Network Contractility during Cytokinesis—From Molecular to Global Views

 ,
,
Abstract
:1. Cytokinesis Overview
2. Actin and Myosin—Main Contractile Ring Components
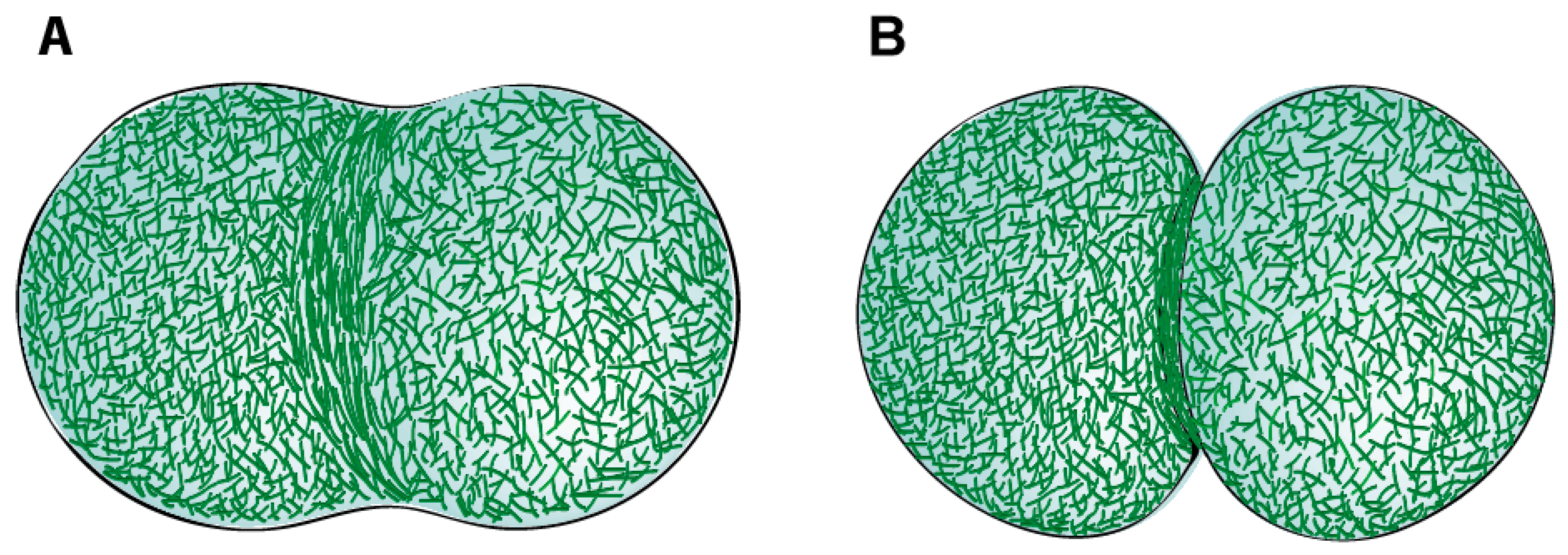
2.1. Actin Filaments
2.2. Myosin Filaments
3. Actomyosin Contractility in the Contractile Ring
3.1. Actomyosin Structure of the Contractile Ring
3.2. Ring Constriction Driven by Myosin Motor Activity
3.3. Ring Constriction Independently of Myosin Motor Activity
3.4. Importance of F-Actin Dynamics for Ring Constriction
3.5. Involvement of Actin Filament Crosslinkers in Cytokinesis
3.6. Modulation of Actomyosin Contractility by Actin Filament Crosslinkers
4. Theoretical Modeling of Cytokinesis
4.1. Modeling of Actomyosin Networks
4.2. Modeling Cytokinesis
4.3. Models of Contractile Ring Constriction
4.4. Open Questions and Future Directions on Cytokinesis Modeling
4.4.1. The Influence of the Surrounding Cortex
4.4.2. The Importance of Estimating Mechanical and Molecular Parameters in the Same System
5. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Whitman, C.O. A contribution to the history of the germ-layers in Clepsine. J. Morphol. 1887, 1, 105–182. [Google Scholar] [CrossRef]
- Lens, S.M.A.; Medema, R.H. Cytokinesis defects and cancer. Nat. Rev. Cancer 2019, 19, 32–45. [Google Scholar] [CrossRef]
- Mierzwa, B.; Gerlich, D.W. Cytokinetic abscission: Molecular mechanisms and temporal control. Dev. Cell 2014, 31, 525–538. [Google Scholar] [CrossRef]
- Rappaport, R. Repeated furrow formation from a single mitotic apparatus in cylindrical sand dollar eggs. J. Exp. Zool. 1985, 234, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Bement, W.M.; Benink, H.A.; von Dassow, G. A microtubule-dependent zone of active RhoA during cleavage plane specification. J. Cell Biol. 2005, 170, 91–101. [Google Scholar] [CrossRef]
- Wagner, E.; Glotzer, M. Local RhoA activation induces cytokinetic furrows independent of spindle position and cell cycle stage. J. Cell Biol. 2016, 213, 641–649. [Google Scholar] [CrossRef]
- Wittinghofer, A.; Vetter, I.R. Structure-function relationships of the G domain, a canonical switch motif. Annu. Rev. Biochem. 2011, 80, 943–971. [Google Scholar] [CrossRef]
- Chircop, M. Rho GTPases as regulators of mitosis and cytokinesis in mammalian cells. Small GTPases 2014, 5, e29770. [Google Scholar] [CrossRef] [PubMed]
- Jordan, S.N.; Canman, J.C. Rho GTPases in animal cell cytokinesis: An occupation by the one percent. Cytoskeleton 2012, 69, 919–930. [Google Scholar] [CrossRef]
- Kühn, S.; Geyer, M. Formins as effector proteins of Rho GTPases. Small GTPases 2014, 5, e29513. [Google Scholar] [CrossRef]
- Matsumura, F. Regulation of myosin II during cytokinesis in higher eukaryotes. Trends Cell Biol. 2005, 15, 371–377. [Google Scholar] [CrossRef]
- Eggert, U.S.; Mitchison, T.J.; Field, C.M. Animal Cytokinesis: From Parts List to Mechanisms. Annu. Rev. Biochem. 2006, 75, 543–566. [Google Scholar] [CrossRef]
- Wu, J.-Q.; Pollard, T.D. Counting cytokinesis proteins globally and locally in fission yeast. Science 2005, 310, 310–314. [Google Scholar] [CrossRef]
- Courtemanche, N.; Pollard, T.D.; Chen, Q. Avoiding artefacts when counting polymerized actin in live cells with LifeAct fused to fluorescent proteins. Nat. Cell Biol. 2016, 18, 676–683. [Google Scholar] [CrossRef]
- Laplante, C.; Huang, F.; Tebbs, I.R.; Bewersdorf, J.; Pollard, T.D. Molecular organization of cytokinesis nodes and contractile rings by super-resolution fluorescence microscopy of live fission yeast. Proc. Natl. Acad. Sci. USA 2016, 113, E5876–E5885. [Google Scholar] [CrossRef]
- McDonald, N.A.; Lind, A.L.; Smith, S.E.; Li, R.; Gould, K.L. Nanoscale architecture of the Schizosaccharomyces pombe contractile ring. eLife 2017, 6, e28865. [Google Scholar] [CrossRef]
- Swulius, M.T.; Nguyen, L.T.; Ladinsky, M.S.; Ortega, D.R.; Aich, S.; Mishra, M.; Jensen, G.J. Structure of the fission yeast actomyosin ring during constriction. Proc. Natl. Acad. Sci. USA 2018, 115, E1455–E1464. [Google Scholar] [CrossRef]
- Fenix, A.M.; Taneja, N.; Buttler, C.A.; Lewis, J.; Van Engelenburg, S.B.; Ohi, R.; Burnette, D.T. Expansion and concatenation of non-muscle myosin IIA filaments drive cellular contractile system formation during interphase and mitosis. Mol. Biol. Cell 2016, 27, 1465–1478. [Google Scholar] [CrossRef]
- Henson, J.H.; Ditzler, C.E.; Germain, A.; Irwin, P.M.; Vogt, E.T.; Yang, S.; Wu, X.; Shuster, C.B. The ultrastructural organization of actin and myosin II filaments in the contractile ring: New support for an old model of cytokinesis. Mol. Biol. Cell 2017, 28, 613–623. [Google Scholar] [CrossRef]
- O’Shaughnessy, B.; Thiyagarajan, S. Mechanisms of contractile ring tension production and constriction. Biophys. Rev. 2018, 10, 1667–1681. [Google Scholar] [CrossRef]
- Proctor, S.A.; Minc, N.; Boudaoud, A.; Chang, F. Contributions of turgor pressure, the contractile ring, and septum assembly to forces in cytokinesis in fission yeast. Curr. Biol. 2012, 22, 1601–1608. [Google Scholar] [CrossRef]
- Bi, E.; Maddox, P.; Lew, D.J.; Salmon, E.D.; McMillan, J.N.; Yeh, E.; Pringle, J.R. Involvement of an actomyosin contractile ring in Saccharomyces cerevisiae cytokinesis. J. Cell Biol. 1998, 142, 1301–1312. [Google Scholar] [CrossRef]
- Chugh, P.; Paluch, E.K. The actin cortex at a glance. J. Cell Sci. 2018, 131, jcs186254. [Google Scholar] [CrossRef] [PubMed]
- Pollard, T.D. Nine unanswered questions about cytokinesis. J. Cell Biol. 2017, 216, 3007–3016. [Google Scholar] [CrossRef]
- Schroeder, T.E. The contractile ring. I. Fine structure of dividing mammalian (HeLa) cells and the effects of cytochalasin B. Z. Zellforsch. Mikrosk. Anat. 1970, 109, 431–449. [Google Scholar] [CrossRef] [PubMed]
- Krishan, A. Fine structure of cytochalasin-induced multinucleated cells. J. Ultrastruct. Res. 1971, 36, 191–204. [Google Scholar] [CrossRef]
- Schroeder, T.E. The contractile ring. II. Determining its brief existence, volumetric changes, and vital role in cleaving Arbacia eggs. J. Cell Biol. 1972, 53, 419–434. [Google Scholar] [CrossRef]
- Hamaguchi, Y.; Mabuchi, I. Effects of phalloidin microinjection and localization of fluorescein-labeled phalloidin in living sand dollar eggs. Cell Motil. 1982, 2, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Schatten, G.; Schatten, H.; Spector, I.; Cline, C.; Paweletz, N.; Simerly, C.; Petzelt, C. Latrunculin inhibits the microfilament-mediated processes during fertilization, cleavage and early development in sea urchins and mice. Exp. Cell Res. 1986, 166, 191–208. [Google Scholar] [CrossRef]
- Pollard, T.D.; Blanchoin, L.; Dyche Mullins, R. Molecular Mechanisms Controlling Actin Filament Dynamics in Nonmuscle Cells. Annu. Rev. Biophys. Biomol. Struct. 2000, 29, 545–576. [Google Scholar] [CrossRef] [PubMed]
- Blanchoin, L.; Boujemaa-Paterski, R.; Sykes, C.; Plastino, J. Actin dynamics, architecture, and mechanics in cell motility. Physiol. Rev. 2014, 94, 235–263. [Google Scholar] [CrossRef]
- Shekhar, S.; Pernier, J.; Carlier, M.-F. Regulators of actin filament barbed ends at a glance. J. Cell Sci. 2016, 129, 1085–1091. [Google Scholar] [CrossRef]
- Breitsprecher, D.; Goode, B.L. Formins at a glance. J. Cell Sci. 2013, 126, 1–7. [Google Scholar] [CrossRef]
- Kanellos, G.; Frame, M.C. Cellular functions of the ADF/cofilin family at a glance. J. Cell Sci. 2016, 129, 3211–3218. [Google Scholar] [CrossRef]
- Chan, F.-Y.; Silva, A.M.; Saramago, J.; Pereira-Sousa, J.; Brighton, H.E.; Pereira, M.; Oegema, K.; Gassmann, R.; Carvalho, A.X. The ARP2/3 complex prevents excessive formin activity during cytokinesis. Mol. Biol. Cell 2018, 30, 96–107. [Google Scholar] [CrossRef]
- Hartman, M.A.; Spudich, J.A. The myosin superfamily at a glance. J. Cell Sci. 2012, 125, 1627–1632. [Google Scholar] [CrossRef]
- Liu, X.; Shu, S.; Korn, E.D. Polymerization pathway of mammalian nonmuscle myosin 2s. Proc. Natl. Acad. Sci. USA 2018, 115, E7101–E7108. [Google Scholar] [CrossRef]
- Liu, X.; Billington, N.; Shu, S.; Yu, S.-H.; Piszczek, G.; Sellers, J.R.; Korn, E.D. Effect of ATP and regulatory light-chain phosphorylation on the polymerization of mammalian nonmuscle myosin II. Proc. Natl. Acad. Sci. USA 2017, 114, E6516–E6525. [Google Scholar] [CrossRef]
- Billington, N.; Wang, A.; Mao, J.; Adelstein, R.S.; Sellers, J.R. Characterization of Three Full-length Human Nonmuscle Myosin II Paralogs. J. Biol. Chem. 2013, 288, 33398–33410. [Google Scholar] [CrossRef]
- Shutova, M.S.; Spessott, W.A.; Giraudo, C.G.; Svitkina, T. Endogenous species of mammalian nonmuscle myosin IIA and IIB include activated monomers and heteropolymers. Curr. Biol. 2014, 24, 1958–1968. [Google Scholar] [CrossRef]
- Scholey, J.M.; Taylor, K.A.; Kendrick-Jones, J. Regulation of non-muscle myosin assembly by calmodulin-dependent light chain kinase. Nature 1980, 287, 233–235. [Google Scholar] [CrossRef]
- Scholey, J.M.; Smith, R.C.; Drenckhahn, D.; Groschel-Stewart, U.; Kendrick-Jones, J. Thymus myosin. Isolation and characterization of myosin from calf thymus and thymic lymphocytes, and studies on the effect of phosphorylation of its Mr = 20,000 light chain. J. Biol. Chem. 1982, 257, 7737–7745. [Google Scholar]
- Pollard, T.D. Structure and polymerization of Acanthamoeba myosin-II filaments. J. Cell Biol. 1982, 95, 816–825. [Google Scholar] [CrossRef]
- Niederman, R.; Pollard, T.D. Human platelet myosin. II. In vitro assembly and structure of myosin filaments. J. Cell Biol. 1975, 67, 72–92. [Google Scholar] [CrossRef]
- Verkhovsky, A.B.; Borisy, G.G. Non-sarcomeric mode of myosin II organization in the fibroblast lamellum. J. Cell Biol. 1993, 123, 637–652. [Google Scholar] [CrossRef]
- Geeves, M.A. Review: The ATPase mechanism of myosin and actomyosin. Biopolymers 2016, 105, 483–491. [Google Scholar] [CrossRef]
- Billington, N.; Beach, J.R.; Heissler, S.M.; Remmert, K.; Guzik-Lendrum, S.; Nagy, A.; Takagi, Y.; Shao, L.; Li, D.; Yang, Y.; et al. Myosin 18A Coassembles with Nonmuscle Myosin 2 to Form Mixed Bipolar Filaments. Curr. Biol. 2015, 25, 942–948. [Google Scholar] [CrossRef]
- Fujiwara, K.; Pollard, T.D. Fluorescent antibody localization of myosin in the cytoplasm, cleavage furrow, and mitotic spindle of human cells. J. Cell Biol. 1976, 71, 848–875. [Google Scholar] [CrossRef]
- Mabuchi, I.; Okuno, M. The effect of myosin antibody on the division of starfish blastomeres. J. Cell Biol. 1977, 74, 251–263. [Google Scholar] [CrossRef]
- De Lozanne, A.; Spudich, J.A. Disruption of the Dictyostelium myosin heavy chain gene by homologous recombination. Science 1987, 236, 1086–1091. [Google Scholar] [CrossRef]
- Guo, S.; Kemphues, K.J. A non-muscle myosin required for embryonic polarity in Caenorhabditis elegans. Nature 1996, 382, 455–458. [Google Scholar] [CrossRef]
- Straight, A.F.; Cheung, A.; Limouze, J.; Chen, I.; Westwood, N.J.; Sellers, J.R.; Mitchison, T.J. Dissecting Temporal and Spatial Control of Cytokinesis with a Myosin II Inhibitor. Science 2003, 299, 1743–1747. [Google Scholar] [CrossRef]
- Sanger, J.M.; Sanger, J.W. Banding and polarity of actin filaments in interphase and cleaving cells. J. Cell Biol. 1980, 86, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, T.E. Cytokinesis: Filaments in the cleavage furrow. Exp. Cell Res. 1968, 53, 272–276. [Google Scholar] [CrossRef]
- Schroeder, T.E. Actin in dividing cells: Contractile ring filaments bind heavy meromyosin. Proc. Natl. Acad. Sci. USA 1973, 70, 1688–1692. [Google Scholar] [CrossRef]
- Nunnally, M.H.; D’Angelo, J.M.; Craig, S.W. Filamin concentration in cleavage furrow and midbody region: Frequency of occurrence compared with that of alpha-actinin and myosin. J. Cell Biol. 1980, 87, 219–226. [Google Scholar] [CrossRef]
- Maupin, P.; Pollard, T.D. Arrangement of actin filaments and myosin-like filaments in the contractile ring and of actin-like filaments in the mitotic spindle of dividing HeLa cells. J. Ultrastruct. Mol. Struct. Res. 1986, 94, 92–103. [Google Scholar] [CrossRef]
- Davies, T.; Jordan, S.N.; Chand, V.; Sees, J.A.; Laband, K.; Carvalho, A.X.; Shirasu-Hiza, M.; Kovar, D.R.; Dumont, J.; Canman, J.C. High-Resolution Temporal Analysis Reveals a Functional Timeline for the Molecular Regulation of Cytokinesis. Dev. Cell 2014, 30, 209–223. [Google Scholar] [CrossRef]
- Ding, W.Y.; Ong, H.T.; Hara, Y.; Wongsantichon, J.; Toyama, Y.; Robinson, R.C.; Nédélec, F.; Zaidel-Bar, R. Plastin increases cortical connectivity to facilitate robust polarization and timely cytokinesis. J. Cell Biol. 2017, 216, 1371–1386. [Google Scholar] [CrossRef]
- Reichl, E.M.; Ren, Y.; Morphew, M.K.; Delannoy, M.; Effler, J.C.; Girard, K.D.; Divi, S.; Iglesias, P.A.; Kuo, S.C.; Robinson, D.N. Interactions between Myosin and Actin Crosslinkers Control Cytokinesis Contractility Dynamics and Mechanics. Curr. Biol. 2008, 18, 471–480. [Google Scholar] [CrossRef]
- Reymann, A.-C.; Staniscia, F.; Erzberger, A.; Salbreux, G.; Grill, S.W. Cortical flow aligns actin filaments to form a furrow. eLife 2016, 5, e17807. [Google Scholar] [CrossRef]
- Kishi, K.; Sasaki, T.; Kuroda, S.; Itoh, T.; Takai, Y. Regulation of cytoplasmic division of Xenopus embryo by rho p21 and its inhibitory GDP/GTP exchange protein (rho GDI). J. Cell Biol. 1993, 120, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Loria, A.; Longhini, K.M.; Glotzer, M. The RhoGAP domain of CYK-4 has an essential role in RhoA activation. Curr. Biol. 2012, 22, 213–219. [Google Scholar] [CrossRef]
- Otomo, T.; Otomo, C.; Tomchick, D.R.; Machius, M.; Rosen, M.K. Structural basis of Rho GTPase-mediated activation of the formin mDia1. Mol. Cell 2005, 18, 273–281. [Google Scholar] [CrossRef]
- Watanabe, S.; Ando, Y.; Yasuda, S.; Hosoya, H.; Watanabe, N.; Ishizaki, T.; Narumiya, S. mDia2 Induces the Actin Scaffold for the Contractile Ring and Stabilizes Its Position during Cytokinesis in NIH 3T3 Cells. Mol. Biol. Cell 2008, 19, 2328–2338. [Google Scholar] [CrossRef]
- Hu, S.; Dasbiswas, K.; Guo, Z.; Tee, Y.-H.; Thiagarajan, V.; Hersen, P.; Chew, T.-L.; Safran, S.A.; Zaidel-Bar, R.; Bershadsky, A.D. Long-range self-organization of cytoskeletal myosin II filament stacks. Nat. Cell Biol. 2017, 19, 133–141. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, Y.-L. Distinct pathways for the early recruitment of myosin II and actin to the cytokinetic furrow. Mol. Biol. Cell 2008, 19, 318–326. [Google Scholar] [CrossRef]
- Uehara, R.; Goshima, G.; Mabuchi, I.; Vale, R.D.; Spudich, J.A.; Griffis, E.R. Determinants of myosin II cortical localization during cytokinesis. Curr. Biol. 2010, 20, 1080–1085. [Google Scholar] [CrossRef] [PubMed]
- Spira, F.; Cuylen-Haering, S.; Mehta, S.; Samwer, M.; Reversat, A.; Verma, A.; Oldenbourg, R.; Sixt, M.; Gerlich, D.W. Cytokinesis in vertebrate cells initiates by contraction of an equatorial actomyosin network composed of randomly oriented filaments. eLife 2017, 6, e30867. [Google Scholar] [CrossRef]
- Kimura, K.; Ito, M.; Amano, M.; Chihara, K.; Fukata, Y.; Nakafuku, M.; Yamamori, B.; Feng, J.; Nakano, T.; Okawa, K.; et al. Regulation of myosin phosphatase by Rho and Rho-associated kinase (Rho-kinase). Science 1996, 273, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Khaliullin, R.N.; Green, R.A.; Shi, L.Z.; Gomez-Cavazos, J.S.; Berns, M.W.; Desai, A.; Oegema, K. A positive-feedback-based mechanism for constriction rate acceleration during cytokinesis in Caenorhabditis elegans. eLife 2018, 7, e36073. [Google Scholar] [CrossRef]
- Ren, Y.; Effler, J.C.; Norstrom, M.; Luo, T.; Firtel, R.A.; Iglesias, P.A.; Rock, R.S.; Robinson, D.N. Mechanosensing through cooperative interactions between myosin II and the actin crosslinker cortexillin I. Curr. Biol. 2009, 19, 1421–1428. [Google Scholar] [CrossRef]
- Laporte, D.; Coffman, V.C.; Lee, I.-J.; Wu, J.-Q. Assembly and architecture of precursor nodes during fission yeast cytokinesis. J. Cell Biol. 2011, 192, 1005–1021. [Google Scholar] [CrossRef]
- Wu, J.-Q.; Sirotkin, V.; Kovar, D.R.; Lord, M.; Beltzner, C.C.; Kuhn, J.R.; Pollard, T.D. Assembly of the cytokinetic contractile ring from a broad band of nodes in fission yeast. J. Cell Biol. 2006, 174, 391–402. [Google Scholar] [CrossRef]
- Stachowiak, M.R.; Laplante, C.; Chin, H.F.; Guirao, B.; Karatekin, E.; Pollard, T.D.; O’Shaughnessy, B. Mechanism of cytokinetic contractile ring constriction in fission yeast. Dev. Cell 2014, 29, 547–561. [Google Scholar] [CrossRef]
- Padmanabhan, A.; Bakka, K.; Sevugan, M.; Naqvi, N.I.; D’souza, V.; Tang, X.; Mishra, M.; Balasubramanian, M.K. IQGAP-related Rng2p organizes cortical nodes and ensures position of cell division in fission yeast. Curr. Biol. 2011, 21, 467–472. [Google Scholar] [CrossRef]
- Vavylonis, D.; Wu, J.-Q.; Hao, S.; O’Shaughnessy, B.; Pollard, T.D. Assembly mechanism of the contractile ring for cytokinesis by fission yeast. Science 2008, 319, 97–100. [Google Scholar] [CrossRef]
- Mendes Pinto, I.; Rubinstein, B.; Li, R. Force to divide: Structural and mechanical requirements for actomyosin ring contraction. Biophys. J. 2013, 105, 547–554. [Google Scholar] [CrossRef]
- Murrell, M.; Oakes, P.W.; Lenz, M.; Gardel, M.L. Forcing cells into shape: The mechanics of actomyosin contractility. Nat. Rev. Mol. Cell Biol. 2015, 16, 486–498. [Google Scholar] [CrossRef]
- Wollrab, V.; Thiagarajan, R.; Wald, A.; Kruse, K.; Riveline, D. Still and rotating myosin clusters determine cytokinetic ring constriction. Nat. Commun. 2016, 7, 11860. [Google Scholar] [CrossRef]
- Carvalho, A.; Desai, A.; Oegema, K. Structural Memory in the Contractile Ring Makes the Duration of Cytokinesis Independent of Cell Size. Cell 2009, 137, 926–937. [Google Scholar] [CrossRef]
- De La Cruz, E.M.; Gardel, M.L. Actin Mechanics and Fragmentation. J. Biol. Chem. 2015, 290, 17137–17144. [Google Scholar] [CrossRef]
- Lenz, M.; Thoresen, T.; Gardel, M.L.; Dinner, A.R. Contractile Units in Disordered Actomyosin Bundles Arise from F-Actin Buckling. Phys. Rev. Lett. 2012, 108, 238107. [Google Scholar] [CrossRef] [PubMed]
- Murrell, M.P.; Gardel, M.L. F-actin buckling coordinates contractility and severing in a biomimetic actomyosin cortex. Proc. Natl. Acad. Sci. USA 2012, 109, 20820–20825. [Google Scholar] [CrossRef]
- Vogel, S.K.; Petrasek, Z.; Heinemann, F.; Schwille, P. Myosin motors fragment and compact membrane-bound actin filaments. eLife 2013, 2, e00116. [Google Scholar] [CrossRef]
- Kamasaki, T.; Osumi, M.; Mabuchi, I. Three-dimensional arrangement of F-actin in the contractile ring of fission yeast. J. Cell Biol. 2007, 178, 765–771. [Google Scholar] [CrossRef]
- Bovellan, M.; Romeo, Y.; Biro, M.; Boden, A.; Chugh, P.; Yonis, A.; Vaghela, M.; Fritzsche, M.; Moulding, D.; Thorogate, R.; et al. Cellular control of cortical actin nucleation. Curr. Biol. 2014, 24, 1628–1635. [Google Scholar] [CrossRef]
- Dickinson, R.B.; Caro, L.; Purich, D.L. Force generation by cytoskeletal filament end-tracking proteins. Biophys. J. 2004, 87, 2838–2854. [Google Scholar] [CrossRef]
- Oelz, D.B.; Rubinstein, B.Y.; Mogilner, A. A Combination of Actin Treadmilling and Cross-Linking Drives Contraction of Random Actomyosin Arrays. Biophys. J. 2015, 109, 1818–1829. [Google Scholar] [CrossRef] [PubMed]
- Wollrab, V.; Belmonte, J.M.; Baldauf, L.; Leptin, M.; Nédeléc, F.; Koenderink, G.H. Polarity sorting drives remodeling of actin-myosin networks. J. Cell Sci. 2018, 132. [Google Scholar] [CrossRef]
- Descovich, C.P.; Cortes, D.B.; Ryan, S.; Nash, J.; Zhang, L.; Maddox, P.S.; Nedelec, F.; Maddox, A.S. Cross-linkers both drive and brake cytoskeletal remodeling and furrowing in cytokinesis. Mol. Biol. Cell 2018, 29, 622–631. [Google Scholar] [CrossRef]
- Allingham, J.S.; Smith, R.; Rayment, I. The structural basis of blebbistatin inhibition and specificity for myosin II. Nat. Struct. Mol. Biol. 2005, 12, 378–379. [Google Scholar] [CrossRef]
- Heissler, S.M.; Sellers, J.R. Four things to know about myosin light chains as reporters for non-muscle myosin-2 dynamics in live cells. Cytoskeleton 2015, 72, 65–70. [Google Scholar] [CrossRef]
- Liu, X.; Shu, S.; Billington, N.; Williamson, C.D.; Yu, S.; Brzeska, H.; Donaldson, J.G.; Sellers, J.R.; Korn, E.D. Mammalian Nonmuscle Myosin II Binds to Anionic Phospholipids with Concomitant Dissociation of the Regulatory Light Chain. J. Biol. Chem. 2016, 291, 24828–24837. [Google Scholar] [CrossRef]
- Vasquez, C.G.; Tworoger, M.; Martin, A.C. Dynamic myosin phosphorylation regulates contractile pulses and tissue integrity during epithelial morphogenesis. J. Cell Biol. 2014, 206, 435–450. [Google Scholar] [CrossRef]
- Palani, S.; Chew, T.G.; Ramanujam, S.; Kamnev, A.; Harne, S.; Chapa-y-Lazo, B.; Hogg, R.; Sevugan, M.; Mishra, M.; Gayathri, P.; et al. Motor Activity Dependent and Independent Functions of Myosin II Contribute to Actomyosin Ring Assembly and Contraction in Schizosaccharomyces pombe. Curr. Biol. 2017, 27, 751–757. [Google Scholar] [CrossRef]
- Shimada, T.; Sasaki, N.; Ohkura, R.; Sutoh, K. Alanine Scanning Mutagenesis of the Switch I Region in the ATPase Site of Dictyostelium discoideum Myosin II. Biochemistry 1997, 36, 14037–14043. [Google Scholar] [CrossRef]
- Ma, X.; Kovács, M.; Conti, M.A.; Wang, A.; Zhang, Y.; Sellers, J.R.; Adelstein, R.S. Nonmuscle myosin II exerts tension but does not translocate actin in vertebrate cytokinesis. Proc. Natl. Acad. Sci. USA 2012, 109, 4509–4514. [Google Scholar] [CrossRef]
- Osorio, D.S.; Chan, F.Y.; Saramago, J.; Leite, J.; Silva, A.M.; Sobral, A.F.; Gassmann, R.; Carvalho, A.X. Flow-independent accumulation of motor-competent non-muscle myosin II in the contractile ring is essential for cytokinesis. bioRxiv 2018, 333286. [Google Scholar] [CrossRef]
- Heissler, S.; Billington, N.; Ma, X.; Adelstein, R.; Sellers, J. Tools to Study Nonmuscle Myosin-2 Motor Function Revisited. Biophys. J. 2018, 114, 318a. [Google Scholar] [CrossRef]
- Hu, A.; Wang, F.; Sellers, J.R. Mutations in human nonmuscle myosin IIA found in patients with May-Hegglin anomaly and Fechtner syndrome result in impaired enzymatic function. J. Biol. Chem. 2002, 277, 46512–46517. [Google Scholar] [CrossRef]
- Kanada, M.; Nagasaki, A.; Uyeda, T.Q.P. Adhesion-dependent and Contractile Ring-independent Equatorial Furrowing during Cytokinesis in Mammalian Cells. Mol. Biol. Cell 2005, 16, 3865–3872. [Google Scholar] [CrossRef]
- Nagasaki, A.; Kanada, M.; Uyeda, T.Q. Cell adhesion molecules regulate contractile ring-independent cytokinesis in Dictyostelium discoideum. Cell Res. 2009, 19, 236–246. [Google Scholar] [CrossRef]
- Dix, C.L.; Matthews, H.K.; Uroz, M.; McLaren, S.; Wolf, L.; Heatley, N.; Win, Z.; Almada, P.; Henriques, R.; Boutros, M.; et al. The Role of Mitotic Cell-Substrate Adhesion Re-modeling in Animal Cell Division. Dev. Cell 2018, 45, 132–145.e3. [Google Scholar] [CrossRef]
- Fang, X.; Luo, J.; Nishihama, R.; Wloka, C.; Dravis, C.; Travaglia, M.; Iwase, M.; Vallen, E.A.; Bi, E. Biphasic targeting and cleavage furrow ingression directed by the tail of a myosin II. J. Cell Biol. 2010, 191, 1333–1350. [Google Scholar] [CrossRef]
- Lord, M.; Laves, E.; Pollard, T.D. Cytokinesis Depends on the Motor Domains of Myosin-II in Fission Yeast but Not in Budding Yeast. Mol. Biol. Cell 2005, 16, 5346–5355. [Google Scholar] [CrossRef]
- Mendes Pinto, I.; Rubinstein, B.; Kucharavy, A.; Unruh, J.R.; Li, R. Actin depolymerization drives actomyosin ring contraction during budding yeast cytokinesis. Dev. Cell 2012, 22, 1247–1260. [Google Scholar] [CrossRef]
- Wilson, C.A.; Tsuchida, M.A.; Allen, G.M.; Barnhart, E.L.; Applegate, K.T.; Yam, P.T.; Ji, L.; Keren, K.; Danuser, G.; Theriot, J.A. Myosin II contributes to cell-scale actin network treadmilling through network disassembly. Nature 2010, 465, 373–377. [Google Scholar] [CrossRef]
- Reymann, A.-C.; Boujemaa-Paterski, R.; Martiel, J.-L.; Guérin, C.; Cao, W.; Chin, H.F.; De La Cruz, E.M.; Théry, M.; Blanchoin, L. Actin network architecture can determine myosin motor activity. Science 2012, 336, 1310–1314. [Google Scholar] [CrossRef]
- Turlier, H.; Audoly, B.; Prost, J.; Joanny, J.-F. Furrow Constriction in Animal Cell Cytokinesis. Biophys. J. 2014, 106, 114–123. [Google Scholar] [CrossRef]
- Zhang, W.; Robinson, D.N. Balance of actively generated contractile and resistive forces controls cytokinesis dynamics. Proc. Natl. Acad. Sci. USA 2005, 102, 7186–7191. [Google Scholar] [CrossRef]
- Cooper, J.A. Effects of cytochalasin and phalloidin on actin. J. Cell Biol. 1987, 105, 1473–1478. [Google Scholar] [CrossRef]
- Kashman, Y.; Groweiss, A.; Shmueli, U. Latrunculin, a new 2-thiazolidinone macrolide from the marine sponge latrunculia magnifica. Tetrahedron Lett. 1980, 21, 3629–3632. [Google Scholar] [CrossRef]
- Morton, W.M.; Ayscough, K.R.; McLaughlin, P.J. Latrunculin alters the actin-monomer subunit interface to prevent polymerization. Nat. Cell Biol. 2000, 2, 376–378. [Google Scholar] [CrossRef]
- Fujiwara, I.; Zweifel, M.E.; Courtemanche, N.; Pollard, T.D. Latrunculin A Accelerates Actin Filament Depolymerization in Addition to Sequestering Actin Monomers. Curr. Biol. 2018, 28, 3183–3192.e2. [Google Scholar] [CrossRef]
- Zabriskie, T.M.; Klocke, J.A.; Ireland, C.M.; Marcus, A.H.; Molinski, T.F.; Faulkner, D.J.; Xu, C.; Clardy, J. Jaspamide, a modified peptide from a Jaspis sponge, with insecticidal and antifungal activity. J. Am. Chem. Soc. 1986, 108, 3123–3124. [Google Scholar] [CrossRef]
- Crews, P.; Manes, L.V.; Boehler, M. Jasplakinolide, a cyclodepsipeptide from the marine sponge, Jaspis SP. Tetrahedron Lett. 1986, 27, 2797–2800. [Google Scholar] [CrossRef]
- Bubb, M.R.; Spector, I.; Beyer, B.B.; Fosen, K.M. Effects of jasplakinolide on the kinetics of actin polymerization. An explanation for certain in vivo observations. J. Biol. Chem. 2000, 275, 5163–5170. [Google Scholar] [CrossRef] [PubMed]
- Murthy, K.; Wadsworth, P. Myosin-II-dependent localization and dynamics of F-actin during cytokinesis. Curr. Biol. 2005, 15, 724–731. [Google Scholar] [CrossRef]
- Chew, T.G.; Huang, J.; Palani, S.; Sommese, R.; Kamnev, A.; Hatano, T.; Gu, Y.; Oliferenko, S.; Sivaramakrishnan, S.; Balasubramanian, M.K. Actin turnover maintains actin filament homeostasis during cytokinetic ring contraction. J. Cell Biol. 2017, 216, 2657–2667. [Google Scholar] [CrossRef]
- Carvalho, A.; Olson, S.K.; Gutierrez, E.; Zhang, K.; Noble, L.B.; Zanin, E.; Desai, A.; Groisman, A.; Oegema, K. Acute Drug Treatment in the Early C. elegans Embryo. PLoS ONE 2011, 6, e24656. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.M.; Osório, D.S.; Pereira, A.J.; Maiato, H.; Pinto, I.M.; Rubinstein, B.; Gassmann, R.; Telley, I.A.; Carvalho, A.X. Robust gap repair in the contractile ring ensures timely completion of cytokinesis. J. Cell Biol. 2016, 215, 789–799. [Google Scholar] [CrossRef]
- O’Connell, C.B.; Warner, A.K.; Wang, Y. Distinct roles of the equatorial and polar cortices in the cleavage of adherent cells. Curr. Biol. 2001, 11, 702–707. [Google Scholar] [CrossRef]
- Motegi, F.; Sugimoto, A. Sequential functioning of the ECT-2 RhoGEF, RHO-1 and CDC-42 establishes cell polarity in Caenorhabditis elegans embryos. Nat. Cell Biol. 2006, 8, 978–985. [Google Scholar] [CrossRef]
- Watanabe, S.; Okawa, K.; Miki, T.; Sakamoto, S.; Morinaga, T.; Segawa, K.; Arakawa, T.; Kinoshita, M.; Ishizaki, T.; Narumiya, S. Rho and anillin-dependent control of mDia2 localization and function in cytokinesis. Mol. Biol. Cell 2010, 21, 3193–3204. [Google Scholar] [CrossRef] [PubMed]
- Gunsalus, K.C.; Bonaccorsi, S.; Williams, E.; Verni, F.; Gatti, M.; Goldberg, M.L. Mutations in twinstar, a Drosophila gene encoding a cofilin/ADF homologue, result in defects in centrosome migration and cytokinesis. J. Cell Biol. 1995, 131, 1243–1259. [Google Scholar] [CrossRef] [PubMed]
- Somma, M.P.; Fasulo, B.; Cenci, G.; Cundari, E.; Gatti, M.; McIntosh, J.R. Molecular Dissection of Cytokinesis by RNA Interference inDrosophila Cultured Cells. Mol. Biol. Cell 2002, 13, 2448–2460. [Google Scholar] [CrossRef]
- Hotulainen, P.; Paunola, E.; Vartiainen, M.K.; Lappalainen, P. Actin-depolymerizing Factor and Cofilin-1 Play Overlapping Roles in Promoting Rapid F-Actin Depolymerization in Mammalian Nonmuscle Cells. Mol. Biol. Cell 2005, 16, 649–664. [Google Scholar] [CrossRef]
- Nakano, K.; Mabuchi, I. Actin-depolymerizing protein Adf1 is required for formation and maintenance of the contractile ring during cytokinesis in fission yeast. Mol. Biol. Cell 2006, 17, 1933–1945. [Google Scholar] [CrossRef]
- Mishra, M.; Kashiwazaki, J.; Takagi, T.; Srinivasan, R.; Huang, Y.; Balasubramanian, M.K.; Mabuchi, I. In vitro contraction of cytokinetic ring depends on myosin II but not on actin dynamics. Nat. Cell Biol. 2013, 15, 853–859. [Google Scholar] [CrossRef]
- Djinović-Carugo, K.; Young, P.; Gautel, M.; Saraste, M. Structure of the alpha-actinin rod: Molecular basis for cross-linking of actin filaments. Cell 1999, 98, 537–546. [Google Scholar] [CrossRef]
- Klein, M.G.; Shi, W.; Ramagopal, U.; Tseng, Y.; Wirtz, D.; Kovar, D.R.; Staiger, C.J.; Almo, S.C. Structure of the actin crosslinking core of fimbrin. Structure 2004, 12, 999–1013. [Google Scholar] [CrossRef]
- Stossel, T.P.; Condeelis, J.; Cooley, L.; Hartwig, J.H.; Noegel, A.; Schleicher, M.; Shapiro, S.S. Filamins as integrators of cell mechanics and signalling. Nat. Rev. Mol. Cell Biol. 2001, 2, 138–145. [Google Scholar] [CrossRef]
- Coleman, T.R.; Fishkind, D.J.; Mooseker, M.S.; Morrow, J.S. Functional diversity among spectrin isoforms. Cell Motil. Cytoskelet. 1989, 12, 225–247. [Google Scholar] [CrossRef]
- Kinoshita, M.; Field, C.M.; Coughlin, M.L.; Straight, A.F.; Mitchison, T.J. Self- and actin-templated assembly of Mammalian septins. Dev. Cell 2002, 3, 791–802. [Google Scholar] [CrossRef]
- Jananji, S.; Risi, C.; Lindamulage, I.K.S.; Picard, L.-P.; Van Sciver, R.; Laflamme, G.; Albaghjati, A.; Hickson, G.R.X.; Kwok, B.H.; Galkin, V.E. Multimodal and Polymorphic Interactions between Anillin and Actin: Their Implications for Cytokinesis. J. Mol. Biol. 2017, 429, 715–731. [Google Scholar] [CrossRef]
- Mavrakis, M.; Azou-Gros, Y.; Tsai, F.-C.; Alvarado, J.; Bertin, A.; Iv, F.; Kress, A.; Brasselet, S.; Koenderink, G.H.; Lecuit, T. Septins promote F-actin ring formation by crosslinking actin filaments into curved bundles. Nat. Cell Biol. 2014, 16, 322–334. [Google Scholar] [CrossRef]
- Wu, J.Q.; Bähler, J.; Pringle, J.R. Roles of a fimbrin and an alpha-actinin-like protein in fission yeast cell polarization and cytokinesis. Mol. Biol. Cell 2001, 12, 1061–1077. [Google Scholar] [CrossRef]
- Fujiwara, K.; Porter, M.E.; Pollard, T.D. Alpha-actinin localization in the cleavage furrow during cytokinesis. J. Cell Biol. 1978, 79, 268–275. [Google Scholar] [CrossRef]
- Watanabe, A.; Kurasawa, Y.; Watanabe, Y.; Numata, O. A new Tetrahymena actin-binding protein is localized in the division furrow. J. Biochem. 1998, 123, 607–613. [Google Scholar] [CrossRef]
- Watanabe, A.; Yonemura, I.; Gonda, K.; Numata, O. Cloning and Sequencing of the Gene for a Tetrahymena Fimbrin-Like Protein. J. Biochem. 2000, 127, 85–94. [Google Scholar] [CrossRef]
- Nakano, K.; Satoh, K.; Morimatsu, A.; Ohnuma, M.; Mabuchi, I. Interactions among a fimbrin, a capping protein, and an actin-depolymerizing factor in organization of the fission yeast actin cytoskeleton. Mol. Biol. Cell 2001, 12, 3515–3526. [Google Scholar] [CrossRef]
- Field, C.M.; Alberts, B.M. Anillin, a contractile ring protein that cycles from the nucleus to the cell cortex. J. Cell Biol. 1995, 131, 165–178. [Google Scholar] [CrossRef]
- Maddox, A.S.; Lewellyn, L.; Desai, A.; Oegema, K. Anillin and the Septins Promote Asymmetric Ingression of the Cytokinetic Furrow. Dev. Cell 2007, 12, 827–835. [Google Scholar] [CrossRef]
- Mukhina, S.; Wang, Y.-L.; Murata-Hori, M. Alpha-actinin is required for tightly regulated remodeling of the actin cortical network during cytokinesis. Dev. Cell 2007, 13, 554–565. [Google Scholar] [CrossRef]
- Piekny, A.J.; Glotzer, M. Anillin is a scaffold protein that links RhoA, actin, and myosin during cytokinesis. Curr. Biol. 2008, 18, 30–36. [Google Scholar] [CrossRef]
- Faix, J.; Steinmetz, M.; Boves, H.; Kammerer, R.A.; Lottspeich, F.; Mintert, U.; Murphy, J.; Stock, A.; Aebi, U.; Gerisch, G. Cortexillins, major determinants of cell shape and size, are actin-bundling proteins with a parallel coiled-coil tail. Cell 1996, 86, 631–642. [Google Scholar] [CrossRef]
- Maddox, A.S.; Habermann, B.; Desai, A.; Oegema, K. Distinct roles for two C. elegans anillins in the gonad and early embryo. Development 2005, 132, 2837–2848. [Google Scholar] [CrossRef]
- Robinson, D.N.; Spudich, J.A. Dynacortin, a genetic link between equatorial contractility and global shape control discovered by library complementation of a Dictyostelium discoideum cytokinesis mutant. J. Cell Biol. 2000, 150, 823–838. [Google Scholar] [CrossRef]
- Laporte, D.; Ojkic, N.; Vavylonis, D.; Wu, J.-Q. α-Actinin and fimbrin cooperate with myosin II to organize actomyosin bundles during contractile-ring assembly. Mol. Biol. Cell 2012, 23, 3094–3110. [Google Scholar] [CrossRef]
- Li, Y.; Christensen, J.R.; Homa, K.E.; Hocky, G.M.; Fok, A.; Sees, J.A.; Voth, G.A.; Kovar, D.R. The F-actin bundler α-actinin Ain1 is tailored for ring assembly and constriction during cytokinesis in fission yeast. Mol. Biol. Cell 2016, 27, 1821–1833. [Google Scholar] [CrossRef]
- Srivastava, V.; Robinson, D.N. Mechanical Stress and Network Structure Drive Protein Dynamics during Cytokinesis. Curr. Biol. 2015, 25, 663–670. [Google Scholar] [CrossRef]
- Effler, J.C.; Kee, Y.-S.; Berk, J.M.; Tran, M.N.; Iglesias, P.A.; Robinson, D.N. Mitosis-specific mechanosensing and contractile-protein redistribution control cell shape. Curr. Biol. 2006, 16, 1962–1967. [Google Scholar] [CrossRef]
- Orlova, A.; Egelman, E.H. Cooperative rigor binding of myosin to actin is a function of F-actin structure. J. Mol. Biol. 1997, 265, 469–474. [Google Scholar] [CrossRef]
- Luo, T.; Mohan, K.; Srivastava, V.; Ren, Y.; Iglesias, P.A.; Robinson, D.N. Understanding the cooperative interaction between myosin II and actin cross-linkers mediated by actin filaments during mechanosensation. Biophys. J. 2012, 102, 238–247. [Google Scholar] [CrossRef]
- Ennomani, H.; Letort, G.; Guérin, C.; Martiel, J.-L.; Cao, W.; Nédélec, F.; De La Cruz, E.M.; Théry, M.; Blanchoin, L. Architecture and Connectivity Govern Actin Network Contractility. Curr. Biol. 2016, 26, 616–626. [Google Scholar] [CrossRef]
- Kruse, K.; Joanny, J.F.; Jülicher, F.; Prost, J.; Sekimoto, K. Generic theory of active polar gels: A paradigm for cytoskeletal dynamics. Eur. Phys. J. E. Soft Matter 2005, 16, 5–16. [Google Scholar] [CrossRef]
- Joanny, J.F.; Jülicher, F.; Kruse, K.; Prost, J. Hydrodynamic theory for multi-component active polar gels. New J. Phys. 2007, 9, 422. [Google Scholar] [CrossRef]
- Prost, J.; Jülicher, F.; Joanny, J.-F. Active gel physics. Nat. Phys. 2015, 11, 111–117. [Google Scholar] [CrossRef]
- Kruse, K.; Joanny, J.F.; Jülicher, F.; Prost, J. Contractility and retrograde flow in lamellipodium motion. Phys. Biol. 2006, 3, 130–137. [Google Scholar] [CrossRef]
- Salbreux, G.; Prost, J.; Joanny, J.F. Hydrodynamics of cellular cortical flows and the formation of contractile rings. Phys. Rev. Lett. 2009, 103, 058102. [Google Scholar] [CrossRef]
- Marsland, D.; Landau, J.V. The mechanisms of cytokinesis: Temperature-pressure studies on the cortical gel system in various marine eggs. J. Exp. Zool. 1954, 125, 507–539. [Google Scholar] [CrossRef]
- Mitchison, J.M.; Swann, M.M. The mechanical properties of the cell surface: III. The sea-urchin egg from fertilization to cleavage. J. Exp. Biol. 1955, 32, 734–750. [Google Scholar]
- Wolpert, L. Some problems of cleavage in relation to the cell membrane. In Cell Growth and Cell Division; Academic Press: New York, NY, USA, 1963; pp. 277–298. [Google Scholar]
- Wolpert, L. The mechanical properties of the membrane of the sea urchin egg during cleavage. Exp. Cell Res. 1966, 41, 385–396. [Google Scholar] [CrossRef]
- Rappaport, R. Cell Division: Direct Measurement of Maximum Tension Exerted by Furrow of Echinoderm Eggs. Science 1967, 156, 1241–1243. [Google Scholar] [CrossRef]
- Hiramoto, Y. Observations and measurements of sea urchin eggs with a centrifuge microscope. J. Cell. Physiol. 1967, 69, 219–230. [Google Scholar] [CrossRef]
- Hiramoto, Y. The mechanics and mechanism of cleavage in the sea-urchin egg. Symp. Soc. Exp. Biol. 1968, 22, 311–327. [Google Scholar]
- Yoneda, M.; Dan, K. Tension at the surface of the dividing sea-urchin egg. J. Exp. Biol. 1972, 57, 575–587. [Google Scholar]
- Mitchison, J.M. Cell membranes and cell division. Symp. Soc. Exp. Biol. 1952, 6, 105–127. [Google Scholar]
- Swann, M.M.; Mitchison, J.M. The mechanism of cleavage in animal cells. Biol. Rev. Camb. Philos. Soc. 1958, 33, 103–135. [Google Scholar] [CrossRef]
- Marsland, D. The mechanisms of cell division; temperature-pressure experiments on the cleaving eggs of Arbacia punctulata. J. Cell. Comp. Physiol. 1950, 36, 205–227. [Google Scholar] [CrossRef]
- Marsland, D. Protoplasmic contractility in relation to cytokinesis. Pubblicazioni della Stazione Zoologica di Napoli 1956, 28, 182–203. [Google Scholar]
- Marsland, D. Protoplasmic Contractility in Relation to Gel Structure: Temperature—Pressure Experiments on Cytokinesis and Amoeboid Movement. In International Review of Cytology; Elsevier: Amsterdam, The Netherlands, 1956; Volume 5, pp. 199–227. [Google Scholar]
- Wolpert, L. The mechanics and mechanism of cleavage. Int. Rev. Cytol. 1960, 10, 163–216. [Google Scholar]
- Greenspan, H.P. On the Deformation of a Viscous Droplet Caused by Variable Surface Tension. Stud. Appl. Math. 1977, 57, 45–58. [Google Scholar] [CrossRef]
- Akkas, N.; Barez, F.; Goldsmith, W. Elastic wave propagation in an exponential rod. Int. J. Mech. Sci. 1980, 22, 199–208. [Google Scholar] [CrossRef]
- White, J.G.; Borisy, G.G. On the mechanisms of cytokinesis in animal cells. J. Theor. Biol. 1983, 101, 289–316. [Google Scholar] [CrossRef]
- Zinemanas, D.; Nir, A. Fluid Mechanical Simulations of Cell Furrowing Due to Anisotropic Surface Forces. In Biomechanics of Cell Division; Akkas, N., Ed.; Springer US: Boston, MA, USA, 1987; pp. 281–305. ISBN 9781468412710. [Google Scholar]
- Zinemanas, D.; Nir, A. Surface viscoelastic effects in cell cleavage. J. Biomech. 1990, 23, 417–424. [Google Scholar] [CrossRef]
- He, X.; Dembo, M. A Dynamical Model of Cell Division. In Dynamics of Cell and Tissue Motion; Alt, W., Deutsch, A., Dunn, G.A., Eds.; Birkhäuser: Basel, Switzerland, 1997; pp. 55–65. ISBN 9783034889162. [Google Scholar]
- Zumdieck, A.; Cosentino Lagomarsino, M.; Tanase, C.; Kruse, K.; Mulder, B.; Dogterom, M.; Jülicher, F. Continuum description of the cytoskeleton: Ring formation in the cell cortex. Phys. Rev. Lett. 2005, 95, 258103. [Google Scholar] [CrossRef]
- Sain, A.; Inamdar, M.M.; Jülicher, F. Dynamic Force Balances and Cell Shape Changes during Cytokinesis. Phys. Rev. Lett. 2015, 114, 048102. [Google Scholar] [CrossRef]
- Dorn, J.F.; Zhang, L.; Phi, T.-T.; Lacroix, B.; Maddox, P.S.; Liu, J.; Maddox, A.S. A theoretical model of cytokinesis implicates feedback between membrane curvature and cytoskeletal organization in asymmetric cytokinetic furrowing. Mol. Biol. Cell 2016, 27, 1286–1299. [Google Scholar] [CrossRef]
- Biron, D.; Alvarez-Lacalle, E.; Tlusty, T.; Moses, E. Molecular model of the contractile ring. Phys. Rev. Lett. 2005, 95, 098102. [Google Scholar] [CrossRef]
- Zumdieck, A.; Kruse, K.; Bringmann, H.; Hyman, A.A.; Jülicher, F. Stress Generation and Filament Turnover during Actin Ring Constriction. PLoS ONE 2007, 2, e696. [Google Scholar] [CrossRef]
- Carlsson, A.E. Contractile stress generation by actomyosin gels. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2006, 74, 051912. [Google Scholar] [CrossRef]
- Rappaport, R. Tensiometric studies of cytokinesis in cleaving sand dollar eggs. J. Exp. Zool. 1977, 201, 375–378. [Google Scholar] [CrossRef]
- Jochova, J.; Rupe, I.; Streiblova, E. F-actin contractile rings in protoplasts of the yeast. Cell Biol. Int. Rep. 1991, 15, 607–610. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Swulius, M.T.; Aich, S.; Mishra, M.; Jensen, G.J. Coarse-grained simulations of actomyosin rings point to a nodeless model involving both unipolar and bipolar myosins. Mol. Biol. Cell 2018, 29, 1318–1331. [Google Scholar] [CrossRef]
- Surcel, A.; Kee, Y.-S.; Luo, T.; Robinson, D.N. Cytokinesis through biochemical-mechanical feedback loops. Semin. Cell Dev. Biol. 2010, 21, 866–873. [Google Scholar] [CrossRef]
- Poirier, C.C.; Ng, W.P.; Robinson, D.N.; Iglesias, P.A. Deconvolution of the cellular force-generating subsystems that govern cytokinesis furrow ingression. PLoS Comput. Biol. 2012, 8, e1002467. [Google Scholar] [CrossRef]
- Bray, D.; White, J.G. Cortical flow in animal cells. Science 1988, 239, 883–888. [Google Scholar] [CrossRef]
- Dan, K.; Dan, J.C.; Yanagita, T. Behaviour of the Cell Surface During Cleavage. II. Cytologia 1938, 8, 521–531. [Google Scholar] [CrossRef]
- Dan, K.; Ono, T. A method of computation of the surface area of the cell. Dev. Growth Differ. 1954, 2, 87–99. [Google Scholar] [CrossRef]
- Dan, K.; Yanagita, T.; Sugiyama, M. Behavior of the cell surface during cleavage. I. Protoplasma 1937, 28, 66–81. [Google Scholar] [CrossRef]
- Ellis-Davies, G.C.R. Caged compounds: Photorelease technology for control of cellular chemistry and physiology. Nat. Methods 2007, 4, 619–628. [Google Scholar] [CrossRef]
- Valon, L.; Marín-Llauradó, A.; Wyatt, T.; Charras, G.; Trepat, X. Optogenetic control of cellular forces and mechanotransduction. Nat. Commun. 2017, 8, 14396. [Google Scholar] [CrossRef]
- Fielmich, L.-E.; Schmidt, R.; Dickinson, D.J.; Goldstein, B.; Akhmanova, A.; van den Heuvel, S. Optogenetic dissection of mitotic spindle positioning in vivo. eLife 2018, 7, e38198. [Google Scholar] [CrossRef] [PubMed]
- Girard, K.D.; Chaney, C.; Delannoy, M.; Kuo, S.C.; Robinson, D.N. Dynacortin contributes to cortical viscoelasticity and helps define the shape changes of cytokinesis. EMBO J. 2004, 23, 1536–1546. [Google Scholar] [CrossRef]
- Cartagena-Rivera, A.X.; Logue, J.S.; Waterman, C.M.; Chadwick, R.S. Actomyosin Cortical Mechanical Properties in Nonadherent Cells Determined by Atomic Force Microscopy. Biophys. J. 2016, 110, 2528–2539. [Google Scholar] [CrossRef]
- Fritzsche, M.; Erlenkämper, C.; Moeendarbary, E.; Charras, G.; Kruse, K. Actin kinetics shapes cortical network structure and mechanics. Sci. Adv. 2016, 2, e1501337. [Google Scholar] [CrossRef]
- Olson, S.K.; Greenan, G.; Desai, A.; Müller-Reichert, T.; Oegema, K. Hierarchical assembly of the eggshell and permeability barrier in C. elegans. J. Cell Biol. 2012, 198, 731–748. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Nag, S.; Pollard, T.D. Formins filter modified actin subunits during processive elongation. J. Struct. Biol. 2012, 177, 32–39. [Google Scholar] [CrossRef]
- Robin, F.B.; McFadden, W.M.; Yao, B.; Munro, E.M. Single-molecule analysis of cell surface dynamics in Caenorhabditis elegans embryos. Nat. Methods 2014, 11, 677–682. [Google Scholar] [CrossRef]
- Melak, M.; Plessner, M.; Grosse, R. Actin visualization at a glance. J. Cell Sci. 2017, 130, 525–530. [Google Scholar] [CrossRef]
- Keppler, A.; Gendreizig, S.; Gronemeyer, T.; Pick, H.; Vogel, H.; Johnsson, K. A general method for the covalent labeling of fusion proteins with small molecules in vivo. Nat. Biotechnol. 2003, 21, 86–89. [Google Scholar] [CrossRef]
- Resch-Genger, U.; Grabolle, M.; Cavaliere-Jaricot, S.; Nitschke, R.; Nann, T. Quantum dots versus organic dyes as fluorescent labels. Nat. Methods 2008, 5, 763–775. [Google Scholar] [CrossRef]
- Chang, Y.-P.; Pinaud, F.; Antelman, J.; Weiss, S. Tracking bio-molecules in live cells using quantum dots. J. Biophotonics 2008, 1, 287–298. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Drugs | Source | Binding Site | Overall Effect on F-actin Networks | References |
|---|---|---|---|---|
| Cytochalasins (Cytochalasin D is most commonly used) | Fungi | F-actin filaments | Cap barbed ends preventing assembly and disassembly of monomers at that end. Prevent polymerization. | Reviewed in [112] |
| Latrunculins (Latrunculin A is most commonly used) | Marine sponges | Actin monomers | Prevent polymerization by sequestering monomers, increase depolymerization at both filament ends and promote filament severing | [113,114,115] |
| Jasplakinolide | Marine sponges | F-actin filaments | Stabilizes F-actin filaments, induces polymerization of actin into amorphous masses | [116,117,118] |
| Phalloidin | Fungi | F-actin filaments | Stabilizes F-actin filaments | Reviewed in [112] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leite, J.; Osorio, D.S.; Sobral, A.F.; Silva, A.M.; Carvalho, A.X. Network Contractility during Cytokinesis—From Molecular to Global Views. Biomolecules 2019, 9, 194. https://doi.org/10.3390/biom9050194
Leite J, Osorio DS, Sobral AF, Silva AM, Carvalho AX. Network Contractility during Cytokinesis—From Molecular to Global Views. Biomolecules. 2019; 9(5):194. https://doi.org/10.3390/biom9050194
Chicago/Turabian StyleLeite, Joana, Daniel Sampaio Osorio, Ana Filipa Sobral, Ana Marta Silva, and Ana Xavier Carvalho. 2019. "Network Contractility during Cytokinesis—From Molecular to Global Views" Biomolecules 9, no. 5: 194. https://doi.org/10.3390/biom9050194
APA StyleLeite, J., Osorio, D. S., Sobral, A. F., Silva, A. M., & Carvalho, A. X. (2019). Network Contractility during Cytokinesis—From Molecular to Global Views. Biomolecules, 9(5), 194. https://doi.org/10.3390/biom9050194