Genotype-by-Sequencing Analysis of Mutations and Recombination in Pepper Progeny of Gamma-Irradiated Gametophytes




Abstract
:1. Introduction
2. Results
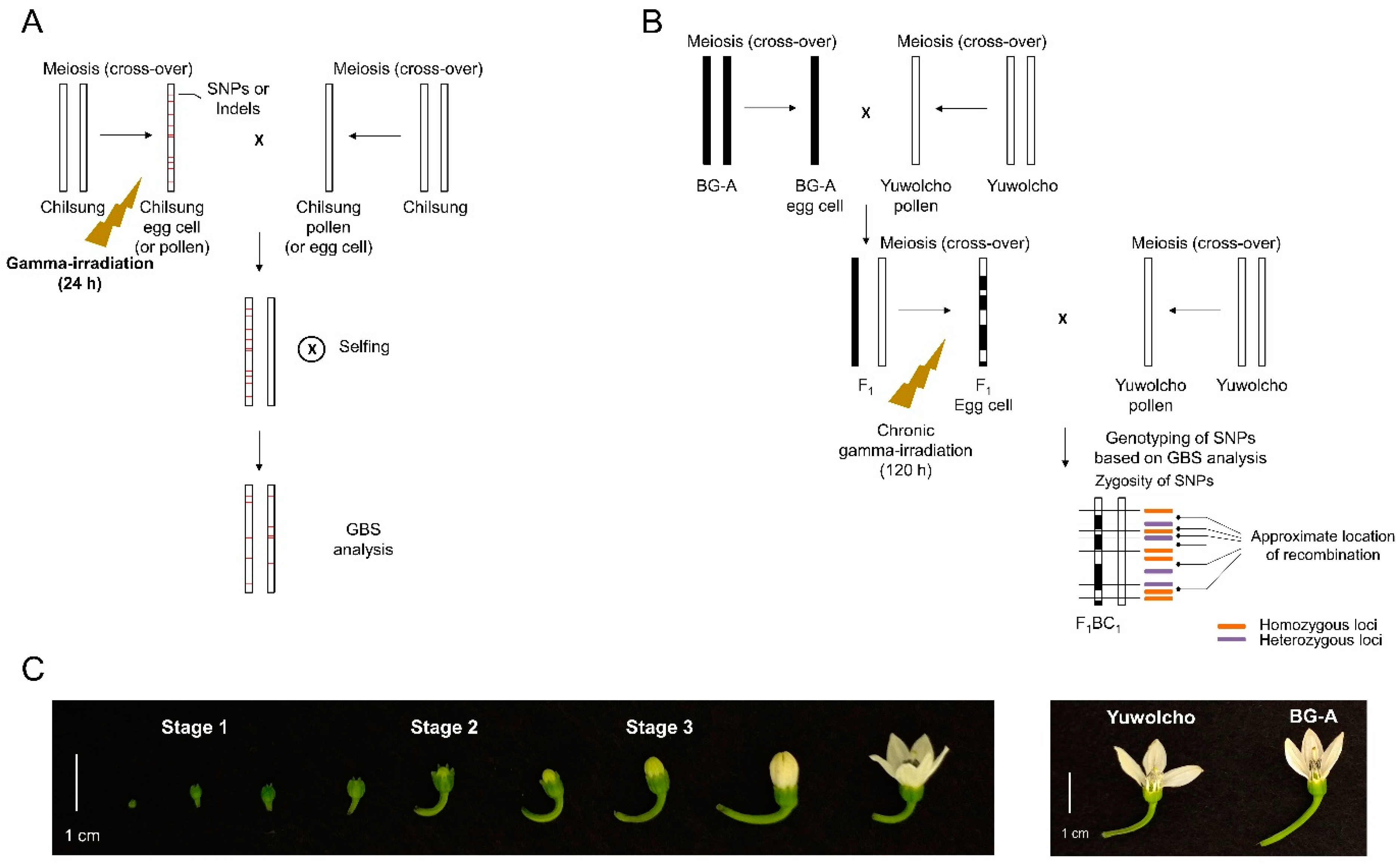
2.1. Development of Pepper Lines by Artificial Crosses Using Irradiated Reproductive Organs, and a GBS Analysis of the Resulting DNA Mutations
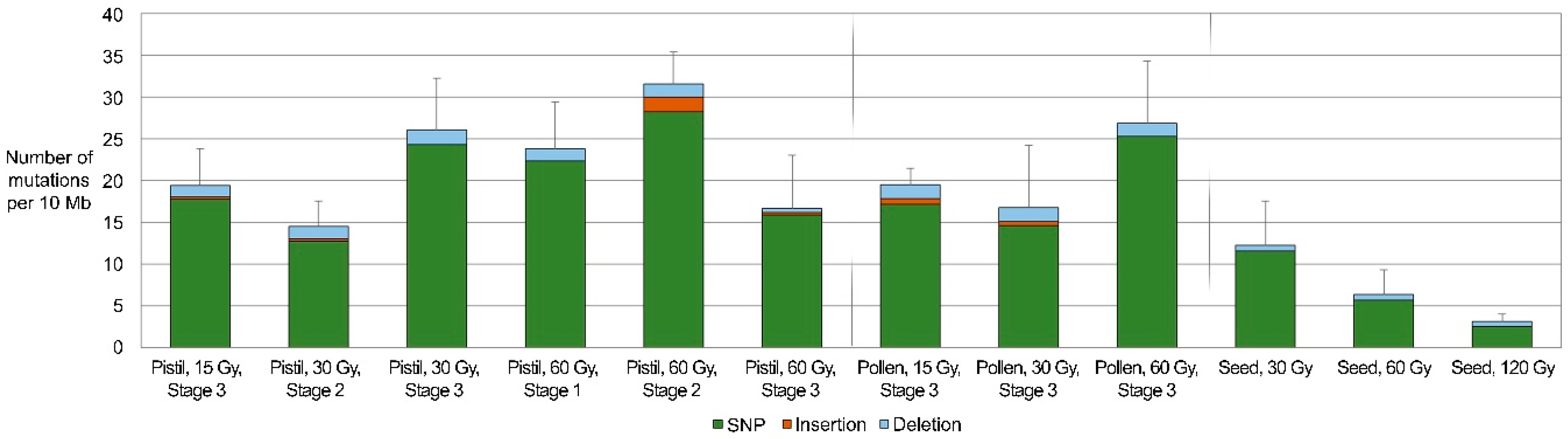
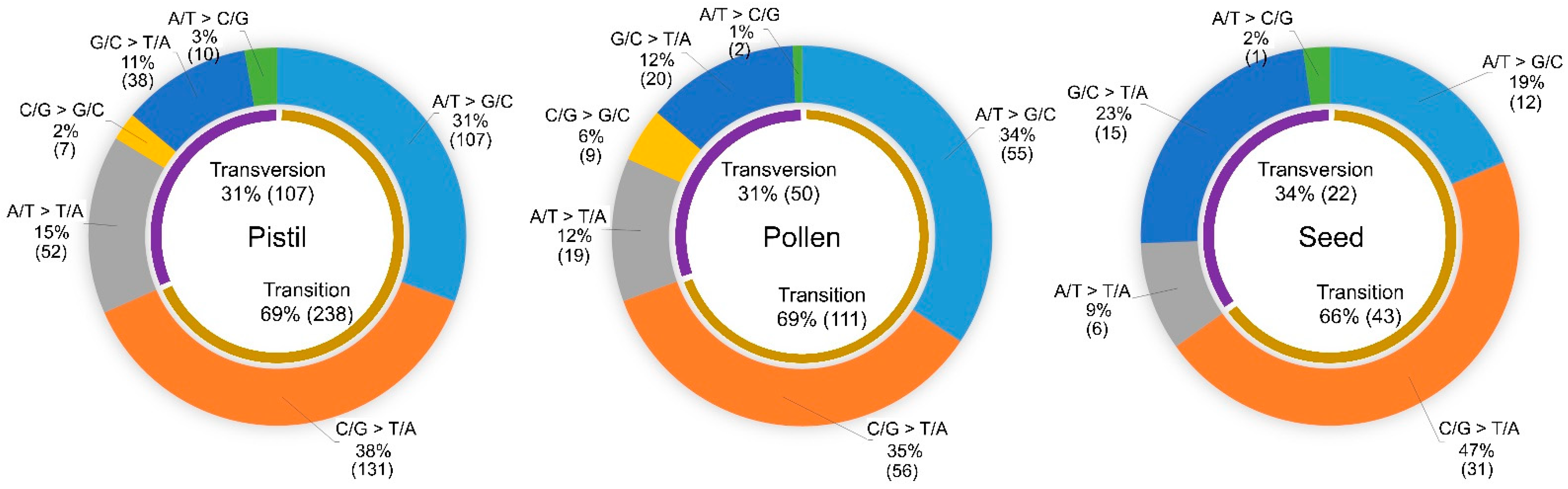
2.2. Frequencies and Spectra of DNA Mutations Induced by the Irradiation of Reproductive Organs
2.3. Development of Progeny Lines by Artificial Crosses Using Irradiated Reproductive Organs, and a GBS Analysis of the Resulting Meiotic Recombination Frequencies
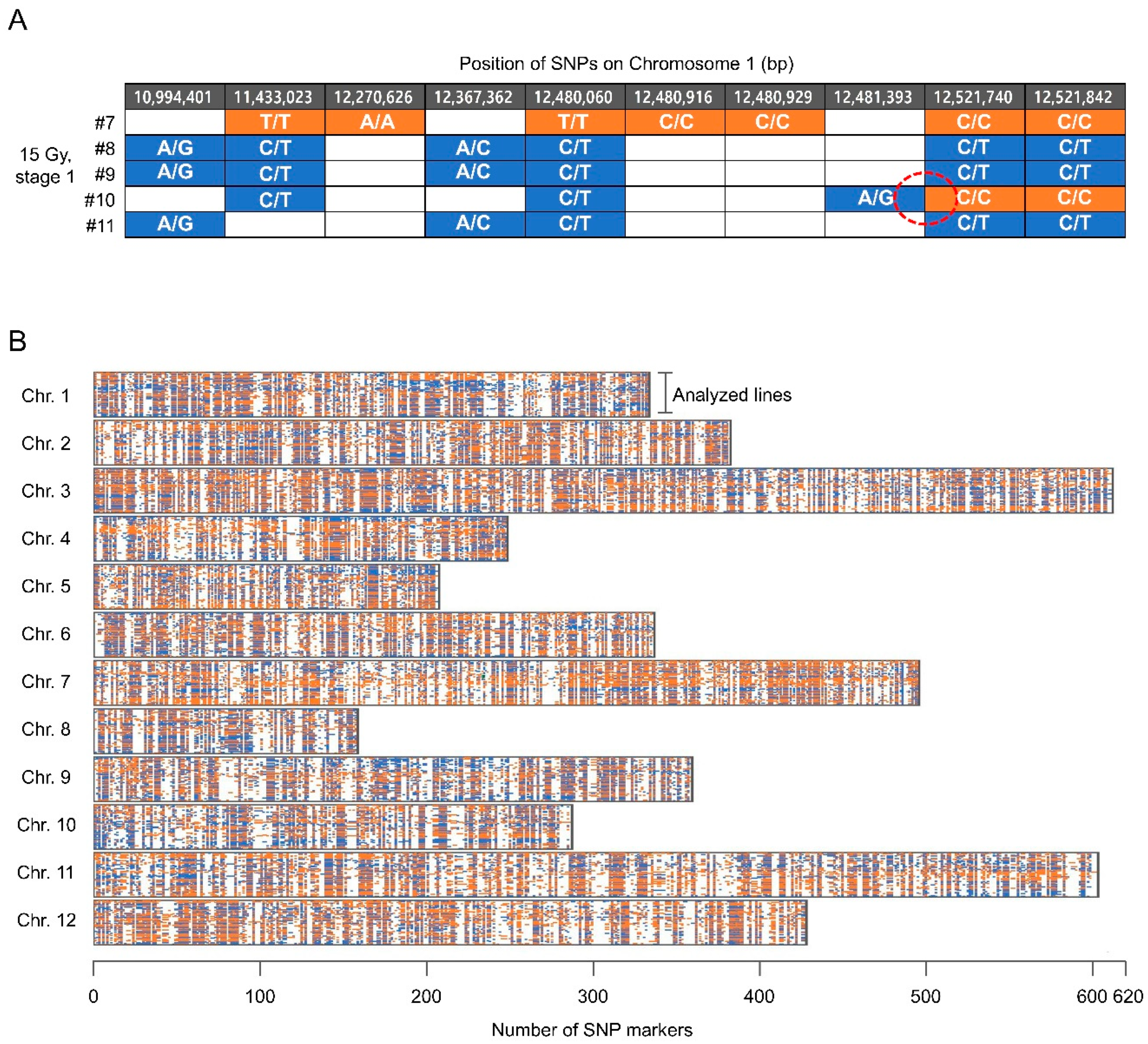
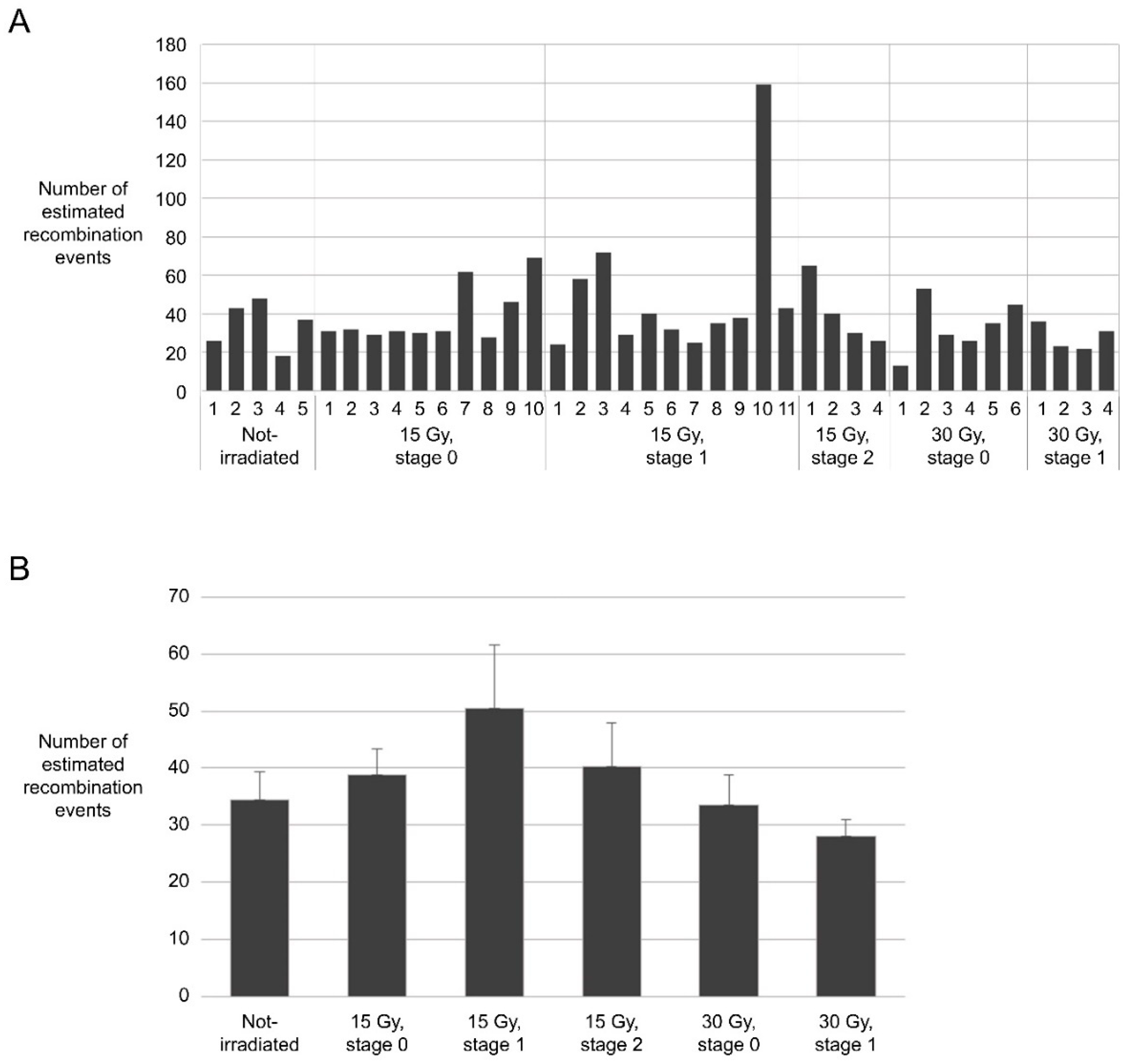
2.4. Frequency of Meiotic Recombiantion in the Irradiated Reproductive Organs
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Gamma-Irradiation
4.2. GBS Analysis
4.3. Analysis of Mutation Frequencies and Spectra
4.4. Analysis of Recombination
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GBS | Genotype-by-sequencing |
| SBS | Single-base substitution |
| InDel | Insertion and deletion |
| CMS | Cytoplasmic male sterility |
| SBS | Single-base substitution |
| SNP | Single nucleotide polymorphism |
References
- Stadler, L.J. Mutations in barley induced by X-rays and radium. Science 1928, 68, 186–187. [Google Scholar] [CrossRef] [PubMed]
- Stadler, L.J. Genetic effects of X-rays in maize. Proc. Natl. Acad. Sci. USA 1928, 14, 69–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harn, C.H.; Jung, M.; Jang, C.; Shin, J.S.; Lee, S.W.; In, D.-S.; Joo, Y. NWB-CMS Brassica oleracea Having Cytoplasmic Male Sterility and Use Thereof. US Patent US 2017/0367289 A1, 28 December 2017. [Google Scholar]
- Sugiyama, K.; Morishita, M. Production of seedless watermelon using soft-X-irradiated pollen. Sci. Hort. 2000, 84, 255–264. [Google Scholar] [CrossRef]
- Preuss, D.; Copenhaver, G.P. Chemical and Physical Treatment That Stimulate Recombination. International Application WO/2000/054574, 21 September 2000. [Google Scholar]
- Jo, Y.D.; Kim, J.-B. Frequency and spectrum of radiation-induced mutations revealed by whole-genome sequencing analyses of plants. Quantum Beam Sci. 2019, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Shikazono, N.; Suzuki, C.; Kitamura, S.; Watanabe, H.; Tano, S. Analysis of mutations induced by carbon ions in Arabidopsis thaliana. J. Exp. Bot. 2005, 56, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Kazama, Y.; Hirano, T.; Saito, H.; Liu, Y.; Ohbu, S.; Hayashi, Y.; Abe, T. Characterization of highly efficient heavy-ion mutagenesis in Arabidopsis thaliana. BMC Plant Biol. 2011, 11, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Chern, M.; Jain, R.; Martin, J.A.; Schackwitz, W.S.; Jiang, L.; Vega-Sánchez, M.E.; Lipzen, A.M.; Barry, K.W.; Schmutz, J.; et al. Genome-wide sequencing of 41 rice (Oryza sativa L.) mutated lines reveals diverse mutations induced by fast neutron irradiation. Mol. Plant 2016, 9, 1078–1081. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Jain, R.; Chern, M.; Pham, N.T.; Martin, J.A.; Wei, T. The sequences of 1504 mutants in the model rice variety Kitaake facilitate rapid functional genomic studies. Plant Cell 2017, 29, 1218–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazama, Y.; Ishii, K.; Hirano, T.; Wakana, T.; Yamada, M.; Ohbu, S.; Abe, T. Different mutational function of low- and high-linear energy transfer heavy-ion irradiation demonstrated by whole-genome resequencing of Arabidopsis mutants. Plant J. 2017, 92, 1020–1030. [Google Scholar] [CrossRef] [Green Version]
- Hase, Y.; Satoh, K.; Seito, H.; Oono, Y. Genetic consequences of acute/chronic gamma and carbon ion irradiation of Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 336. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.J.; Ryu, J.; Im, J.; Kim, S.H.; Kang, S.-Y.; Lee, J.-H.; Jo, S.-H.; Ha, B.-K. Molecular characterization of proton beam-induced mutations in soybean using genotyping-by-sequencing. Mol. Genet. Genom. 2018, 293, 1169–1180. [Google Scholar] [CrossRef]
- Ryu, J.; Kim, W.J.; Im, J.; Kang, K.-W.; Kim, S.H.; Jo, Y.D.; Kang, S.-Y.; Lee, J.-H.; Ha, B.-K. Single nucleotide polymorphism (SNP) discovery through genotyping-by-sequencing (GBS) and genetic characterization of Dendrobium mutants and cultivars. Sci. Hortic. 2019, 244, 225–233. [Google Scholar] [CrossRef]
- Hase, Y.; Satoh, K.; Kitamu, S.; Oono, Y. Physiological status of plant tissue affects the frequency and types of mutations induced by carbon-ion irradiation in Arabidopsis. Sci. Rep. 2018, 8, 1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saccardo, F. Gametophyte irradiation in some dicotyledonous crop species. Prospects and results. IAEA TECDOC 1983, 289, 31–32. [Google Scholar]
- Viccini, L.F.; De Carvalho, C.R. Meiotic chromosomal variation resulting from irradiation of pollen in maize. J. Appl. Genet. 2002, 43, 463–469. [Google Scholar] [PubMed]
- Kowyama, Y.; Saba, T.; Tsuji, T.; Kawase, T. Specific developmental stages of gametogenesis for radiosensitivity and mutagenesis in rice. Euphytica 1994, 80, 27–38. [Google Scholar] [CrossRef]
- Blary, A.; Jenczewski, E. Manipulation of crossover frequency and distribution for plant breeding. Theor. Appl. Genet. 2019, 132, 575–592. [Google Scholar] [CrossRef] [Green Version]
- Wijnker, E.; de Jong, H. Managing meiotic recombination in plant breeding. Trends Plant Sci. 2008, 13, 640–646. [Google Scholar] [CrossRef]
- Lloyd, A.; Morgan, C.; FC, H.F.; Bomblies, K. Plasticity of meiotic recombination rates in response to temperature in Arabidopsis. Genetics 2018, 208, 1409–1420. [Google Scholar] [CrossRef] [Green Version]
- Modliszewski, J.L.; Wang, H.; Albright, A.R.; Lewis, S.M.; Bennett, A.R.; Huang, J.; Ma, H.; Wang, Y.; Copenhaver, G.P. Elevated temperature increases meiotic crossover frequency via the interfering (Type I) pathway in Arabidopsis thaliana. PLoS Genet. 2018, 14, e1007384. [Google Scholar] [CrossRef]
- Law, C.N. An effect of potassium on chiasma frequency and recombination. Genetica 1963, 33, 313–329. [Google Scholar] [CrossRef]
- Shu, Q.Y.; Forster, B.P.; Nakagawa, H. Plant Mutation Breeding and Biotechnology; CAB International: Wallingford, UK, 2012; ISBN 9789251070222. [Google Scholar]
- Sandoval-Oliveros, R.; Guevara-Olvera, L.; Beltran, J.P.; Gomez-Mena, C.; Acosta-Garcia, G. Developmental landmarks during floral ontogeny of jalapeno chili pepper (Capsicum annuum L.) and the effect of gibberellin on ovary growth. Plant Reprod. 2017, 30, 119–129. [Google Scholar] [CrossRef]
- Shirasawa, K.; Hirakawa, H.; Nunome, T.; Tabata, S.; Isobe, S. Genome-wide survey of artificial mutations induced by ethyl methanesulfonate and gamma rays in tomato. Plant Biotechnol. J. 2016, 14, 51–60. [Google Scholar] [CrossRef]
- Saito, T.; Ariizumi, T.; Okabe, Y.; Asamizu, E.; Hiwasa-Tanase, K.; Fukuda, N.; Mizoguchi, T.; Yamazaki, Y.; Aoki, K.; Ezura, H. TOMATOMA: A novel tomato mutant database distributing Micro-Tom mutant collections. Plant Cell Physiol. 2011, 52, 283–296. [Google Scholar] [CrossRef] [Green Version]
- Ossowski, S.; Schneeberger, K.; Lucas-Lledo, J.I.; Warthmann, N.; Clark, R.M.; Shaw, R.G.; Weigel, D.; Lynch, M. The rate and molecular spectrum of spontaneous mutations in Arabidopsis thaliana. Science 2010, 327, 92–94. [Google Scholar] [CrossRef] [Green Version]
- Taheri, S.; Abdullah, T.L. TILLING, high-resolution melting (HRM), and next-generation sequencing (NGS) techniques in plant mutation breeding. Mol. Breed. 2017, 40, 37. [Google Scholar] [CrossRef]
- Naito, K.; Kusaba, M.; Shikazono, N.; Takano, T.; Tanaka, A.; Tanisaka, T.; Nishimura, M. Transmissible and nontransmissible mutations induced by irradiating Arabidopsis thaliana pollen With λ-rays and carbon ions. Genetics 2005, 169, 881–889. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17. [Google Scholar] [CrossRef]
- Cox, M.P.; Peterson, D.A.; Biggs, P.J. SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data. BMC Bioinform. 2010, 11, 485. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Genome Project Data Processing, S. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics of GBS Analysis | Average Among Lines | |
|---|---|---|
| Analysis of Mutation Frequency | Analysis of Recombination Frequency | |
| Number of raw reads | 7,701,874 | 5,575,688 |
| Total length of raw reads (bp) | 777,889,296 | 563,144,465 |
| Number of trimmed reads | 6,498,878 | 4,978,217 |
| Total length of trimmed reads (bp) | 498,825,520 | 417,179,109 |
| Average length of trimmed reads (bp) | 76.74 | 83.92 |
| Number of mapped reads (%) | 6,176,211 (95.10) | 3,143,631 (68.26) |
| Number of mapped region | 79,402 | 90,051 |
| Average depth of mapped region | 35.27 | 14.33 |
| Total length of mapped region (bp) | 11,449,437 | 14,965,995 |
| Average length of mapped region (bp) | 143.91 | 165.7 |
| Reference Genome coverage (%) | 0.37 | 0.49 |
| Characteristics of SNP Markers | Statistics |
|---|---|
| Total number of applied SNP markers z | 4467 |
| Average number of applied SNP markers (per line) | 1554 |
| Range of number of applied markers | 1399–1732 |
| Percentage of markers scored to be homozygous (%) | 55.78 |
| Percentage of markers scored to be heterozygous (%) | 44.24 |
| Average number of applied markers per chromosome | 129.50 |
| Average distance between applied markers on reference genome (Mb) | 1.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo, Y.D.; Kang, H.S.; Choi, H.-I.; Kim, J.-B. Genotype-by-Sequencing Analysis of Mutations and Recombination in Pepper Progeny of Gamma-Irradiated Gametophytes. Plants 2021, 10, 144. https://doi.org/10.3390/plants10010144
Jo YD, Kang HS, Choi H-I, Kim J-B. Genotype-by-Sequencing Analysis of Mutations and Recombination in Pepper Progeny of Gamma-Irradiated Gametophytes. Plants. 2021; 10(1):144. https://doi.org/10.3390/plants10010144
Chicago/Turabian StyleJo, Yeong Deuk, Han Sol Kang, Hong-Il Choi, and Jin-Baek Kim. 2021. "Genotype-by-Sequencing Analysis of Mutations and Recombination in Pepper Progeny of Gamma-Irradiated Gametophytes" Plants 10, no. 1: 144. https://doi.org/10.3390/plants10010144
APA StyleJo, Y. D., Kang, H. S., Choi, H.-I., & Kim, J.-B. (2021). Genotype-by-Sequencing Analysis of Mutations and Recombination in Pepper Progeny of Gamma-Irradiated Gametophytes. Plants, 10(1), 144. https://doi.org/10.3390/plants10010144