
Habitat Connectivity for the Conservation of Small Ungulates in A Human-Dominated Landscape

 and
and
Abstract
:1. Introduction
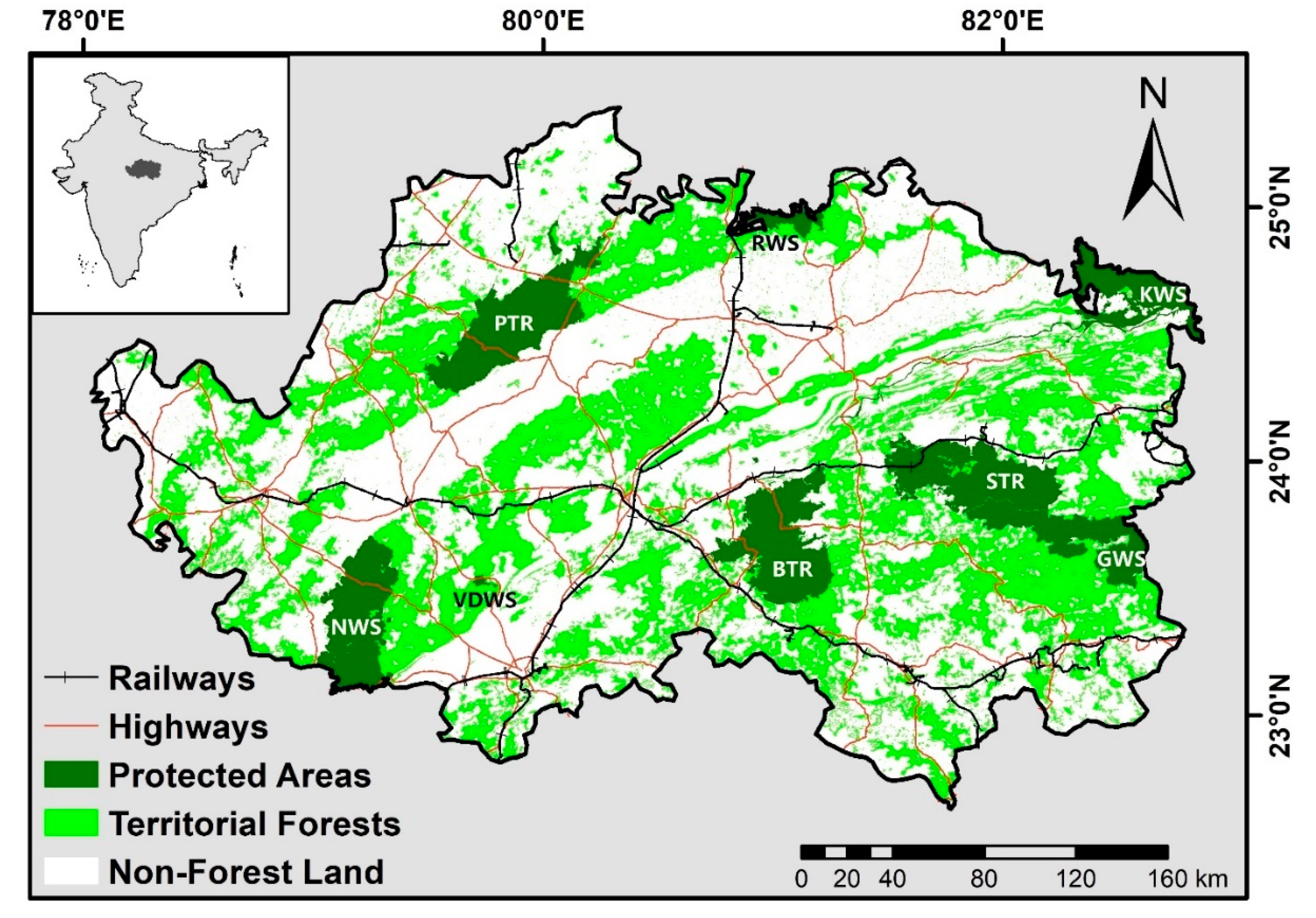
2. Study Area
3. Focal Species
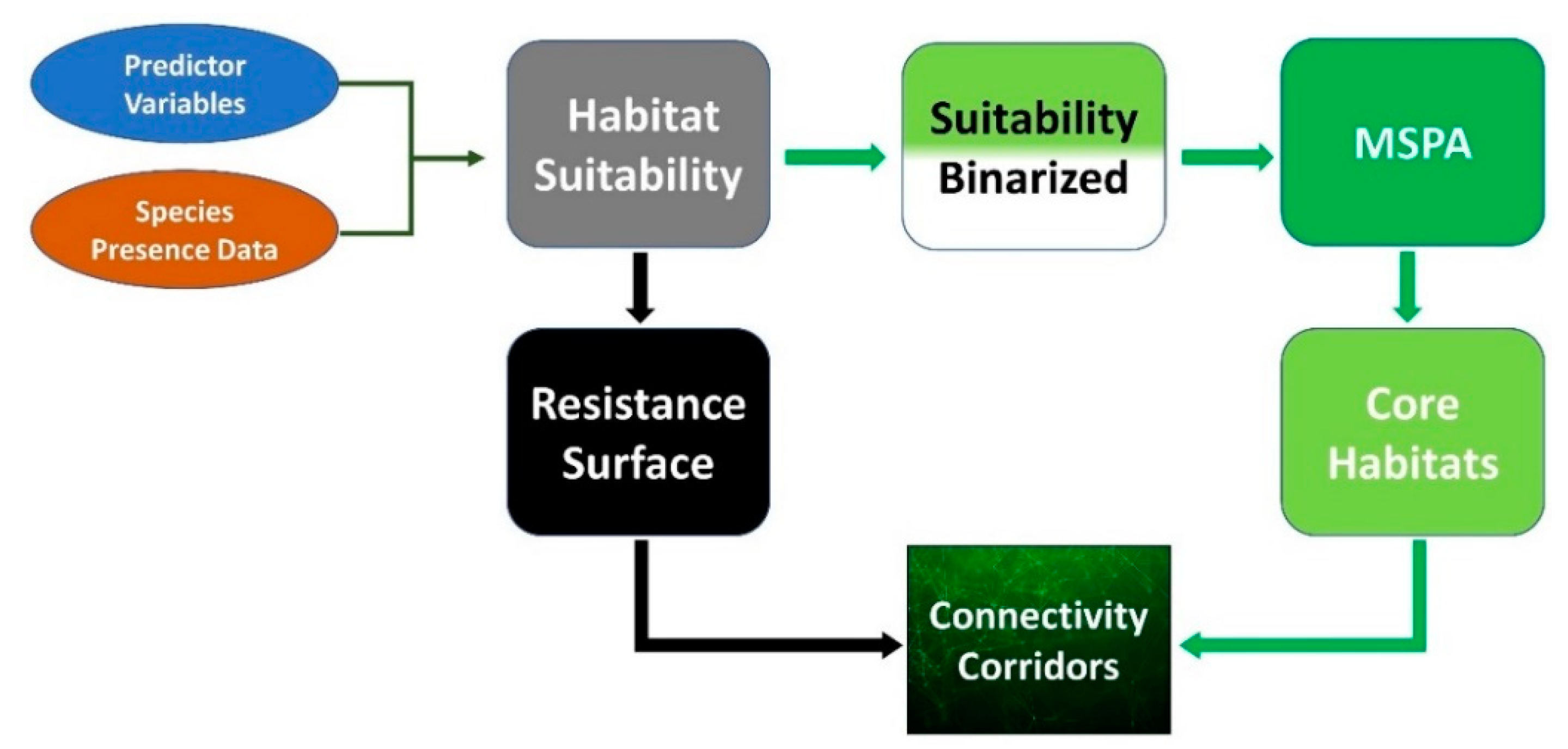
4. Materials and Methods
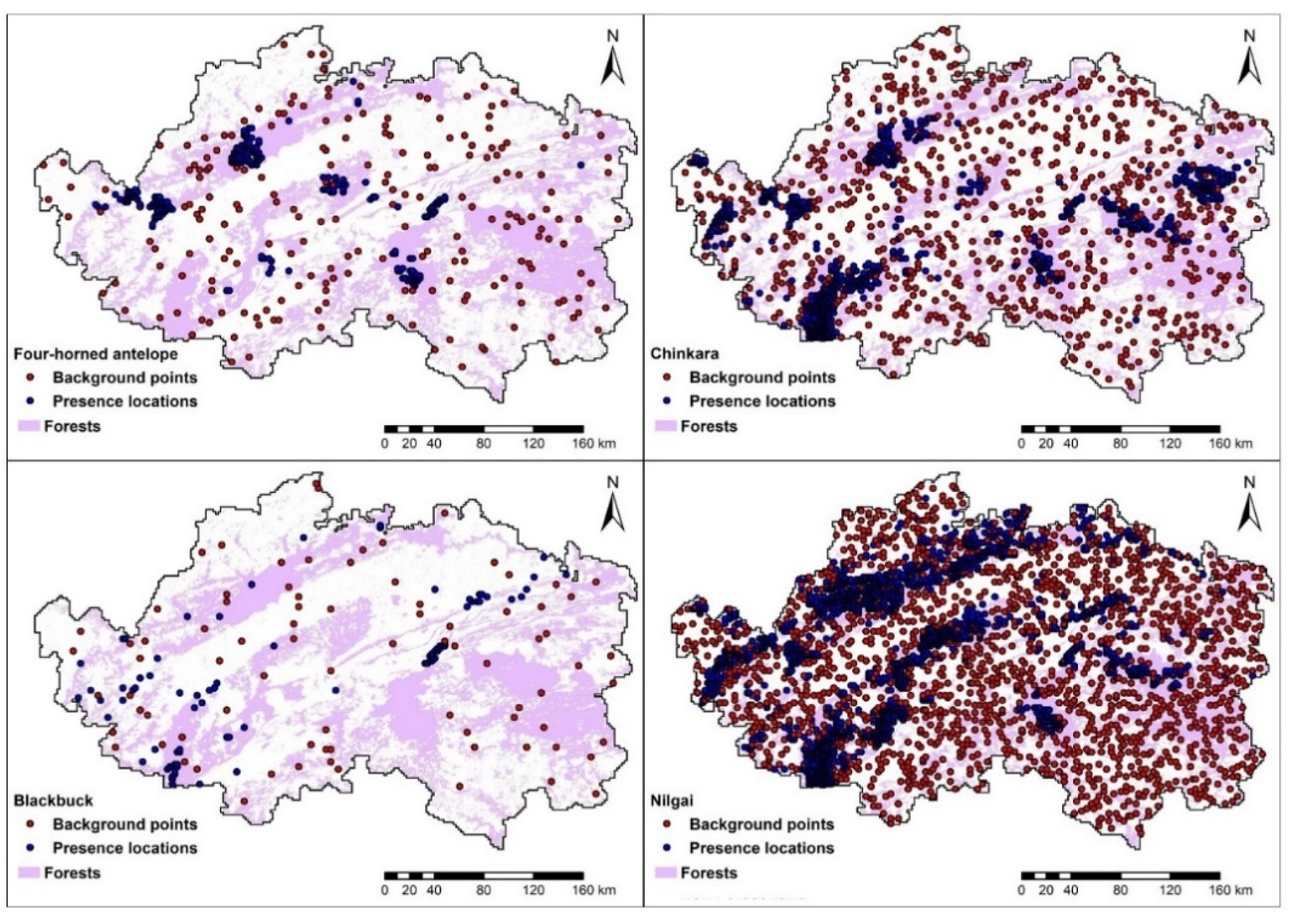
4.1. Ungulate Species Presence Data
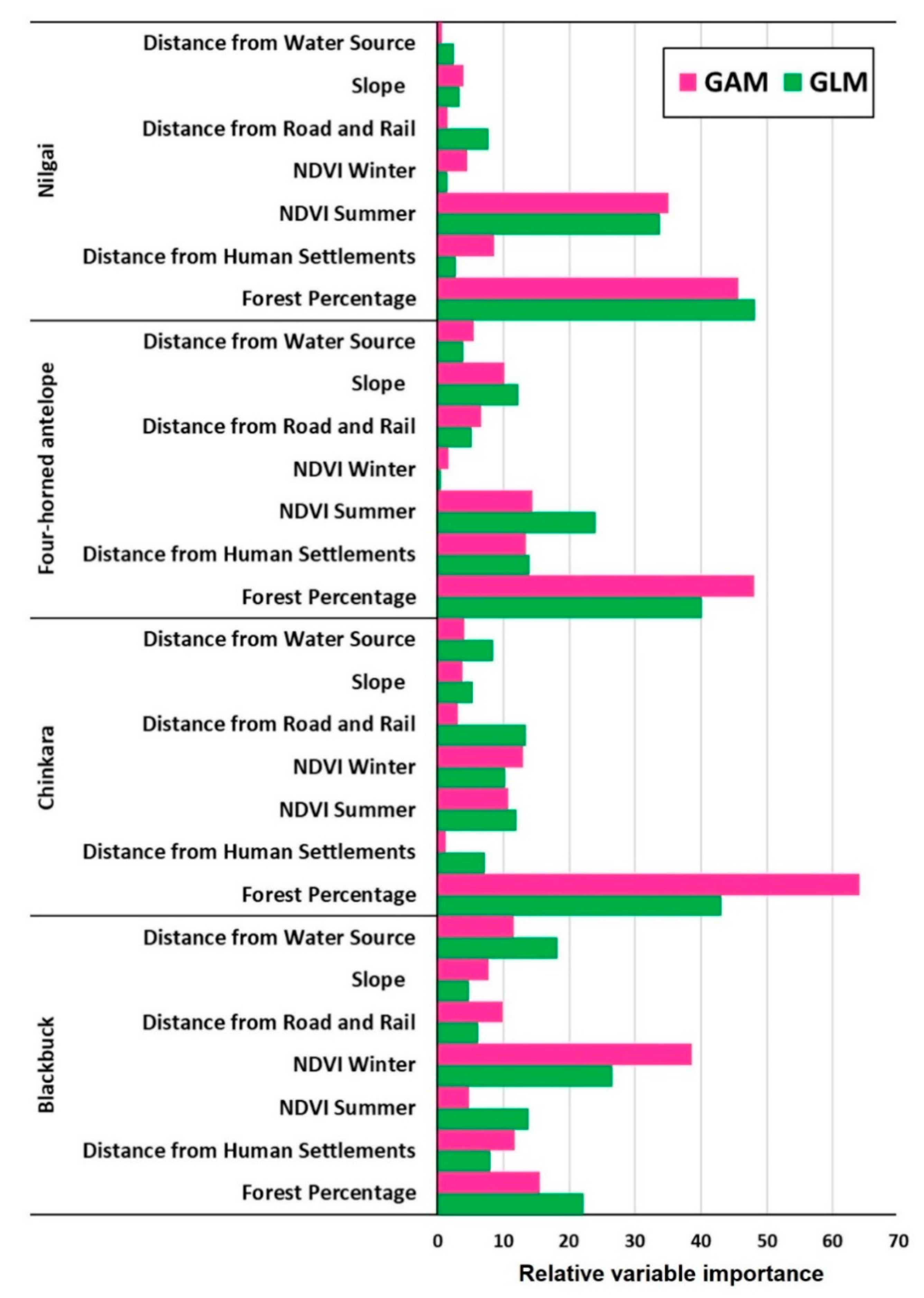
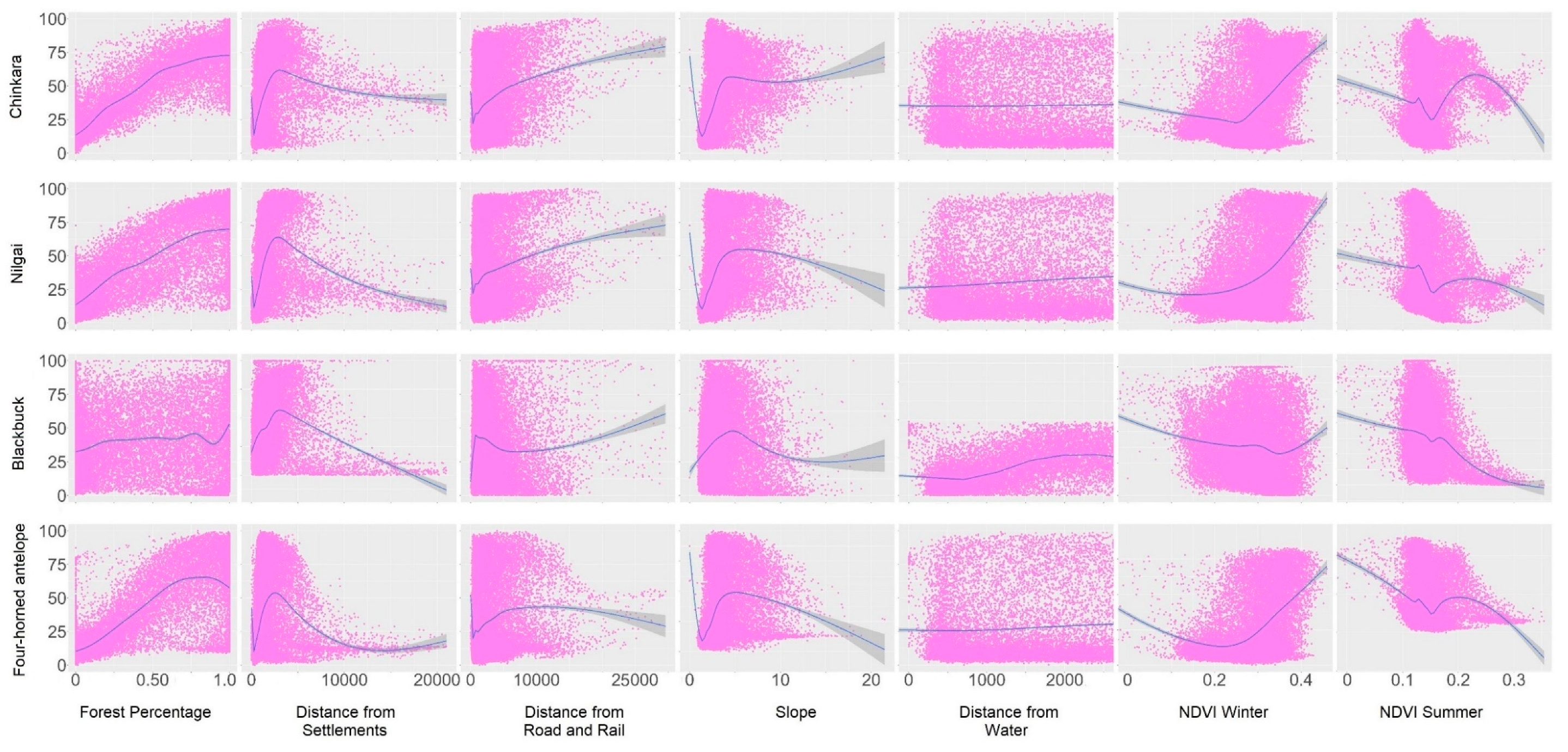
4.2. Modelling the Suitable Habitats
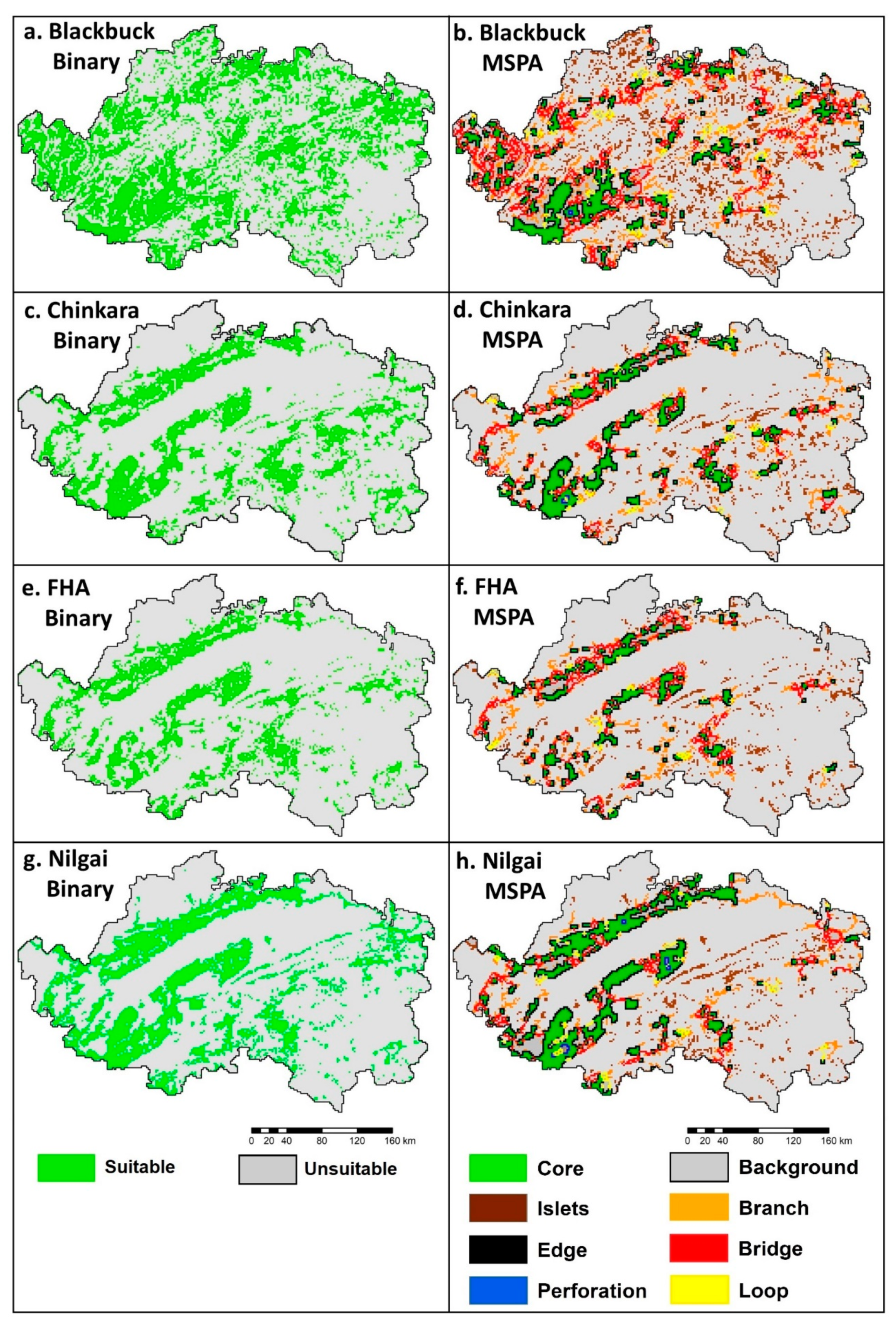
4.3. Map Binarization and Morphological Spatial Pattern Analysis (MSPA)
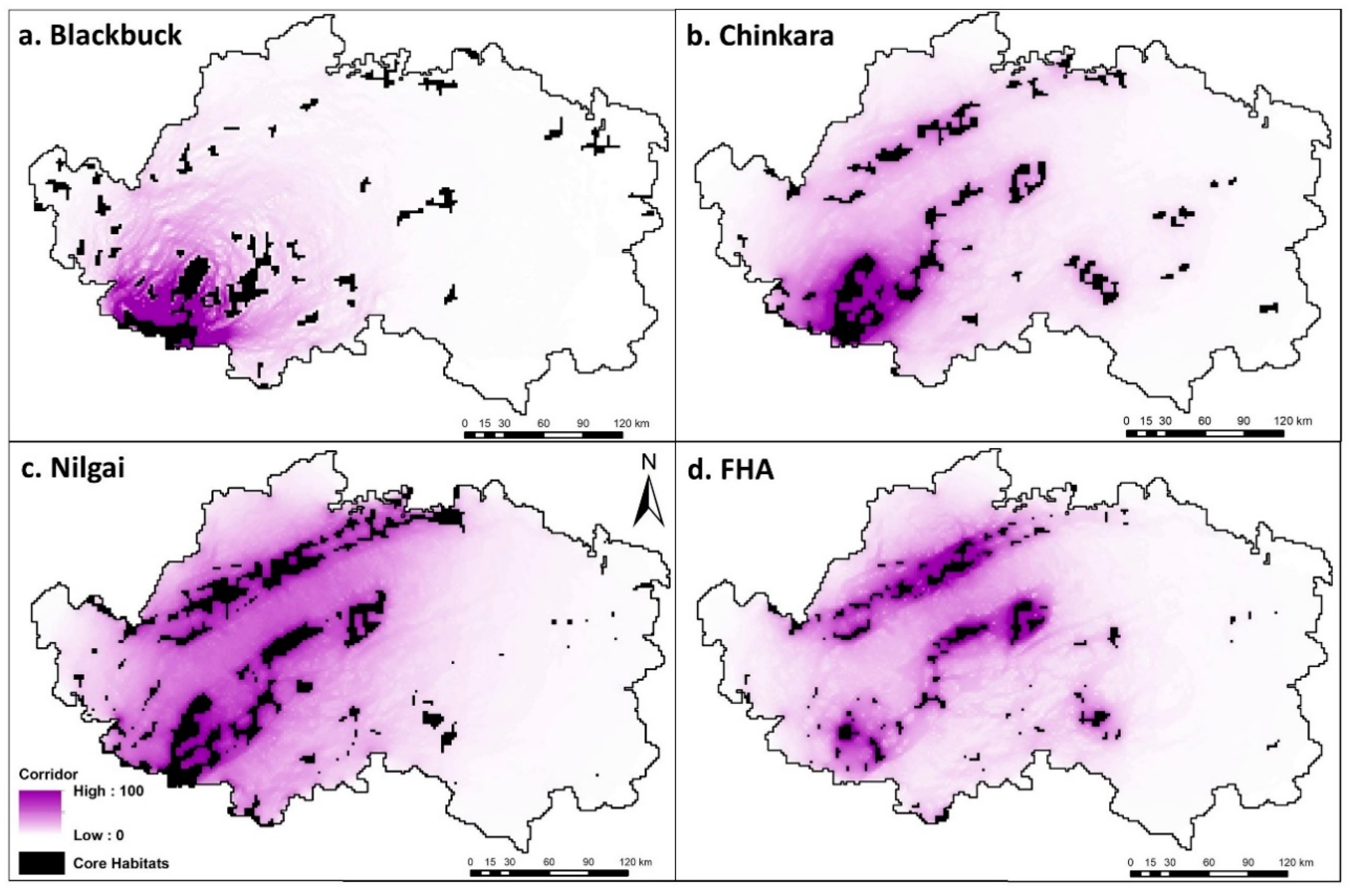
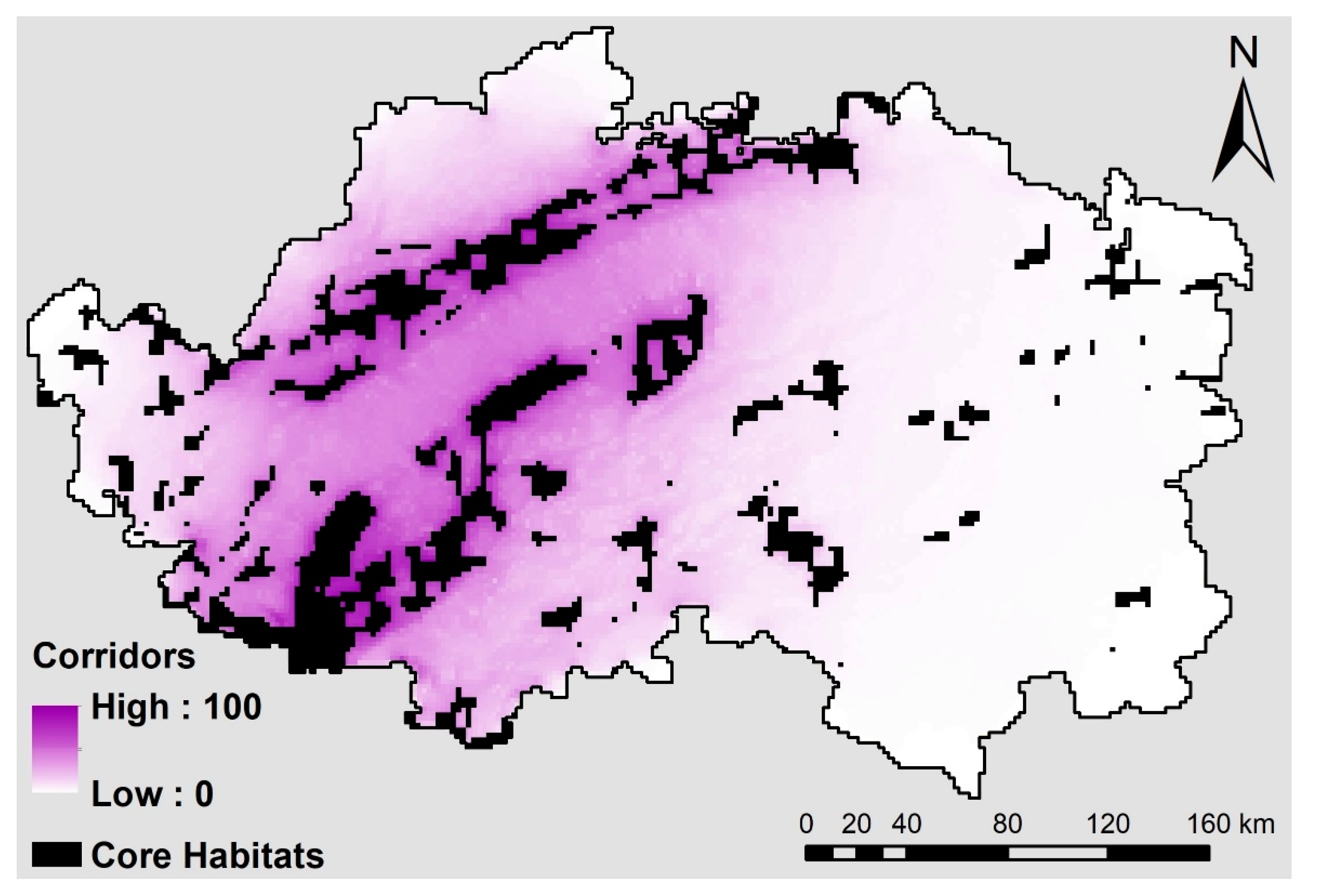
4.4. Modeling Dispersal Corridors
4.5. Quantifying Functional Connectivity in the Landscape
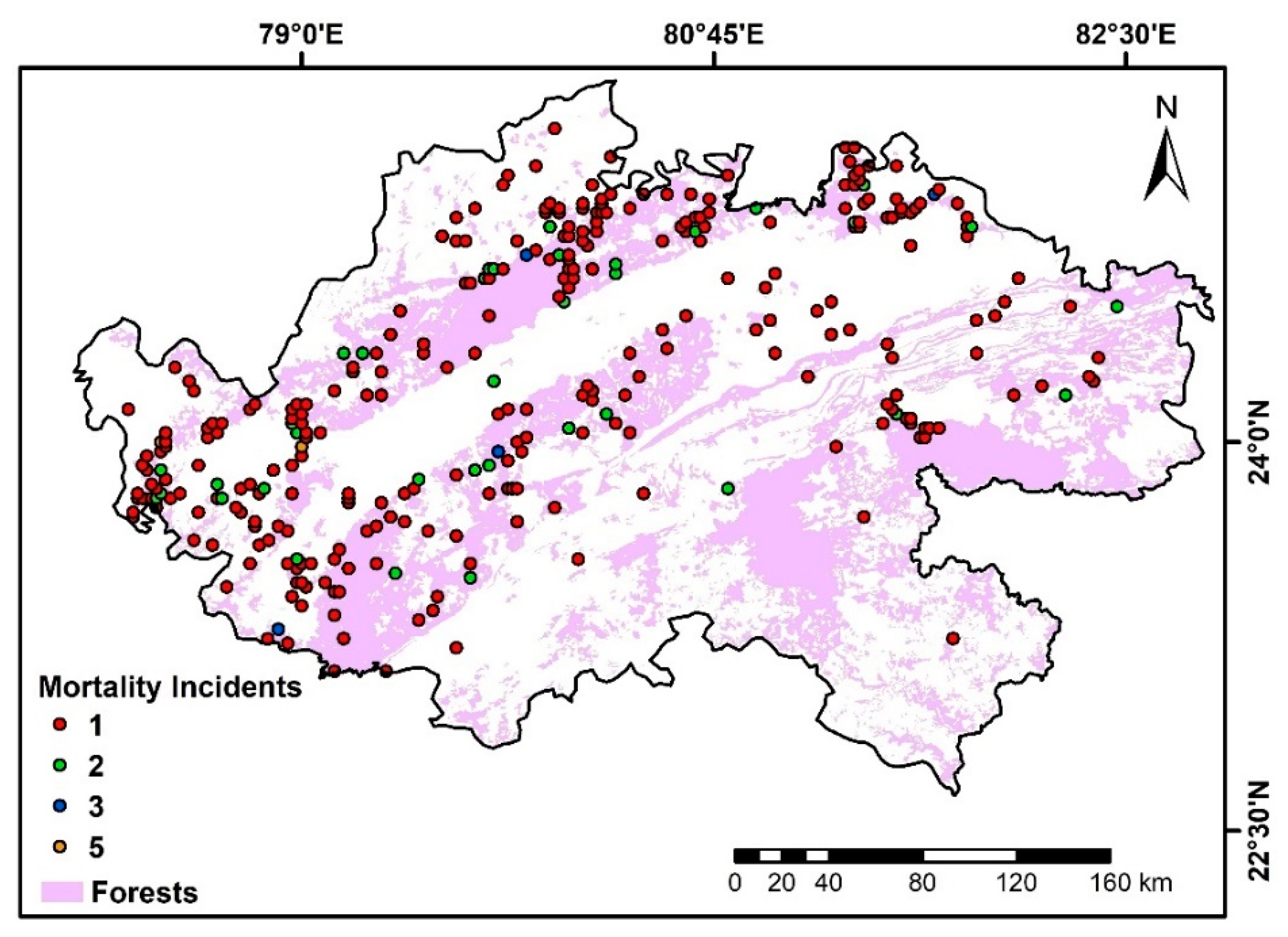
4.6. Modeling Human-Influenced Antelope Mortality
5. Results
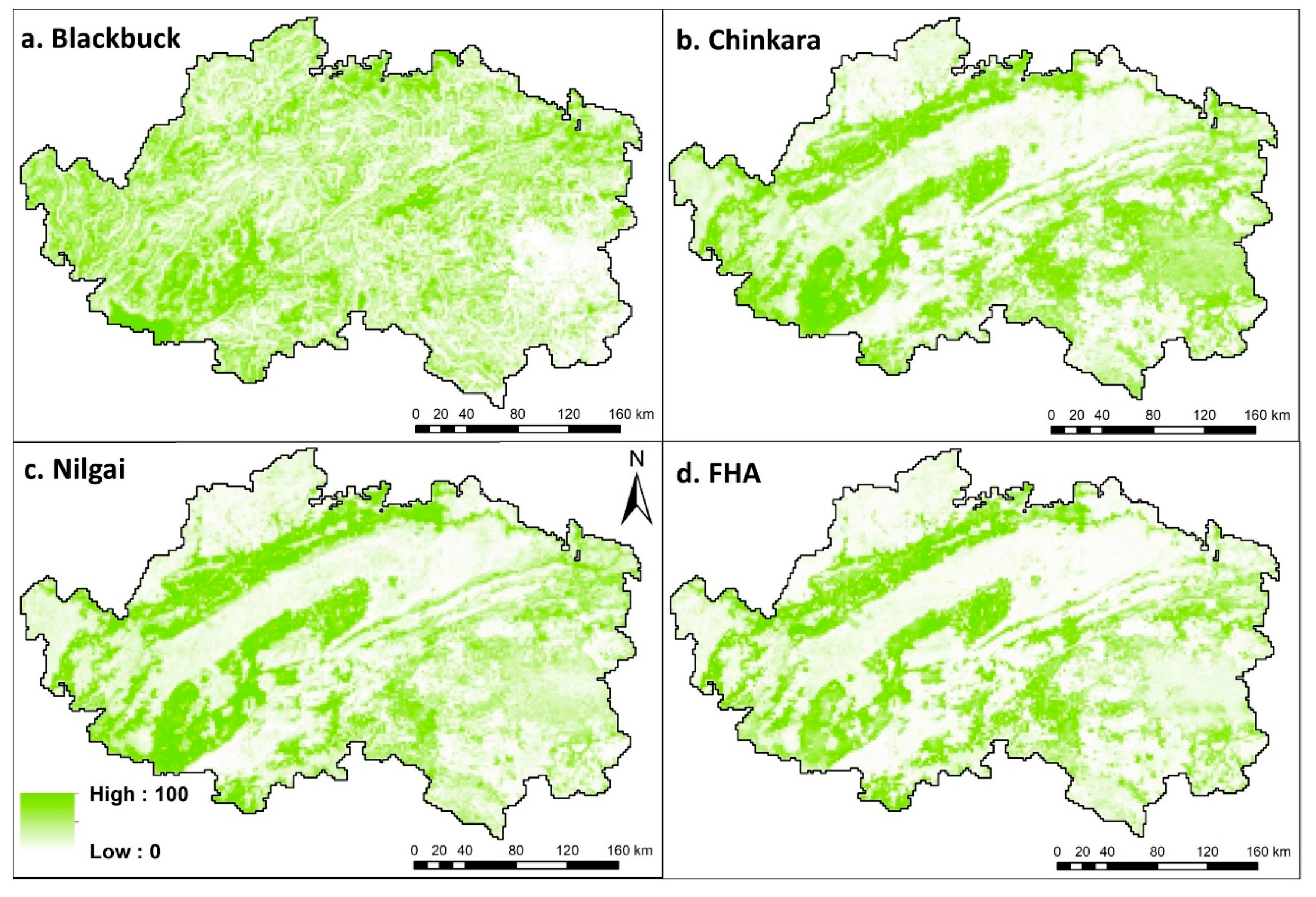
5.1. Modelling the Suitable Habitats
5.2. Map Binarization and Morphological Spatial Pattern Analysis (MSPA)
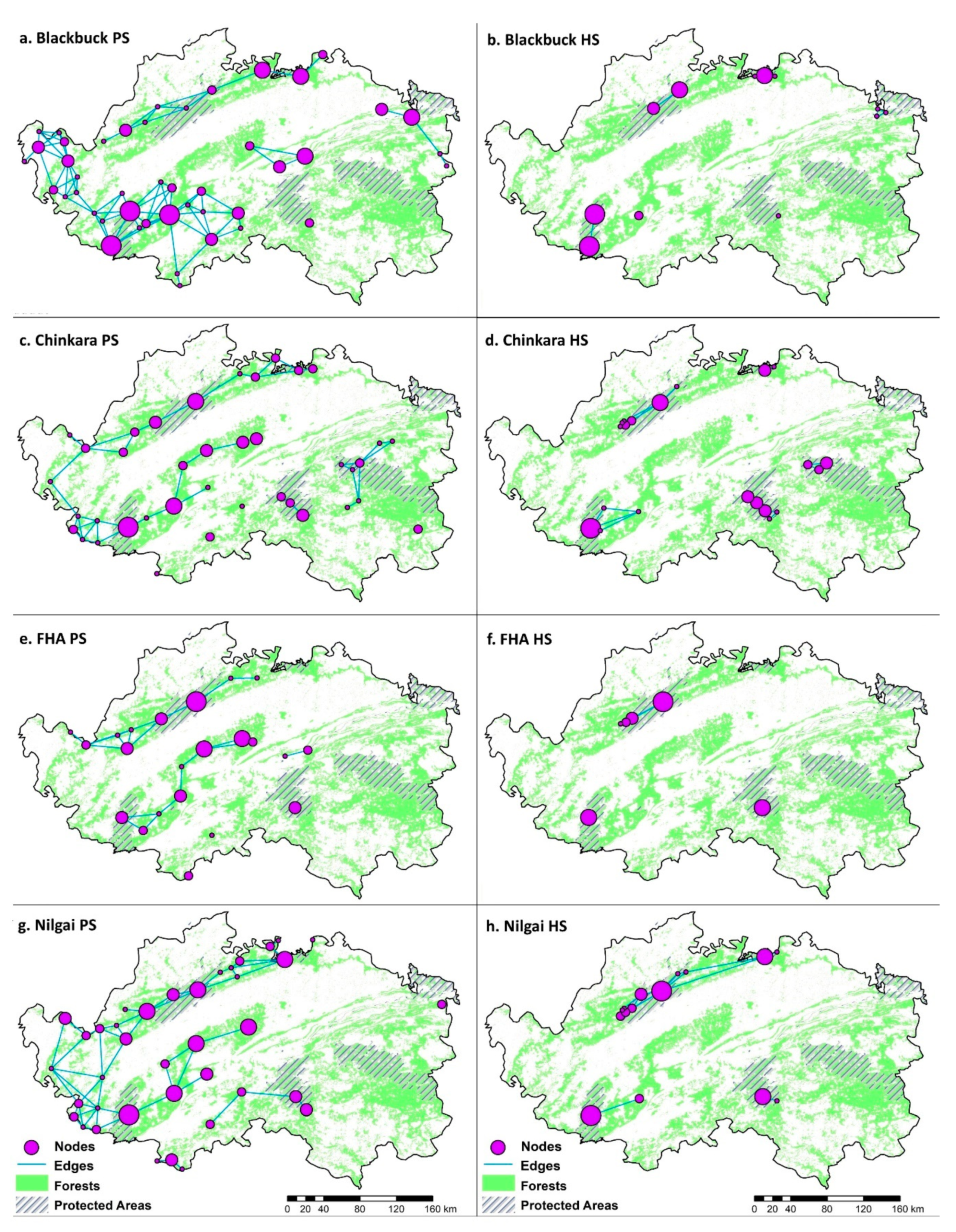
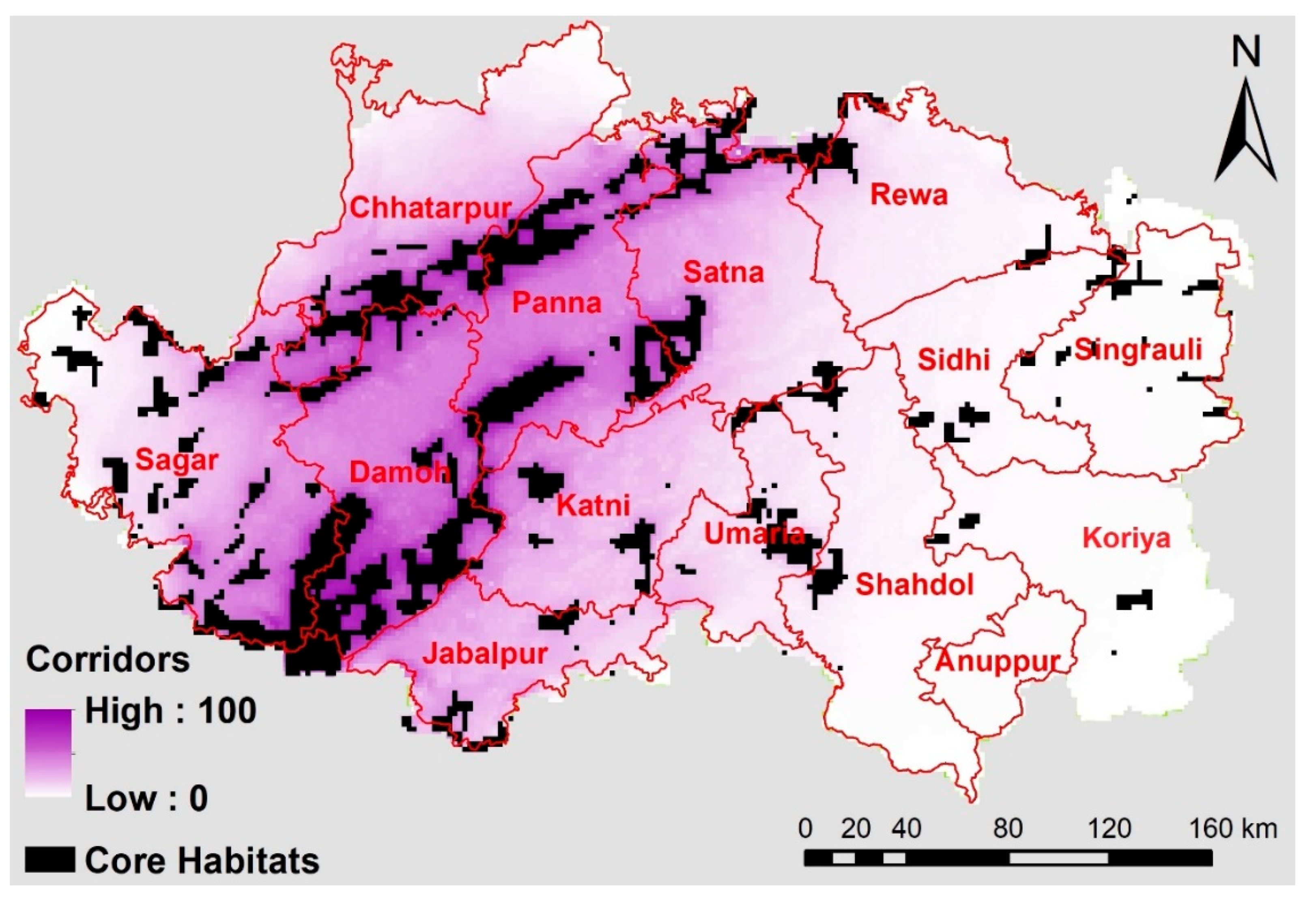
5.3. Modeling Dispersal Corridors
5.4. Quantifying Potential Functional Connectivity
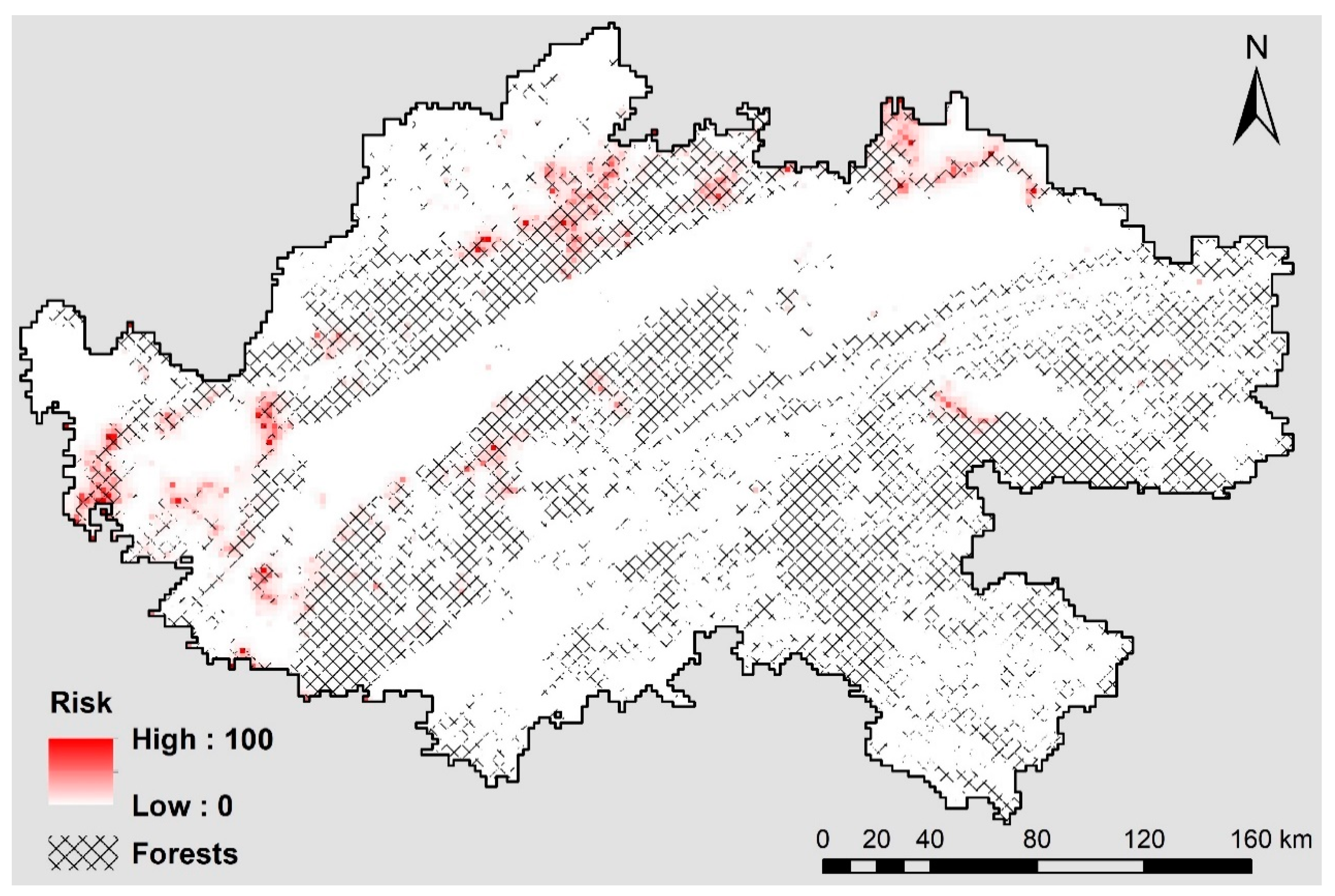
5.5. Modeling Human-Influenced Antelope Mortality
6. Discussion
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blackbuck | Chinkara | FHA | Nilgai | |||||
|---|---|---|---|---|---|---|---|---|
| GLM | GAM | GLM | GAM | GLM | GAM | GLM | GAM | |
| Forest percentage | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ |
| Pre-monsoon NDVI | ✗ | ✗ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ |
| Post-monsoon NDVI | ✓ | ✓ | ✓ | ✓ | ✗ | ✗ | ✗ | ✓ |
| Slope | ✗ | ✗ | ✓ | ✓ | ✓ | ✓ | ✗ | ✓ |
| Dist. from roads & rails | ✗ | ✗ | ✓ | ✓ | ✗ | ✓ | ✓ | ✓ |
| Dist. from water source | ✓ | ✗ | ✓ | ✓ | ✗ | ✓ | ✗ | ✗ |
| Dist. from settlements | ✓ | ✗ | ✓ | ✗ | ✓ | ✓ | ✗ | ✓ |
| Median | 2.50% | 97.50% | n.Effective | Geweke.Diag | |
|---|---|---|---|---|---|
| (Intercept) | −6.1251 | −6.5779 | −5.553 | 114.7 | 3.3 |
| dhustmts | 0.0996 | −0.0845 | 0.2827 | 1802.7 | −1.1 |
| drora | −0.2505 | −0.4468 | −0.0644 | 1291.6 | 2.7 |
| distancefb | −0.5859 | −0.8221 | −0.3579 | 2009.2 | 1.7 |
| population | 0.0102 | −0.1534 | 0.1525 | 3645.7 | −0.6 |
| forestp | 0.3725 | 0.2266 | 0.5195 | 3531 | 0 |
| Omega—(Intercept) | −181.765 | −634.328 | −1.0285 | 234.6 | 140.1 |
| tau2 | 10.3691 | 7.6972 | 13.742 | 179 | −2.2 |
| rho | 0.9984 | 0.9939 | 0.9998 | 1221.1 | 0.4 |
| DIC = 6310.09 | |||||
| p.d = 2141.987 | |||||
| LMPL = −3762.73 |

References
- Baguette, M.; Blanchet, S.; Legrand, D.; Stevens, V.M.; Turlure, C. Individual dispersal, landscape connectivity and ecological networks: Dispersal, connectivity and networks. Biol. Rev. 2013, 88, 310–326. [Google Scholar] [CrossRef]
- Taylor, P.D.; Fahrig, L.; Henein, K.; Merriam, G. Connectivity is a vital element of landscape structure. Oikos 1993, 68, 571–573. [Google Scholar] [CrossRef] [Green Version]
- Churko, G.; Kienast, F.; Bolliger, J.A. Multi-species assessment to identify the functional connectivity of amphibians in a human-dominated landscape. Int. J. Geo-Inf. 2020, 9, 287. [Google Scholar] [CrossRef]
- Kashwan, P. Power asymmetries and institutions: Landscape conservation in central India. Reg. Environ. Chang. 2016, 16, 97–109. [Google Scholar] [CrossRef]
- Trombulak, S.C.; Baldwin, R.F. Introduction: Creating a context for landscape-Scale conservation planning. In Landscape-Scale Conservation Planning; Trombulak, S.C., Baldwin, R.F., Eds.; Springer: New York, NY, USA, 2010; pp. 1–15. ISBN 978-90-481-9575-6. [Google Scholar]
- Hanski, I. Metapopulation Ecology; Oxford University Press: New York, NY, USA, 1999; ISBN 0-19-854065-5. [Google Scholar]
- Hanski, I. Spatially realistic theory of metapopulation ecology. Naturwissenschaften 2001, 88, 372–381. [Google Scholar] [CrossRef]
- Williams, S.T.; Williams, K.S.; Lewis, B.P.; Hill, R.A. Population dynamics and threats to an apex predator outside protected areas: Implications for carnivore management. R. Soc. Open Sci. 2017, 4, 161090. [Google Scholar] [CrossRef] [Green Version]
- Devictor, V.; Godet, L.; Julliard, R.; Couvet, D.; Jiguet, F. Can common species benefit from protected areas? Biol. Conserv. 2007, 139, 29–36. [Google Scholar] [CrossRef]
- Lopoukhine, N.; Crawhall, N.; Dudley, N.; Figgis, P.; Karibuhoye, C.; Laffoley, D.; Londoño, J.M.; MacKinnon, K.; Sandwith, T. Protected areas: Providing natural solutions to 21st century challenges. SAPI EN. S. Surv. Perspect. Integr. Environ. Soc. 2012, 5, 117–131. [Google Scholar]
- Hughes, J.; Macdonald, D.W. A review of the interactions between free-Roaming domestic dogs and wildlife. Biol Conserv. 2013, 157, 341–351. [Google Scholar] [CrossRef]
- Hussain, S.; Ram, M.S.; Kumar, A.; Shivaji, S.; Umapathy, G. Human presence increases parasitic load in endangered lion-tailed macaques (macaca silenus) in its fragmented rainforest habitats in Southern India. PLoS ONE 2013, 8, e63685. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, D.; Reddy, M.; Tiwari, S.; Umapathy, G. Land use change increases wildlife parasite diversity in Anamalai Hills, Western Ghats, India. Sci. Rep. 2019, 9, 11975. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, A.; Kumar, V.; Kittur, S.; Reddy, M.; Naidenko, S.; Ganswindt, A.; Umapathy, G. Physiological stress responses of tigers due to anthropogenic disturbance especially tourism in two Central Indian tiger reserves. Conserv. Physiol. 2019, 7, coz045. [Google Scholar] [CrossRef]
- Naidenko, S.V.; Berezhnoi, M.A.; Kumar, V.; Umapathy, G. Comparison of tigers’ fecal glucocorticoids level in two extreme habitats. PLoS ONE 2019, 14, e0214447. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Pradheeps, M.; Kokkiligadda, A.; Niyogi, R.; Umapathy, G. Non-invasive assessment of physiological stress in captive Asian elephants. Animals 2019, 9, 553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasgupta, S.; Ghosh, A.K. Elephant–Railway conflict in a biodiversity hotspot: Determinants and perceptions of the conflict in Northern West Bengal, India. Hum. Dimens. Wildl. 2015, 20, 81–94. [Google Scholar] [CrossRef]
- Nezval, V.; Bíl, M. Spatial analysis of wildlife-train collisions on the Czech rail network. Appl. Geogr. 2020, 125, 102304. [Google Scholar] [CrossRef]
- Sánchez-Mercado, A.; Ferrer-Paris, J.R.; García-Rangel, S.; Yerena, E.; Robertson, B.A.; Rodríguez-Clark, K.M. Combining threat and occurrence models to predict potential ecological traps for Andean bears in the Cordillera de Mérida, Venezuela: Predicting potential ecological traps. Anim. Conserv. 2014, 17, 388–398. [Google Scholar] [CrossRef]
- Kramer-Schadt, S.; Revilla, E.; Wiegand, T.; Breitenmoser, U. Fragmented landscapes, road mortality and patch connectivity: Modelling influences on the dispersal of Eurasian lynx. J. Appl. Ecol. 2004, 41, 711–723. [Google Scholar] [CrossRef] [Green Version]
- Loveridge, A.J.; Valeix, M.; Elliot, N.B.; Macdonald, D.W. The landscape of anthropogenic mortality: How African lions respond to spatial variation in risk. J. Appl. Ecol. 2017, 54, 815–825. [Google Scholar] [CrossRef]
- Lehman, R.N. Raptor electrocution on power lines: Current issues and outlook. Wildl. Soc. Bull. 2001, 29, 804–813. [Google Scholar]
- Haas, D. Protecting Birds from Powerlines: Convention on the Conservation of European Wildlife and Habitats (Bern Convention); Council of Europe: Strasbourg, France, 2005; Volume 140, ISBN 92-871-5630-1. [Google Scholar]
- Ehlers Smith, Y.C.; Ehlers Smith, D.A.; Ramesh, T.; Downs, C.T. Novel predators and anthropogenic disturbance influence spatio-temporal distribution of forest antelope species. Behav. Process. 2019, 159, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Vanak, A.T.; Gompper, M.E. Dogs canis familiaris as carnivores: Their role and function in intraguild competition. Mammal Rev. 2009, 39, 265–283. [Google Scholar] [CrossRef]
- Reinhardt, K.; Köhler, G.; Maas, S.; Detzel, P. Low dispersal ability and habitat specificity promote extinctions in rare but not in widespread species: The orthoptera of Germany. Ecography 2005, 28, 593–602. [Google Scholar] [CrossRef]
- Poethke, H.J.; Hovestadt, T.; Mitesser, O. Local extinction and the evolution of dispersal rates: Causes and correlations. Am. Nat. 2003, 161, 631–640. [Google Scholar] [CrossRef] [Green Version]
- Brennan, A.; Beytell, P.; Aschenborn, O.; Du Preez, P.; Funston, P.J.; Hanssen, L.; Kilian, J.W.; Stuart-Hill, G.; Taylor, R.D.; Naidoo, R. Characterizing multispecies connectivity across a transfrontier conservation landscape. J. Appl. Ecol. 2020, 57, 1700–1710. [Google Scholar] [CrossRef]
- Sarkar, M.S.; Krishnamurthy, R.; Johnson, J.A.; Sen, S.; Nigam, P.; Gupta, S.; Murthy, R.; Saha, G. Movement and home range characteristics of reintroduced tiger (Panthera tigris) population in Panna tiger reserve, Central India. Eur. J. Wildl. Res. 2016, 62, 537–547. [Google Scholar] [CrossRef]
- Sarkar, M.S.; Niyogi, R.; Hazra, P.; John, R. Panna Landscape Corridor Connections: Fine Scale Assessment of Tiger (Panthera tigris) Connectivity Networks in the Panna Landscape, Madhya Pradesh; Madhya Pradesh State Biodiversity Board: Bhopal, India, 2019; ISBN 978-81-941948-1-1. [Google Scholar]
- Sarkar, M.S.; Krishnamurthy, R.; Johnson, J.A.; Sen, S.; Saha, G.K. Assessment of fine-scale resource selection and spatially explicit habitat suitability modelling for a re-introduced tiger (Panthera tigris) population in Central India. PeerJ 2017, 5, e3920. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.A.; Cushman, S.A.; Srivastava, A.; Sarkar, M.S.; Shivaji, S. Tiger abundance and gene flow in Central India are driven by Disparate combinations of topography and land cover. Divers. Distrib. 2017, 23, 863–874. [Google Scholar] [CrossRef]
- Li, C.; Jiang, Z.; Fang, H.; Li, C.A. Spatially explicit model of functional connectivity for the endangered Przewalski’s gazelle (Procapra przewalskii) in a patchy landscape. PLoS ONE 2013, 8, e80065. [Google Scholar] [CrossRef] [Green Version]
- Zhuge, H.; Lin, D.; Li, X. Identification of ecological corridors for tibetan antelope and assessment of their human disturbances in the Alpine desert of Qinghai-Tibet Plateau. J. Appl. Ecol. 2015, 26, 2504. [Google Scholar]
- Berger, J.; Young, J.K.; Berger, K.M. Protecting migration corridors: Challenges and optimism for Mongolian saiga. PLoS Biol. 2008, 6, e165. [Google Scholar] [CrossRef] [PubMed]
- Malakoutikhah, S.; Fakheran, S.; Hemami, M.; Tarkesh, M.; Senn, J. Assessing future distribution, suitability of corridors and efficiency of protected areas to conserve vulnerable ungulates under climate change. Divers. Distrib. 2020, 26, 1383–1396. [Google Scholar] [CrossRef]
- Esmaeili, M.; Shayesteh, K.; Karami, P. Investigation of Habitat Suitability and Connectivity Pathways of Persian Gazelle (Gazella Subgutturusa Subgutturosa) in the West of Kermanshah Province and East of Iraq (Case Study: Qaraviz No-Hunting Area). J. Anim. Environ. 2020, 12, 23–30. [Google Scholar]
- Khosravi, R.; Hemami, M.R.; Malekian, M.; Silva, T.L.; Rezaei, H.R.; Brito, J.C. Effect of landscape features on genetic structure of the Goitered Gazelle (Gazella subgutturosa) in Central Iran. Conserv. Genet. 2018, 19, 323–336. [Google Scholar] [CrossRef]
- Yang, J.; Jiang, Z.; Zeng, Y.; Turghan, M.; Fang, H.; Li, C. Effect of anthropogenic landscape features on population genetic differentiation of Przewalski’s Gazelle: Main role of human settlement. PLoS ONE 2011, 6, e20144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnamurthy, G.; Rajput, A.; Majumder, A.; Ali, Z. Status Report: Tigers, Co-Predators and Prey in Protected Areas of Madhya Pradesh; Technical Report No. QMSP-6.3A-04; State Forest Research Institute: Jabalpur, India, 2016. [Google Scholar]
- Sankaran, M.; Augustine, D.J.; Ratnam, J. Native ungulates of diverse body sizes collectively regulate long-term woody plant demography and structure of a Semi-Arid Savanna. J. Ecol. 2013, 101, 1389–1399. [Google Scholar] [CrossRef]
- Gadgil, M.; Meher-Homji, V. Ecological diversity. In Conservation in Developing Countries: Problems and Prospects; Daniel, J.C., Serrao, J.S., Eds.; Bombay Natural History Society: Bombay, India, 1990; pp. 175–198. [Google Scholar]
- MoEF. Indian Wildlife (Protection) Act; MoEF: New Delhi, India, 1972.
- Sabogal, C.; Guariguata, M.; Broadhead, J.; Lescuyer, G.; Savilaakso, S.; Essoungou, J.N.; Sist, P. Multiple-Use Forest Management in the Humid Tropics: Opportunities and Challenges for Sustainable Forest Management; FAO: Rome, Italy, 2013. [Google Scholar]
- The Concept of Multiple Use of Forest and Associated Lands – Its Values and Limitations. Available online: http://www.fao.org/3/X5397E/x5397e03.htm (accessed on 15 December 2020).
- Krishnan, P.; Ramakrishnan, R.; Saigal, S.; Nagar, S.; Faizi, S.; Panwar, H.; Singh, S.; Ved, N. Conservation Across Landscapes: India’s Approaches to Biodiversity Governance; United Nations Development Programme: New Delhi, India, 2012; pp. 1–186. [Google Scholar]
- Chhangani, A.K.; Robbins, P.; Mohnot, S.M. Crop raiding and livestock predation at Kumbhalgarh wildlife sanctuary, Rajasthan India. Hum. Dimens. Wildl. 2008, 13, 305–316. [Google Scholar] [CrossRef]
- Leslie, D.M. Boselaphus tragocamelus (Artiodactyla: Bovidae). Mamm. Species 2008, 813, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, N.; Sawarkar, V. Problems of over-abundant populations of ’Nilgai’ and ’Blackbuck’ in Haryana and Madhya Pradesh and their management. Indian For. 1989, 115, 488–493. [Google Scholar]
- Chauhan, N.; Singh, R. Crop damage by overabundant populations of Nilgai and Blackbuck in Haryana (India) and its management. In Proceedings of the Fourteenth Vertebrate Pest Conference, Sacramento, CA, USA, 6–8 March 1990. [Google Scholar]
- Goyal, S.K.; Rajpurohit, L.S. Nilgai, Boselaphus tragocamelus-a mammalian crop pest around Jodhpur. Uttar Pradesh J. Zool. 2000, 20, 55–59. [Google Scholar]
- Jhala, Y.V. Damage to sorghum crop by blackbuck. Int. J. Pest Manag. 1993, 39, 23–27. [Google Scholar] [CrossRef]
- Sankar, K.; Goyal, S. Ungulates of India; WII-ENVIS Centre on ”Wildlife and Protected Areas”: Dehradun, India, 2004. [Google Scholar]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Naimi, B.; Araújo, M.B. Sdm: A reproducible and extensible r platform for species distribution modelling. Ecography 2016, 39, 368–37556. [Google Scholar] [CrossRef] [Green Version]
- Madhya Pradesh Forest Department Official Website. Available online: https://mpforest.gov.in/ (accessed on 18 December 2020).
- Zurell, D.; Franklin, J.; König, C.; Bouchet, P.J.; Dormann, C.F.; Elith, J.; Fandos, G.; Feng, X.; Guillera-Arroita, G.; Guisan, A.; et al. A standard protocol for reporting species distribution models. Ecography 2020, 43, 1261–1277. [Google Scholar] [CrossRef]
- Species Distribution Modeling—R Spatial. Available online: https://rspatial.org/raster/sdm/ (accessed on 15 December 2020).
- Swets, J. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OpenStreetMapData. Available online: http://ww12.openstreetmapdata.com/ (accessed on 15 December 2020).
- Ormsby, T.; Napoleon, E.; Burke, R.; Groessl, C.; Bowden, L. Getting to Know ArcGIS Desktop; ESRI Press: Redlands, CA, USA, 2010; ISBN 1-58948-260-3. [Google Scholar]
- LP DAAC–Homepage. Available online: https://lpdaac.usgs.gov/ (accessed on 15 December 2020).
- Jenness, J. DEM Surface Tools; Jenness Enterprises: Flagstaff, AZ, USA, 2013. [Google Scholar]
- Earth Resources Observation and Science (EROS). Earth Resources Observation and Science (EROS) Center Collection-2 Landsat 8-9 OLI (Operational Land Imager) and TIRS (Thermal Infrared Sensor) Level-1 Data Products; USGS: Reston, VA, USA, 2013.
- Freeman, E. Package ‘PresenceAbsence’; R Package Version 1.1.9; 2012; Available online: https://cran.r-project.org/package=PresenceAbsence (accessed on 7 June 2020).
- Soille, P.; Vogt, P. Morphological segmentation of binary patterns. Pattern Recognit. Lett. 2009, 30, 456–459. [Google Scholar] [CrossRef]
- Vogt, P.; Riitters, K. Guidostoolbox: Universal digital image object analysis. Eur. J. Remote. Sens. 2017, 50, 352–361. [Google Scholar] [CrossRef]
- Vogt, P.; Riitters, K.H.; Estreguil, C.; Kozak, J.; Wade, T.G.; Wickham, J.D. Mapping spatial patterns with morphological image processing. Landsc. Ecol. 2007, 22, 171–177. [Google Scholar] [CrossRef]
- Vogt, P.; Riitters, K.H.; Iwanowski, M.; Estreguil, C.; Kozak, J.; Soille, P. Mapping landscape corridors. Ecol. Indic. 2007, 7, 481–488. [Google Scholar] [CrossRef]
- Zeller, K.A.; Jennings, M.K.; Vickers, T.W.; Ernest, H.B.; Cushman, S.A.; Boyce, W.M. Are all data types and connectivity models created equal? Validating common connectivity approaches with dispersal data. Divers Distrib. 2018, 24, 868–879. [Google Scholar] [CrossRef] [Green Version]
- McRae, B.H.; Beier, P. Circuit theory predicts gene flow in plant and animal populations. Proc. Natl. Acad. Sci. USA 2007, 104, 19885–19890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Etten, J. R package gdistance: Distances and routes on geographical grids. J. Stat. Softw. 2017, 76, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Trainor, A.M.; Walters, J.R.; Morris, W.F.; Sexton, J.; Moody, A. Empirical estimation of dispersal resistance surfaces: A case study with red-cockaded woodpeckers. Landsc. Ecol. 2013, 28, 755–767. [Google Scholar] [CrossRef]
- Keeley, A.T.H.; Beier, P.; Gagnon, J.W. Estimating landscape resistance from habitat suitability: Effects of data source and nonlinearities. Landsc. Ecol. 2016, 31, 2151–2162. [Google Scholar] [CrossRef]
- Santini, L.; Di Marco, M.; Visconti, P.; Baisero, D.; Boitani, L.; Rondinini, C. Ecological correlates of dispersal distance in terrestrial mammals. Hystrix Ital. J. Mammal. 2013, 24. [Google Scholar] [CrossRef]
- Ranjitsinh, M. Indian Blackbuck; Natraj Publishers: Dehradun, India, 1989; ISBN 81-85019-18-5. [Google Scholar]
- Kumar, A. Niraj distribution pattern of Black Buck (Antelope Cervicapra) at Gajner wild life sanctuary, Bikaner (Rajasthan). Int. J. Sci. Res. Rev. 2016, 5, 26–34. [Google Scholar]
- Prasad, N.L.N.S. Home Range Size of Blackbuck, Antilope Cervicapra, at Mudmal. Zeitschrift Fuer Saeugetierkunde 1983, 48, 109–117. [Google Scholar]
- Mahato, A.K.R.; Raziuddin, M. Status, Ecology & Behaviour of Antilope Cervicapra (Linnaeus, 1758) in Proposed Community Reserve for Blackbuck, Ganjam District, Orissa, India; Zoological Survey of India: Kolkata, India, 2010; ISBN 81-8171-265-X. [Google Scholar]
- Khanal, P.; Khanal, S.; Jnawali, S.; Pathak, S. Study on the land use of proposed Blackbuck conservation area, Khairapur, Bardia district and habitat options for translocation of Blackbuck (Antelope cervicapra) at Royal Suklaphanta Wildlife Reserve in Kanchanpur district, Nepal. Local Gov. Programme United Nations Dev. Programme 2002. [Google Scholar] [CrossRef]
- Sankar, K. The Ecology of Three Large Sympatric Herbivores (Chital, Sambar, Nilgai) with Special Reference for Reserve Management in Sariska Tiger Reserve, Rajasthan. Ph.D. Thesis, University of Rajasthan, Jaipur, India, 1994. [Google Scholar]
- Dookia, S. Habitat Preference, Abundance and Group Size of Indian Gazelle (Gazella Bennetti Sykes, 1831) in Semi Arid Region of Rajasthan. Ph.D. Thesis, J.N.V. University, Jodhpur, Rajasthan, 2002. [Google Scholar]
- Jaipal, B. Home range and territorial activities of Chinkara (Gazella bennetti) in the Thar landscape. J. Exp. Zool. India 2020, 23, 1431–1434. [Google Scholar]
- Vaghela, M.; Dodia, P.; Shukla, A. Study on Group Composition of Blue Bull (Boselaphus tragocamelus). Ela Found. 2020, 9, 747. [Google Scholar]
- Sharma, K.; Rahmani, A. Four horned antelope or Chowsingha (Tetracerous Quadricornis, Blainville, 1816). In Ungulates of India; ENVIS Bulletin (Wildlife and Protected Areas); Wildlife Institute of India: Dehradun, India, 2005; Volume 7, pp. 53–60. [Google Scholar]
- Foltête, J.C.; Clauzel, C.; Vuidel, G. A software tool dedicated to the modelling of landscape networks. Environ. Model. Softw. 2012, 38, 316–327. [Google Scholar] [CrossRef]
- Clauzel, C.; Foltête, J.C.; Girardet, X.; Vuidel, G. Graphab 2.4 User Manual. 2019. Available online: https://sourcesup.renater.fr/www/graphab/download/manual-2.4-en.pdf (accessed on 25 July 2020).
- Meiyappan, P.P.S.; Roy, P.S.; Soliman, A.; Li, T.; Mondal, P.; Wang, S.; Jain, A.K. India Village-Level Geospatial Socio-Economic Data Set: 1991, 2001; NASA Socioeconomic Data and Applications Center (SEDAC): Palisades, NY, USA, 2018. [Google Scholar]
- Lambert, D. Zero-inflated poisson regression, with an application to defects in manufacturing. Technometrics 1992, 34, 1–14. [Google Scholar] [CrossRef]
- Zeileis, A.; Kleiber, C.; Jackman, S. Regression models for count data in R. J. Stat. Softw. 2008, 27, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Lee, D. CARBayes: An R package for Bayesian spatial modeling with conditional autoregressive priors. J. Stat. Softw. 2013, 55, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Leroux, B.G.; Lei, X.; Breslow, N. Estimation of disease rates in small areas: A new mixed model for spatial dependence. In Statistical Models in Epidemiology, the Environment, and Clinical Trials; Springer: New York, NY, USA, 2000; pp. 179–191. [Google Scholar]
- Goswami, V.R.; Vasudev, D.; Oli, M.K. The importance of conflict-induced mortality for conservation planning in areas of Human–Elephant co-occurrence. Biol. Conserv. 2014, 176, 191–198. [Google Scholar] [CrossRef]
- Hanski, I.; Simberloff, D. The metapopulation approach, its history, conceptual domain, and application to conservation. In Metapopulation Biology; Elsevier: Amsterdam, The Netherlands, 1997; pp. 5–26. [Google Scholar]
- Bzdok, D.; Altman, N.; Krzywinski, M. Points of significance: Statistics versus machine learning. Nat. Methods 2018, 15, 233–234. [Google Scholar] [CrossRef] [PubMed]










| Metric | Formula | Meaning |
|---|---|---|
| Flux (F) | j≠i | Sum of potential dispersions from all patches |
| Equivalent Probability (EC) | Square root of the sum of products of capacity of all pairs of patches weighted by their interaction probability. | |
| Probability of Connectivity (PC) | This indicates probability that two points randomly placed in the study area are connected. | |
| Integral Index of Connectivity (IIC) | Product of patch capacities divided by the number of links between them, the sum is divided by the square of the area of the study zone. |
| Suitable Habitat | Core Habitat | |||||||
|---|---|---|---|---|---|---|---|---|
| Total | PA | TF | NF | Total | PA | TF | NF | |
| Blackbuck | 40.51 | 12.61 | 38.65 | 48.74 | 5.24 | 17.36 | 52.72 | 29.90 |
| Chinkara | 25.25 | 29.87 | 54.98 | 15.15 | 5.34 | 35.51 | 59.04 | 5.44 |
| FHA | 20.21 | 23.99 | 59.19 | 16.81 | 3.04 | 43.67 | 48.04 | 8.28 |
| Nilgai | 27.71 | 24.09 | 57.73 | 18.18 | 8.07 | 36.30 | 56.99 | 6.71 |
| Integrated Area | 52.88 | 16.79 | 41.12 | 42.08 | 21.70 | 36.74 | 51.48 | 11.78 |
| Connectivity Metric | Chinkara | Nilgai | Blackbuck | Four-Horned Antelope |
|---|---|---|---|---|
| Sum Flux | 59.78 | 18.01 | 11.65 | 25.33 |
| Equivalent Connectivity | 54.78 | 37.96 | 41.86 | 64.24 |
| Probability of connectivity | 30.01 | 14.41 | 17.53 | 41.26 |
| Integral index of connectivity | 17.49 | 10.41 | 7.78 | 23.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niyogi, R.; Sarkar, M.S.; Hazra, P.; Rahman, M.; Banerjee, S.; John, R. Habitat Connectivity for the Conservation of Small Ungulates in A Human-Dominated Landscape. ISPRS Int. J. Geo-Inf. 2021, 10, 180. https://doi.org/10.3390/ijgi10030180
Niyogi R, Sarkar MS, Hazra P, Rahman M, Banerjee S, John R. Habitat Connectivity for the Conservation of Small Ungulates in A Human-Dominated Landscape. ISPRS International Journal of Geo-Information. 2021; 10(3):180. https://doi.org/10.3390/ijgi10030180
Chicago/Turabian StyleNiyogi, Rajashekhar, Mriganka Shekhar Sarkar, Poushali Hazra, Masidur Rahman, Subham Banerjee, and Robert John. 2021. "Habitat Connectivity for the Conservation of Small Ungulates in A Human-Dominated Landscape" ISPRS International Journal of Geo-Information 10, no. 3: 180. https://doi.org/10.3390/ijgi10030180
APA StyleNiyogi, R., Sarkar, M. S., Hazra, P., Rahman, M., Banerjee, S., & John, R. (2021). Habitat Connectivity for the Conservation of Small Ungulates in A Human-Dominated Landscape. ISPRS International Journal of Geo-Information, 10(3), 180. https://doi.org/10.3390/ijgi10030180