central role of Sertoli cells in spermatogenesis. Semin. Cell Dev. Biol. 1998, 9, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Lafuente, R.; Gonzalez-Comadran, M.; Sola, I.; Lopez, G.; Brassesco, M.; Carreras, R.; Checa, M.A. Coenzyme Q10 and male infertility: A meta-analysis. J. Assist. Reprod. Genet. 2013, 30, 1147–1156. [Google Scholar] [CrossRef]
- Al-Amin, M.M.; Reza, H.M.; Saadi, H.M.; Mahmud, W.; Ibrahim, A.A.; Alam, M.M.; Kabir, N.; Saifullah, A.R.; Tropa, S.T.; Quddus, A.H. Astaxanthin ameliorates aluminum chloride-induced spatial memory impairment and neuronal oxidative stress in mice. Eur. J. Pharmacol. 2016, 777, 60–69. [Google Scholar] [CrossRef]
- Banihani, S.A. Vitamin B12 and semen quality. Biomolecules 2017, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kshetrimayum, C.; Sadhu, H.G.; Kumar, S. Arsenic-induced oxidative stress, cholinesterase activity in the brain of swiss albino mice, and its amelioration by antioxidants vitamin E and coenzyme Q10. Environ. Sci. Pollut. Res. Int. 2018, 25, 23946–23953. [Google Scholar] [CrossRef] [PubMed]
- Bergquist, E.R.; Fischer, R.J.; Sugden, K.D.; Martin, B.D. Inhibition by methylated organo-arsenicals of the respiratory 2-oxo-acid dehydrogenases. J. Organomet. Chem. 2009, 694, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Pant, N.; Murthy, R.C.; Srivastava, S.P. Male reproductive toxicity of sodium arsenite in mice. Hum. Exp. Toxicol. 2004, 23, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Banihani, S.A. Role of uric acid in semen. Biomolecules 2018, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Chiou, T.J.; Chu, S.T.; Tzeng, W.F.; Huang, Y.C.; Liao, C.J. Arsenic trioxide impairs spermatogenesis via reducing gene expression levels in testosterone synthesis pathway. Chem. Res. Toxicol. 2008, 21, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Fetherolf, M.M.; Boyd, S.D.; Taylor, A.B.; Kim, H.J.; Wohlschlegel, J.A.; Blackburn, N.J.; Hart, P.J.; Winge, D.R.; Winkler, D.D. Copper-zinc superoxide dismutase is activated through a sulfenic acid intermediate at a copper ion entry site. J. Biol. Chem. 2017, 292, 12025–12040. [Google Scholar] [CrossRef]
- Salem, K.; McCormick, M.L.; Wendlandt, E.; Zhan, F.; Goel, A. Copper-zinc superoxide dismutase-mediated redox regulation of bortezomib resistance in multiple myeloma. Redox. Biol. 2015, 4, 23–33. [Google Scholar] [CrossRef]
- Paunovic, M.G.; Matic, M.M.; Ognjanovic, B.I.; Saicic, Z.S. Antioxidative and haematoprotective activity of coenzyme Q10 and vitamin E against cadmium-induced oxidative stress in Wistar rats. Toxicol. Ind. Health 2017, 33, 746–756. [Google Scholar] [CrossRef]
- Knott, A.; Achterberg, V.; Smuda, C.; Mielke, H.; Sperling, G.; Dunckelmann, K.; Vogelsang, A.; Kruger, A.; Schwengler, H.; Behtash, M.; et al. Topical treatment with coenzyme Q10-containing formulas improves skin’s Q10 level and provides antioxidative effects. Biofactors 2015, 41, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Hu, C.; Xue, Y. In vitro evaluation of chitosan-coated liposome containing both coenzyme Q10 and α-lipoic acid: Cytotoxicity, antioxidant activity, and antimicrobial activity. J. Cosmet. Dermatol. 2018, 17, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Du, J.; Lian, Y.; Zhang, Y.; Li, X.; Liu, Y.; Zou, L.; Wu, T. Protective effects of coenzyme Q10 against hydrogen peroxide-induced oxidative stress in PC12 cell: The role of Nrf2 and antioxidant enzymes. Cell. Mol. Neurobiol. 2016, 36, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Pala, R.; Orhan, C.; Tuzcu, M.; Sahin, N.; Ali, S.; Cinar, V.; Atalay, M.; Sahin, K. Coenzyme Q10 supplementation modulates NFκb and Nrf2 pathways in exercise training. J. Sports Sci. Med. 2016, 15, 196–203. [Google Scholar]
- Rahmani, A.; Abangah, G.; Moradkhani, A.; Hafezi Ahmadi, M.R.; Asadollahi, K. Coenzyme Q10 in combination with triple therapy regimens ameliorates oxidative stress and lipid peroxidation in chronic gastritis associated with H. pylori infection. J. Clin. Pharmacol. 2015, 55, 842–847. [Google Scholar] [CrossRef]
- Ahmadvand, H.; Ghasemi Dehnoo, M.; Dehghani, A.; Bagheri, S.; Cheraghi, R.A. Serum paraoxonase 1 status and its association with atherogenic indexes in gentamicin-induced nephrotoxicity in rats treated with coenzyme Q10. Ren. Fail. 2014, 36, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Orlando, P.; Silvestri, S.; Galeazzi, R.; Antonicelli, R.; Marcheggiani, F.; Cirilli, I.; Bacchetti, T.; Tiano, L. Effect of ubiquinol supplementation on biochemical and oxidative stress indexes after intense exercise in young athletes. Redox Rep. 2018, 23, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Palmeira, C.M.; Santos, D.L.; Seica, R.; Moreno, A.J.; Santos, M.S. Enhanced mitochondrial testicular antioxidant capacity in Goto-Kakizaki diabetic rats: Role of coenzyme Q. Am. J. Physiol. Cell Physiol. 2001, 281, C1023–C1028. [Google Scholar] [CrossRef]


{kind=link}
| Source | Dose (Mode of Treatment) | Duration | Study Population | Effect on Testosterone | Ref. |
|---|---|---|---|---|---|
| Coenzyme Q10 (Bio-Quinon Q10) | 200 mg day−1 (orally) | 21 weeks | Patients with hormonally untreated carcinoma of the prostate | (±) | [37] |
| Coenzyme Q10 (Kaneka, Osaka, Japan) | 300 mg day−1 (orally) | 26 weeks | Infertile men | (±) | [38] |
| Coenzyme Q10 (Nutri Q10, Nutri Century, Toronto, ON, Canada) | 900 mg day−1 (orally) | 12 months | Infertile men with idiopathic oligoasthenoteratozoospermia | (±) | [39] |
| Coenzyme Q10 | Therapeutic dose (orally) | 3 and 6 months | Patients with idiopathic oligoasthenospermia | (±) | [40] |
| Coenzyme Q10 (Nature Made Pharmaceutical Company, Mission Hills, CA, USA) | 200 mg day−1 (orally) | 8 weeks | Patients with polycystic ovary syndrome | (−) | [41] |
| Coenzyme Q10 (Nutralife Co., Richmond Hill, Canada) | 125, 250, and 500 mg kg−1 day−1 (orally) | 96 days | Bilateral orchidectomized male mice | (±) | [42] |
| Coenzyme Q10 (Nutralife Co.) | 125 and 250 mg kg−1 day−1 (orally) | 96 days | Gonadectomized male mice | (−) | [43] |
| Coenzyme Q10 | 10 and 20 mg kg−1 day−1 (orally) | 2 months | Male ostriches | (±) | [44] |
| Coenzyme Q10 (Sigma Chemical Company, St. Louis, MO, USA) | 10 mg kg−1 day−1 (intraperitoneally) | 5 days | Male rats with sodium arsenite-induced reproductive toxicity | (+) | [45] |
| Coenzyme Q10 | 10 mg kg−1 day−1 (orally) | 20 days | Male rats with isoproterenol-induced reproductive toxicity | (+) | [46] |
| Coenzyme Q10 (Arab Co. for Pharmaceuticals & Medicinal Plants, Cairo, Egypt) | 10 mg kg−1 day−1 (orally) | 10 weeks | Male rats with aluminum chloride-induced reproductive toxicity | (+) | [47] |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banihani, S.A. Effect of Coenzyme Q10 Supplementation on Testosterone. Biomolecules 2018, 8, 172. https://doi.org/10.3390/biom8040172
Banihani SA. Effect of Coenzyme Q10 Supplementation on Testosterone. Biomolecules. 2018; 8(4):172. https://doi.org/10.3390/biom8040172
Chicago/Turabian StyleBanihani, Saleem Ali. 2018. "Effect of Coenzyme Q10 Supplementation on Testosterone" Biomolecules 8, no. 4: 172. https://doi.org/10.3390/biom8040172
APA StyleBanihani, S. A. (2018). Effect of Coenzyme Q10 Supplementation on Testosterone. Biomolecules, 8(4), 172. https://doi.org/10.3390/biom8040172