
Tree Height Growth Modelling Using LiDAR-Derived Topography Information


Abstract
:1. Introduction
2. Materials and Methods
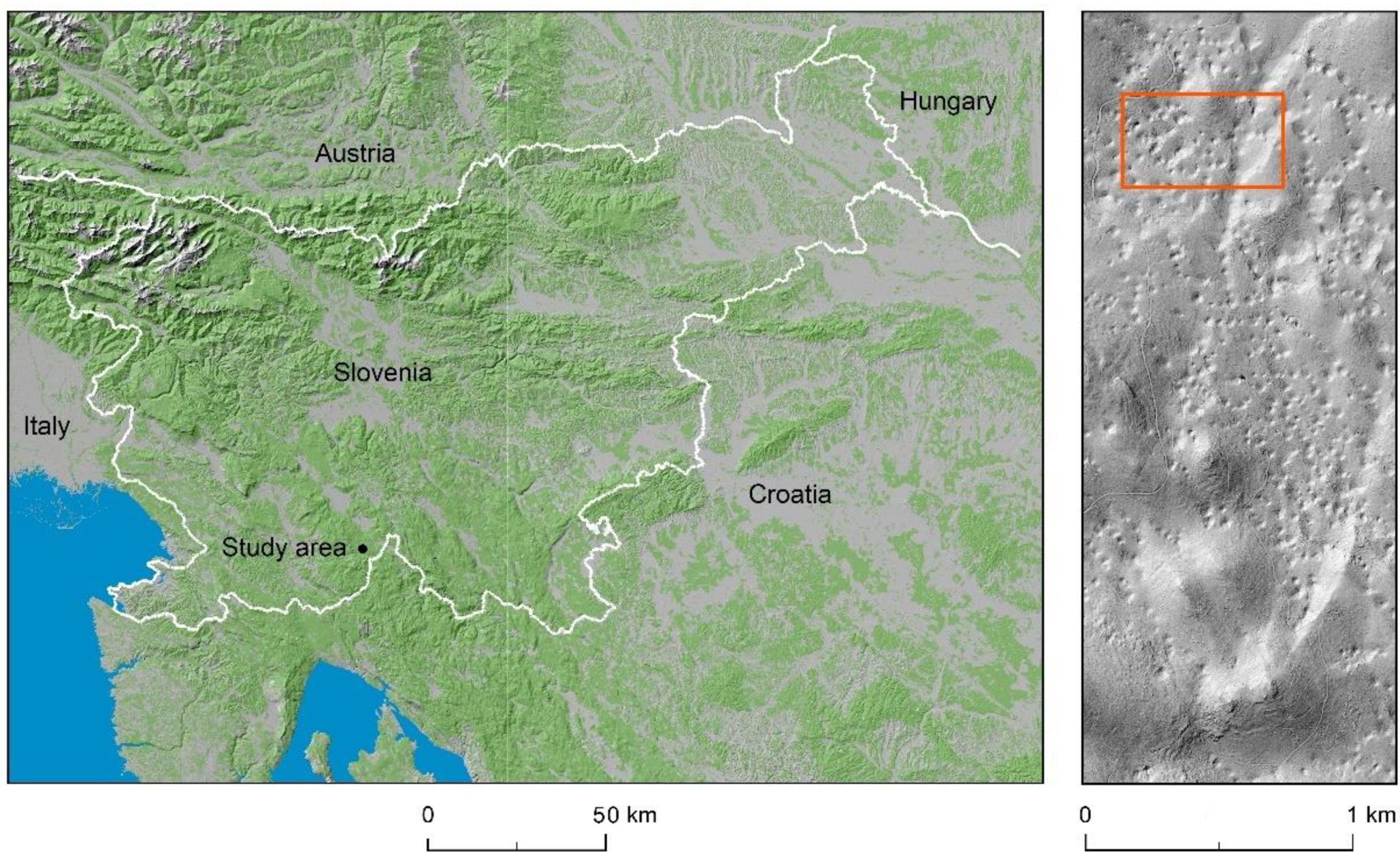
2.1. Study Area
2.2. Experimental Design
2.2.1. Plot Establishment
2.2.2. Airborne Laser Scanning
2.3. Laboratory Work
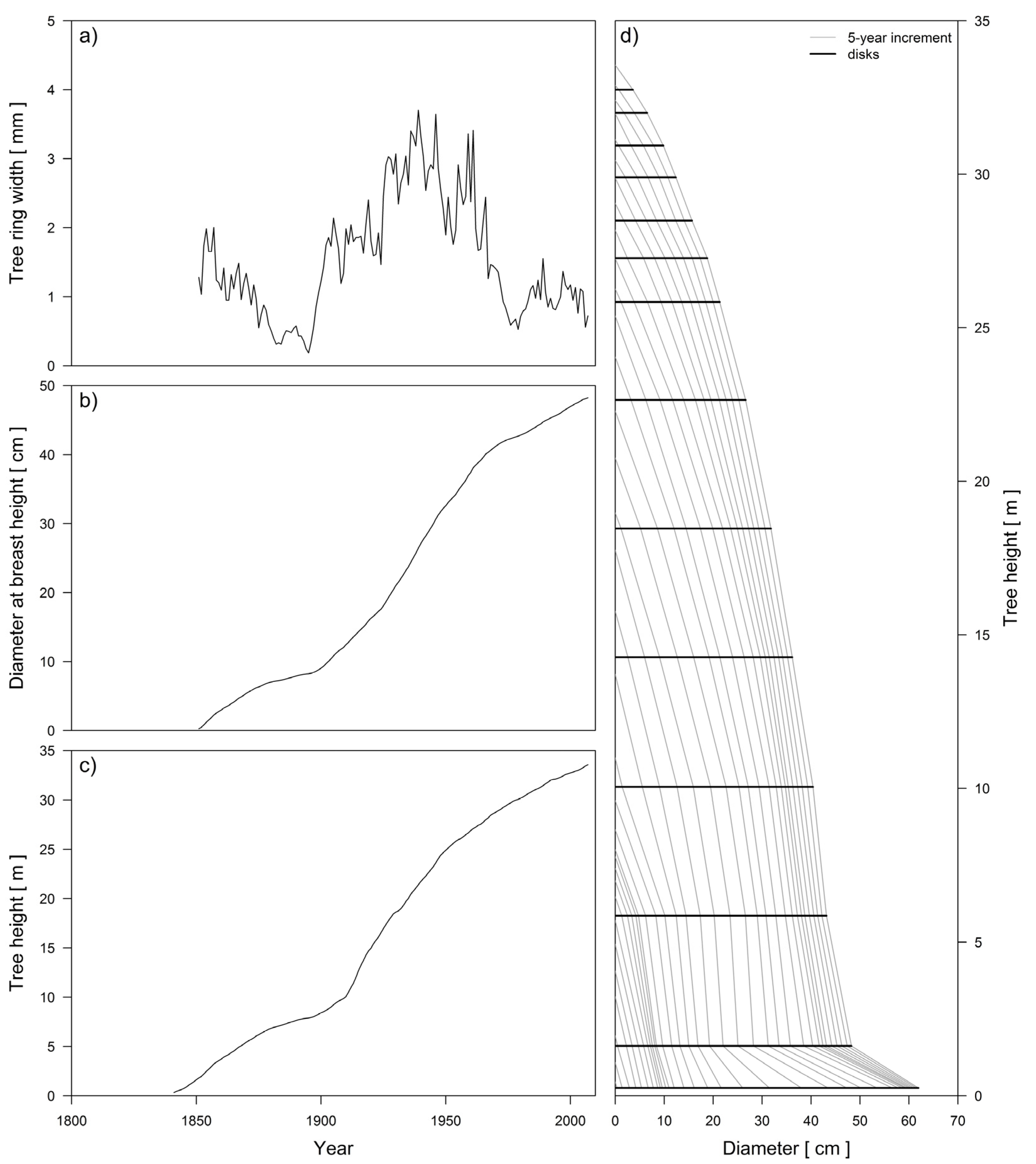
2.3.1. Detailed Stem Analysis
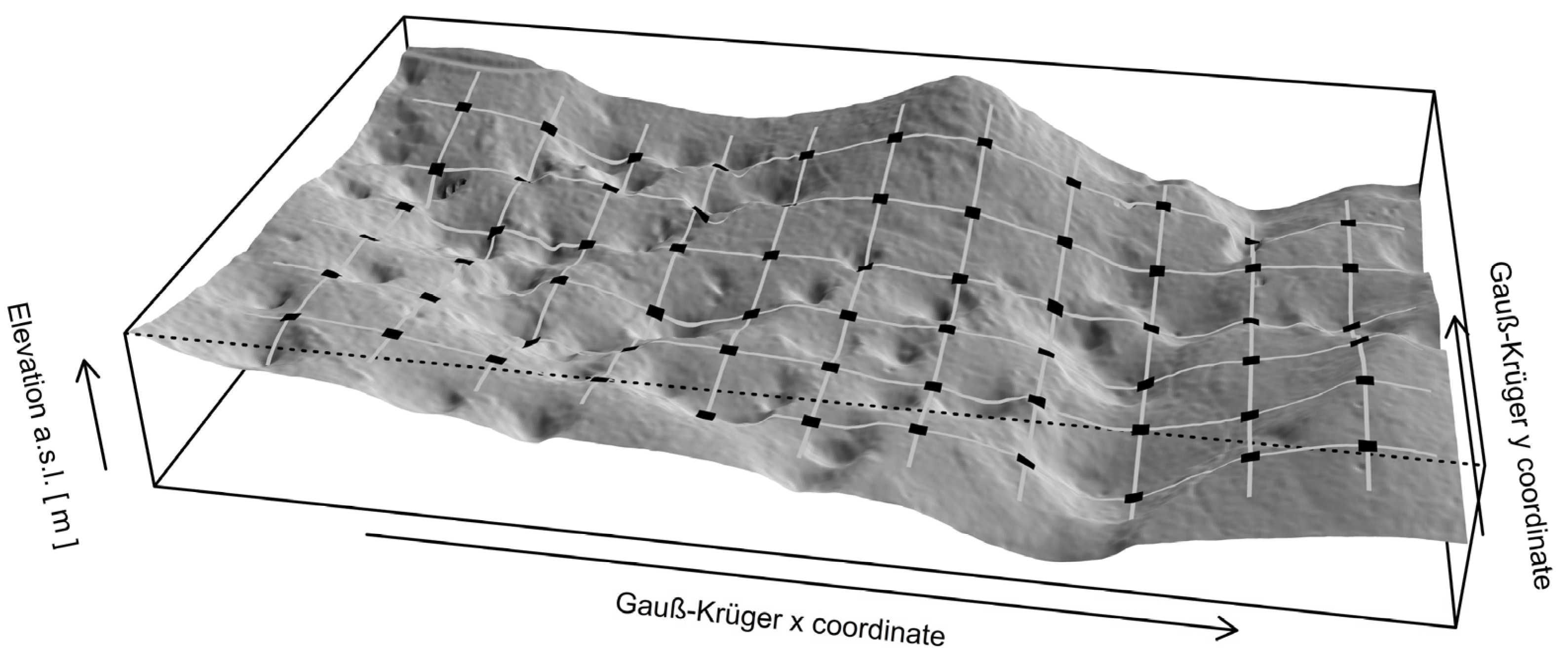
2.3.2. Digital Elevation Model Processing
| Ridge category | TPI > 0.5 × SD |
| Middle slope category | −0.5 × SD < TPI < 0.5 × SD |
| Sinkhole category | TPI ≤ −0.5 × SD |
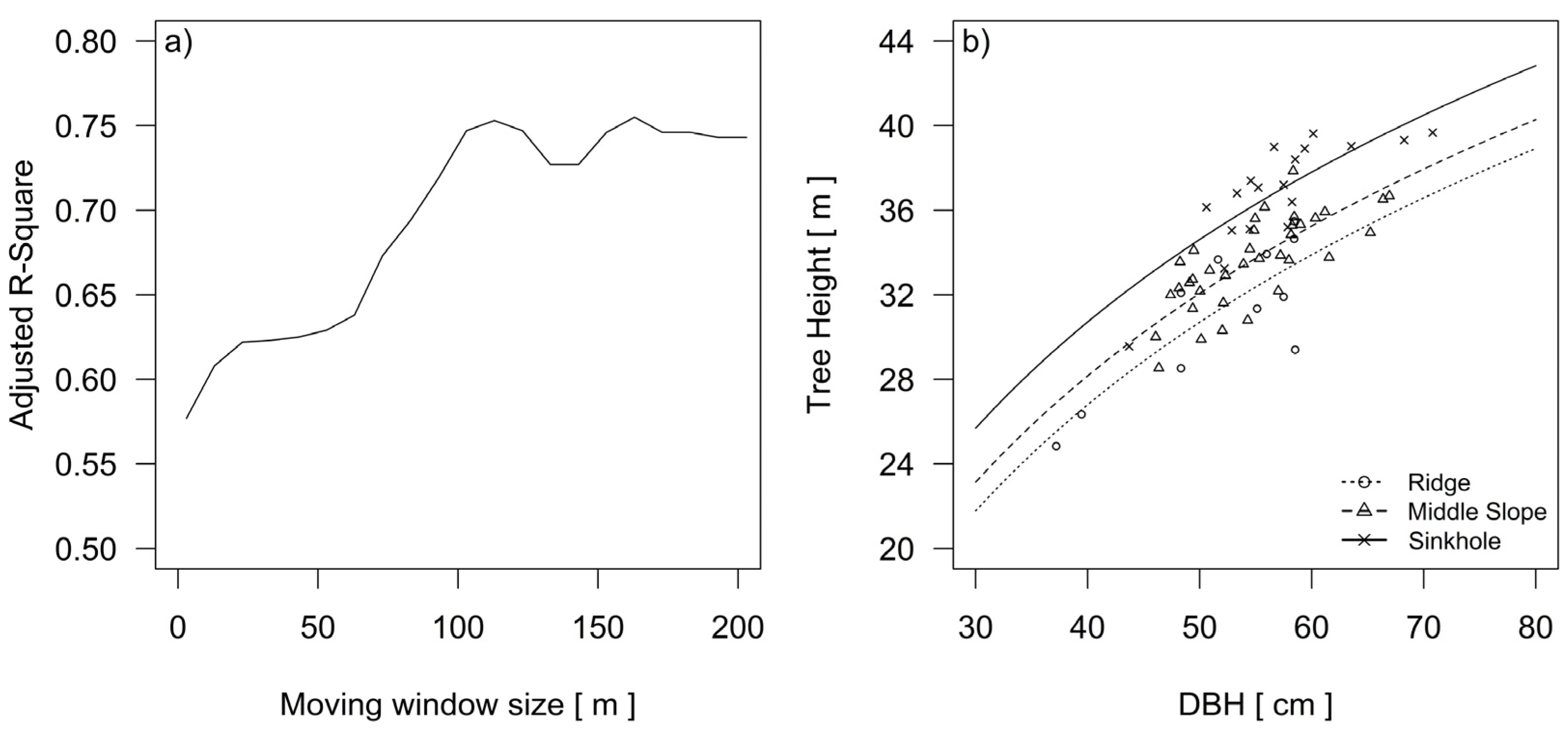
2.4. Statistical Analysis and Model Selection
3. Results
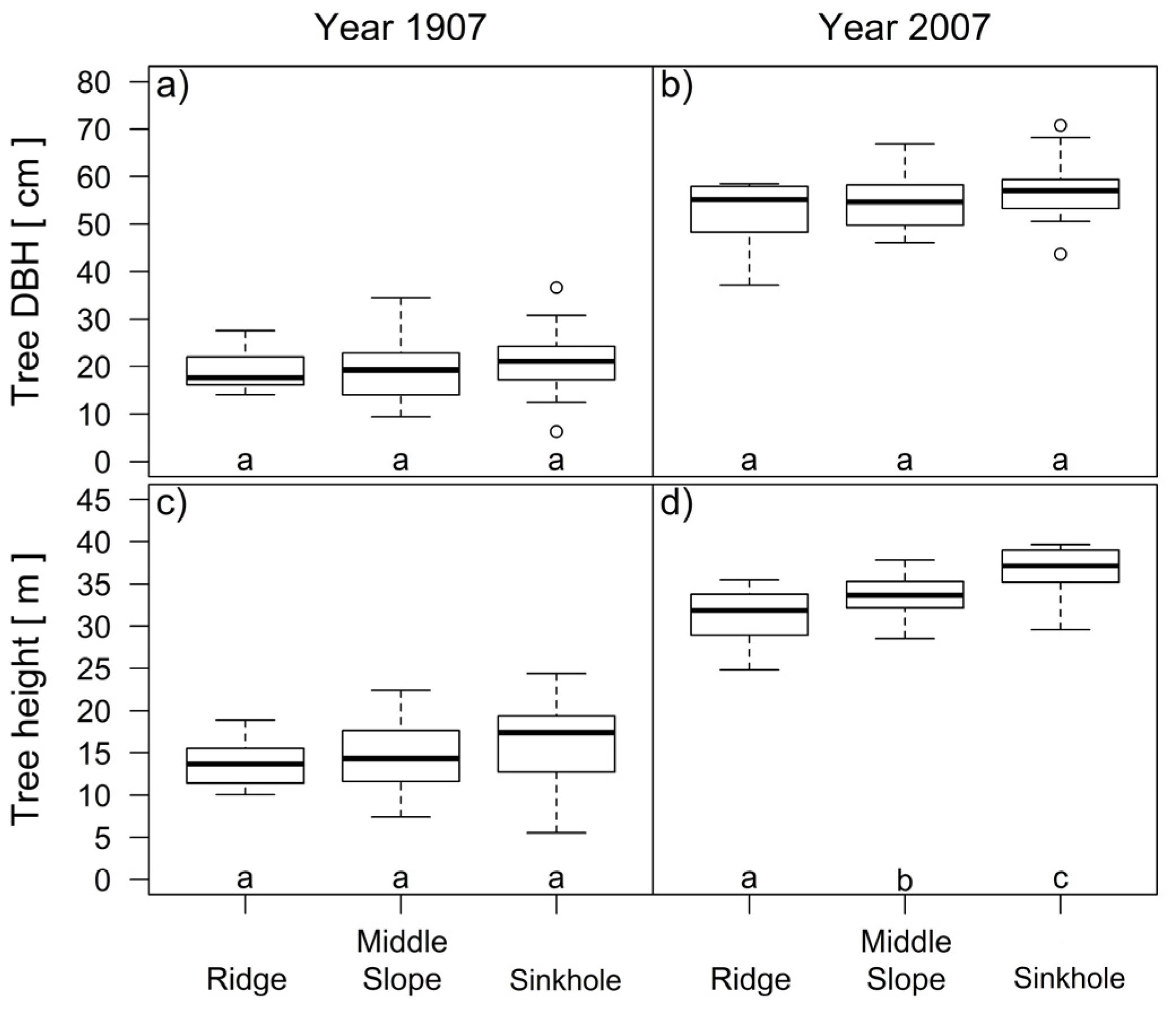
3.1. The Formation of Three Silver fir Groups
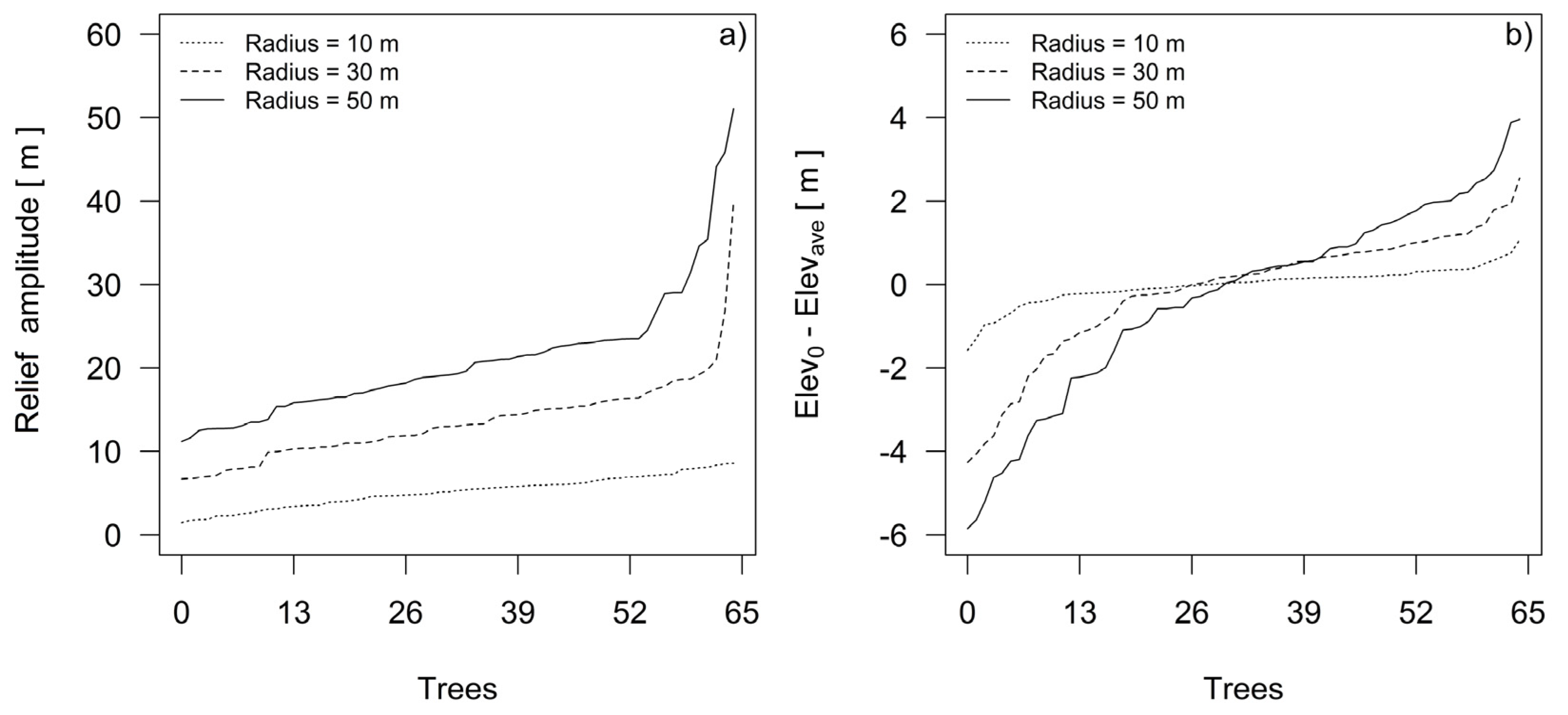
3.2. Spatial Model of Topographic Conditions
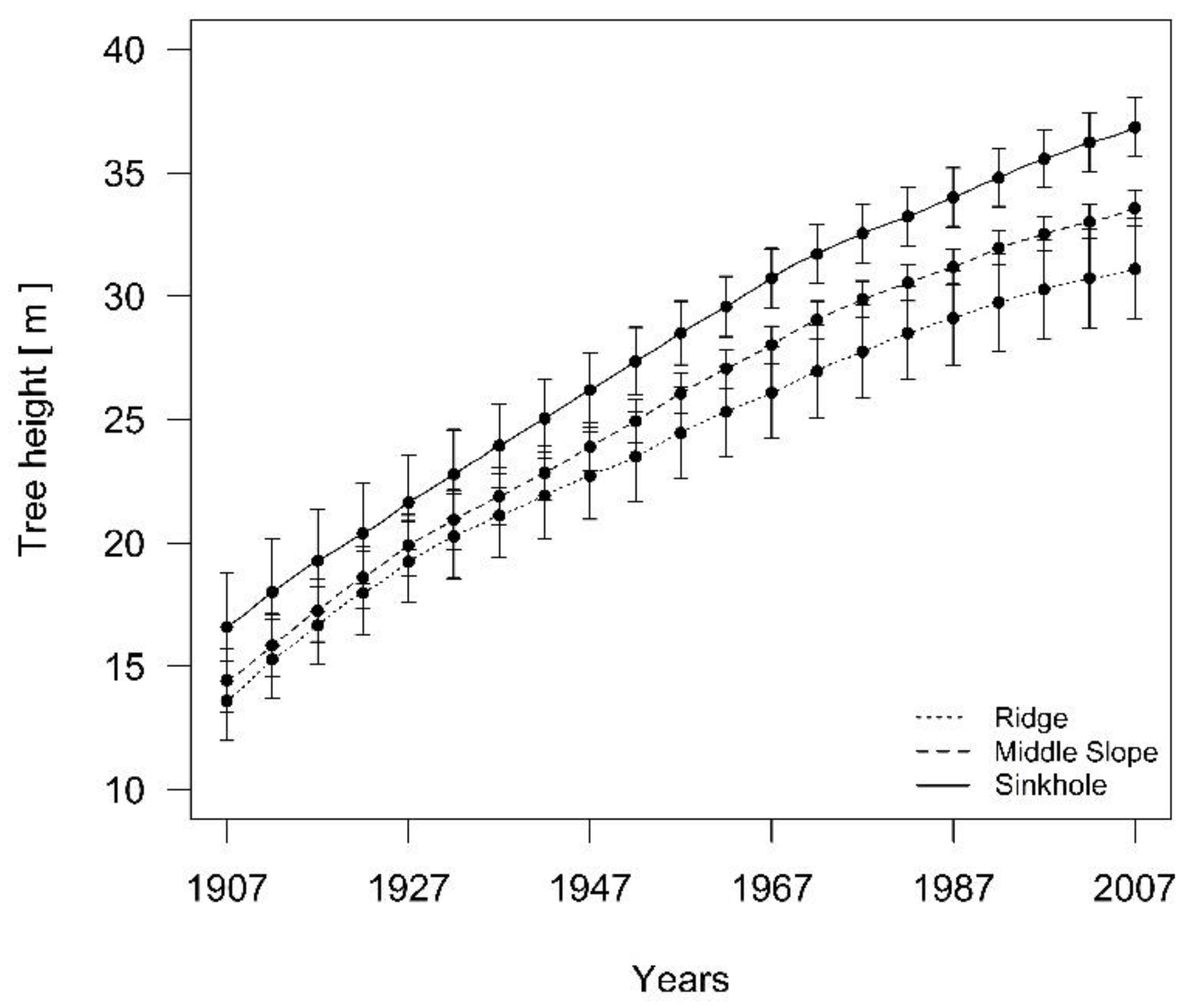
3.3. Differences in the Height Growth and Height Increment of Dominant Silver Fir Trees among Three Landform Categories
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Naveh, Z.; Lieberman, S. Landscape Ecology; Springer: New York, NY, USA, 1984; 356p. [Google Scholar]
- Zonneveld, I.S. The land unit—A fundamental concept in landscape ecology and its applications. Landsc. Ecol. 1989, 3, 67–86. [Google Scholar] [CrossRef]
- Haber, W. Basic Concepts of Landscape Ecology and Their Application in Land Management. Physiol. Ecol. Jpn. 1990, 27, 131–146. [Google Scholar]
- Forman, R.T.T. Land Mosaics; Cambridge University Press: Cambridge, UK, 1995; 632p. [Google Scholar]
- Skovsgaard, J.P.; Vanclay, J.K. Forest site productivity: A review of the evolution of dendrometric concepts for even-aged stands. Forestry 2008, 81, 13–31. [Google Scholar] [CrossRef] [Green Version]
- Bontemps, J.-D.; Bouriaud, O. Predictive approaches to forest site productivity: Recent trends, challenges and future perspectives. Forestry 2014, 87, 109–128. [Google Scholar] [CrossRef]
- Brandl, S.; Mette, T.; Falk, W.; Vallet, P.; Rötzer, T.; Pretzsch, H. Static site indices from different national forest inventories: Harmonization and prediction from site conditions. Ann. For. Sci. 2018, 75, 56. [Google Scholar] [CrossRef] [Green Version]
- Naveh, Z. Introduction to landscape ecology as a practical transdisciplinary science of landscape study, planning and management. In Atti del XXXI Corso di Cultura in Ecologia; Caattaneo, D., Semenzato, P., Eds.; SEDE: San Vito di Cadore, Italy, 1994; pp. 1–48. [Google Scholar]
- Davis, F.W.; Dozier, J. Information Analysis of a Spatial Database for Ecological Land Classification. Photogramm. Eng. Remote Sens. 1990, 56, 605–613. [Google Scholar]
- Franklin, J.; McCullough, P.; Gray, C. Terrain Variables Used for Predictive Mapping of Vegetation Communities in Southern California. In Terrain Analys: Principles and Applications; Wilson, J.P., Gallant, J.C., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2000; pp. 331–354. [Google Scholar]
- Bastian, O.; Steinhardt, U. Development and Perspectives of Landscape Ecology; Kluwer: Dordrecht, The Netherlands, 2002; 498p. [Google Scholar]
- Rasti, B.; Hong, D.; Hang, R.; Ghamisi, P.; Kang, X.; Chanussot, J.; Benediktsson, J.A. Feature Extraction for Hyperspectral Imagery. The evolution from shallow to deep: Overview and toolbox. IEEE Geosci. Remote Sens. Mag. 2020, 4, 60–88. [Google Scholar] [CrossRef]
- Hong, D.; Yokoya, N.; Chanussot, J.; Zhu, X.X. An Augmented Linear Mixing Model to Address Spectral Variability for Hyperspectral Unmixing. IEEE Trans. Image Process. 2019, 28, 1923–1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, D.; Gao, L.; Yao, J.; Zhang, B.; Plaza, A.; Chanussot, J. Graph Convolutional Networks for Hyperspectral Image Classification. IEEE Trans. Geosci. Remote Sens. 2020, 1–13. [Google Scholar] [CrossRef]
- Muster, S.; Elsenbeer, H.; Conedera, M. Small-scale effects of historical land use and topography on post-cultural tree species composition in an Alpine valley in southern Switzerland. Landsc. Ecol. 2007, 22, 1187–1199. [Google Scholar] [CrossRef]
- Matsuura, T.; Suzuki, W. Analysis of topography and vegetation distribution using a digital elevation model: Case study of a snowy mountain basin in northeastern Japan. Landsc. Ecol. Eng. 2013, 9, 143–155. [Google Scholar] [CrossRef]
- Reger, B.; Härig, T.; Ewald, J. The TRM Model of Potential Natural Vegetation in Mountain Forests. Folia Geobot. 2014, 49, 337–359. [Google Scholar] [CrossRef]
- Pfeffer, K.; Pebesma, E.J.; Burrough, P.A. Mapping alpine vegetation using observations and topographic attributes. Landsc. Ecol. 2003, 18, 759–776. [Google Scholar] [CrossRef]
- Dirnboeck, T.; Dullinger, S.; Gottfried, M.; Ginzler, C.; Grabherr, G. Mapping alpine vegetation based on image analysis, topographic variables and Canonical Correspondence Analysis. Appl. Veg. Sci. 2003, 6, 85–96. [Google Scholar] [CrossRef]
- Mihevc, A. Kras morphology. In Slovene Classical Karst—Kras; Kranjc, A., Ed.; ZRC SAZU: Ljubljana, Slovenia, 1997; pp. 43–49. [Google Scholar]
- Urbančič, M.; Simončič, P.; Prus, T.; Kutnar, L. Atlas Gozdnih Tal Slovenije. Zveza Gozdarskih Društev Slovenije, Gozdarski Vestnik, Silva Slovenica; Gozdarski Inštitut Slovenije: Ljubljana, Slovenia, 2005; 100p. [Google Scholar]
- Kobal, M.; Bertoncelj, I.; Pirotti, F.; Dakskobler, I.; Kutnar, L. Using lidar data to analyse sinkhole characteristics relevant for understory vegetation under forest cover—case study of a high karst area in the Dinaric mountains. PLoS ONE 2015, 10, 3. [Google Scholar] [CrossRef] [Green Version]
- Kutnar, L.; Urbančič, M. Vpliv rastiščnih in sestojnih razmer na pestrost tal in vegetacije v izbranih bukovih in jelovo-bukovih gozdovih na Kočevskem. Zb. Gozdarstva Lesar. 2006, 80, 3–30. [Google Scholar]
- Kobal, M.; Grčman, H.; Zupan, M.; Levanič, T.; Simončič, P.; Kadunc, A.; Hladnik, D. Influence of soil properties on silver fir (Abies alba Mill.) growth in the Dinaric Mountains. For. Ecol. Manag. 2015, 337, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Robič, D.; Acetto, M. Ocena rastiščnih razmer na izbrani lokaciji in ekološke implikacije pri prebiralnem gospodarjenju z gozdovi. Gozdarski Vestn. 2002, 60, 343–351. [Google Scholar]
- Hasenauer, H. Concepts within Tree Growth Modelling. In Sustainable Forest Management—Growth Models for Europe; Hasenauer, H., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 3–17. [Google Scholar] [CrossRef]
- Van Laar, A.; Akça, A. Forest Mensuration; Springer: Dordrecht, The Netherlands, 2010; 383p. [Google Scholar]
- Sturtevant, B.R.; Seagle, S.W. Comparing estimates of forest site quality in old second-growth oak forests. For. Ecol. Manag. 2004, 191, 311–328. [Google Scholar] [CrossRef]
- Curt, T.; Bouchaud, M.; Agrech, G. Predicting site index of Douglas-fir plantations from ecological variables in the Massif Central area of France. For. Ecol. Manag. 2001, 149, 61–74. [Google Scholar] [CrossRef]
- Aertsen, W.; Kint, V.; Orshoven, J.V.; Muys, B. Evaluation of modelling techniques for forest site productivity prediction in contrasting ecoregions using stochastic multicriteria acceptability analysis (SMAA). Environ. Model. Softw. 2011, 26, 929–937. [Google Scholar] [CrossRef] [Green Version]
- Aertsen, W.; Kint, V.; Muys, B.; Orshoven, J.V. Effects of scale and scaling in predictive modelling of forest site productivity. Environ. Model. Softw. 2012, 31, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Kobler, A.; Pfeifer, N.; Ogrinc, P.; Todorovski, L.; Oštir, K.; Džeroski, S. Repetitive interpolation: A robust algorithm for DTM generation from Aerial Laser Scanner Data in forested terrain. Remote Sens. Environ. 2007, 108, 9–23. [Google Scholar] [CrossRef]
- Weishampel, J.F.; Hightower, J.N.; Chase, A.F.; Chase, D.Z.; Patrick, R.A. Detection and morphologic analysis of potential below-canopy cave openings in the karts landscape around the Maya polity of Caracol ising airborne LiDAR. J. Cave Karst Stud. 2011, 73, 187–196. [Google Scholar] [CrossRef]
- Bončina, A.; Diaci, J.; Cenčič, L. Comparison of the two main types of selection forests in Slovenia: Distribution, site conditions, stand structure, regeneration and management. Forestry 2002, 75, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Copernicus. Copernicus Land Monitoring Service. Reference Data: EU-DEM. 2015. Available online: http://land.copernicus.eu/pan-european/satellite-derived-products/eu-dem (accessed on 11 September 2018).
- Agencija RS Za Okolje. LIDAR. Available online: http://gis.arso.gov.si/evode/profile.aspx?id=atlas_voda_Lidar@Arso&culture=en-US (accessed on 23 June 2019).
- Bončina, A.; Gašperšič, F.; Diaci, J. Long-term changes in tree species composition in the Dinaric mountain forests of Slovenia. For. Chron. 2003, 79, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Braun-Blanquet, J. Pflanzensociologie. Grundzüge der Vegetations Kunde; Springer: Wien, Austria; New York, NY, USA, 1964; 865p. [Google Scholar]
- Kobal, M.; Kastelec, D.; Eler, K. Temporal changes of forest species composition studied by compositional data approach. iForest 2017, 10, 729–738. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H. Forest Dynamics, Growth and Yield; Springer: Berlin/Heidelberg, Germany, 2009; 664p. [Google Scholar]
- Kotar, M. Zgradba, Rast in Donos Gozda na Ekoloških in Fizioloških Osnovah; Zveza Gozdarskih Društev Slovenije, Zavod za Gozdove Slovenije: Ljubljana, Slovenia, 2005; 500p. [Google Scholar]
- Kotar, M. Volume and Height Growth of Fully Stocked Mature Beech Stands in Slovenia During the Past Three Decades. In Growth Trends in European Forests; Spiecker, H., Mielikäinen, K., Köhl, M., Skovsgaard, J.P., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 291–312. [Google Scholar]
- Kadunc, A. Prirastoslovne značinosti jelke (Abies alba Mill.) v Sloveniji. Gozdarski Vestn. 2010, 68, 403–422. [Google Scholar]
- Koivuniemi, J.; Korhonen, K.T. Inventory by Compartments. In Forest Inventory—Methodology and Applications; Kangas, A., Maltamo, M., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 271–278. [Google Scholar]
- Levanič, T. A new system for image acquisition in dendrochronology. Tree Ring Res. 2007, 63, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Guay, R.; Gagnon, R.; Morin, H. MacDENDRO, a new automatic and interactive tree ring measurement system based on image processing. In Tree Rings and Environment, Proceedings of the International Dendrochronological Symposium, Ystad, Sweden, 3–9 September 1990; Lundqua Report; Lund University, Department of Quaernary Geology: Lund, Sweden, 1992; pp. 128–129. [Google Scholar]
- Baillie, M.G.L.; Pilcher, J.R. A simple cross-dating programme for tree-ring research. Tree Ring Bull. 1973, 33, 7–14. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; ISBN 3-900051-07-0. [Google Scholar]
- Carmean, W.H. Site index curves for upland oaks in the central states. For. Sci. 1972, 18, 109–120. [Google Scholar] [CrossRef]
- Sharma, R.P.; Brunner, A.; Eid, T.; Øyen, B.H. Modelling dominant height growth from national forest inventory individual tree data with short time series and large age errors. For. Ecol. Manag. 2011, 262, 2162–2175. [Google Scholar] [CrossRef]
- Cunningham, D.; Grebby, S.; Tansey, K.; Gosar, A.; Kastelic, V. Application of airborne LiDAR to mapping seismogenic faults in forested mountainous terrain, SE Alps, Slovenia. Geophys. Res. Lett. 2006, 33, L20308. [Google Scholar] [CrossRef] [Green Version]
- Weiss, A. Topographic Position and Landforms Analysis. Poster Presentation. In Proceedings of the ESRI User Conference, San Diego, CA, USA, 9–13 July 2001. [Google Scholar]
- Jenness, J. Topographic Position Index (tpi_jen.avx) Extension for ArcView 3.2, v. 1.3a; Jenness Enterprises: Flagstaff, AZ, USA, 2006. [Google Scholar]
- Badoux, E. Ertragstafeln für Fichte, Tanne, Buche und Lärche; Eidgenössische Anstalt für das Forstliche Versuchswesen, (WSL): Zürich, Switzerland, 1969. [Google Scholar]
- Halaj, J.; Grék, J.; Pánek, F.; Petráš, R.; Řehák, J. Rastové Tabuľky Hlavných Drevín ČSSR; Príroda: Bratislava, Slovakia, 1987; 361p. [Google Scholar]
- Skovsgaard, J.P.; Vanclay, J.K. Forest site productivity: A review of spatial and temporal variability in natural site conditions. Forestry 2013, 86, 305–315. [Google Scholar] [CrossRef]
- Landolt, E. Oekologische Zeigerwerte der Schweizer Flora; Veröffentlichungen des Geobotanischen Institutes der ETH, Stiftung Rübel: Zürich, Switzerland, 1977; Volume 46, 208p. [Google Scholar]
- Ellenberg, H.; Weber, E.H.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Zeigerwerte von Pflanzen in Mitteleuropa; Scripta Geobotanika: Göttingen, Germany, 1991; 248p. [Google Scholar]
- Košir, Ž. Vrednotenje Proizvodne Sposobnosti Gozdnih Rastišč in Ekološkega Značaja Fitocenoz; Ministrstvo za Kmetijstvo in Gozdarstvo RS: Ljubljana, Slovenia, 1992; 58p.
- Landolt, E.; Bäumler, B.; Erhardt, A.; Hegg, O.; Klötzli, F.; Lämmler, W.; Nobis, M.; Rudmann-Maurer, K.; Schweingruber, F.; Theurillat, J.-P.; et al. Flora Indicativa. Ökologische Zeigerwerte und Biologische Kennzeichen zur Flora der Schweiz und der Alpen; Editions des Conservatoire et Jardin Botaniques de la Ville de Genève; Haupt-Verlag: Bern, Switzerland; Stuttgart, Germany; Wien, Austria, 2010; 378p. [Google Scholar]
- Bergès, L.; Gégout, J.C.; Franc, A. Can understory vegetation accurately predict site index? A comparative study using floristic and abiotic indices in sessile oak (Quercus petraea Liebl.) stands in northern France. Ann. For. Sci. 2006, 63, 31–42. [Google Scholar] [CrossRef]
- Kobal, M. Vpliv sestojnih, Talnih in Mikrorastiščnih Razmer na Rast in Razvoj Jelke (Abies Alba Mill.) na Visokem Krasu Snežnika. Ph.D. Thesis, Univerza v Ljubljani, BF, Ljubljana, Slovenia, 2011; 148p. [Google Scholar]
- Bátori, Z.; Körmöczi, L.; Erdős, L.; Zalatnai, M.; Csiky, J. Importance of karst sinkholes in preserving relict, mountain, and wetwoodland plant species under sub-Mediterranean climate. A case study from southern Hungary. J. Cave Karst Stud. 2012, 74, 127–134. [Google Scholar] [CrossRef]
- Kotar, M.; Robič, D. Povezanost proizvodne sposobnosti bukovih gozdov v Sloveniji z njihovo floristično sestavo. Gozdarski Vestn. 2001, 59, 227–247. [Google Scholar]
- Monserud, R.A.; Sterba, H. A basal area increment model for individual trees growing in even- and uneven-aged forest stands in Austria. For. Ecol. Manag. 1996, 80, 57–80. [Google Scholar] [CrossRef]
- Coomes, D.A.; Allen, R.B. Effects of size, competition and altitude on tree growth. J. Ecol. 2007, 95, 1084–1097. [Google Scholar] [CrossRef]
- D’Amato, A.W.; Puettmann, K. The relative dominance hypothesis explains interaction dynamics in mixed species Alnus rubra/Pseudotsuga menziesii stands. J. Ecol. 2004, 92, 450–463. [Google Scholar] [CrossRef]
- Furlani, S.; Cucchi, F.; Forti, F.; Rossi, A. Comparison between coastal and inland Karst limestone lowering rates in the northeastern Adriatic Region (Italy and Croatia). Geomorphology 2009, 104, 73–81. [Google Scholar] [CrossRef]
- Zald, H.S.J.; Spies, T.A.; Huso, M.; Gatziolis, D. Climatic, landform, microtopographic, and overstory canopy controls of tree invasion in a subalpine meadow landscape, Oregon Cascades, USA. Landsc. Ecol. 2012, 27, 1197–1212. [Google Scholar] [CrossRef] [Green Version]
- Trevisani, S.; Cavalli, M.; Marchi, L. Surface texture analysis of a high-resolution DTM: Interpreting an alpine basin. Geomorphology 2012, 161–162, 26–39. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Model | Height Curve | SE | Adjusted R2 | AIC |
|---|---|---|---|---|---|
| M1 | All | H = −48.53 + 20.65 × ln(dbh) | 2.04 | 0.56 | 280.89 |
| Ridge | H = −37.68 + 17.48 × ln(dbh) | ||||
| M2 | Middle slope | H = −36.32 + 17.48 × ln(dbh) | 1.55 | 0.75 | 247.67 |
| Sinkhole | H = −33.76 + 17.48 × ln(dbh) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobal, M.; Hladnik, D. Tree Height Growth Modelling Using LiDAR-Derived Topography Information. ISPRS Int. J. Geo-Inf. 2021, 10, 419. https://doi.org/10.3390/ijgi10060419
Kobal M, Hladnik D. Tree Height Growth Modelling Using LiDAR-Derived Topography Information. ISPRS International Journal of Geo-Information. 2021; 10(6):419. https://doi.org/10.3390/ijgi10060419
Chicago/Turabian StyleKobal, Milan, and David Hladnik. 2021. "Tree Height Growth Modelling Using LiDAR-Derived Topography Information" ISPRS International Journal of Geo-Information 10, no. 6: 419. https://doi.org/10.3390/ijgi10060419
APA StyleKobal, M., & Hladnik, D. (2021). Tree Height Growth Modelling Using LiDAR-Derived Topography Information. ISPRS International Journal of Geo-Information, 10(6), 419. https://doi.org/10.3390/ijgi10060419