Application of Exogenous Phytohormones at Silking Stage Improve Grain Quality under Post-Silking Drought Stress in Waxy Maize



Abstract
:1. Introduction
2. Results and Discussion
2.1. Grain Weight
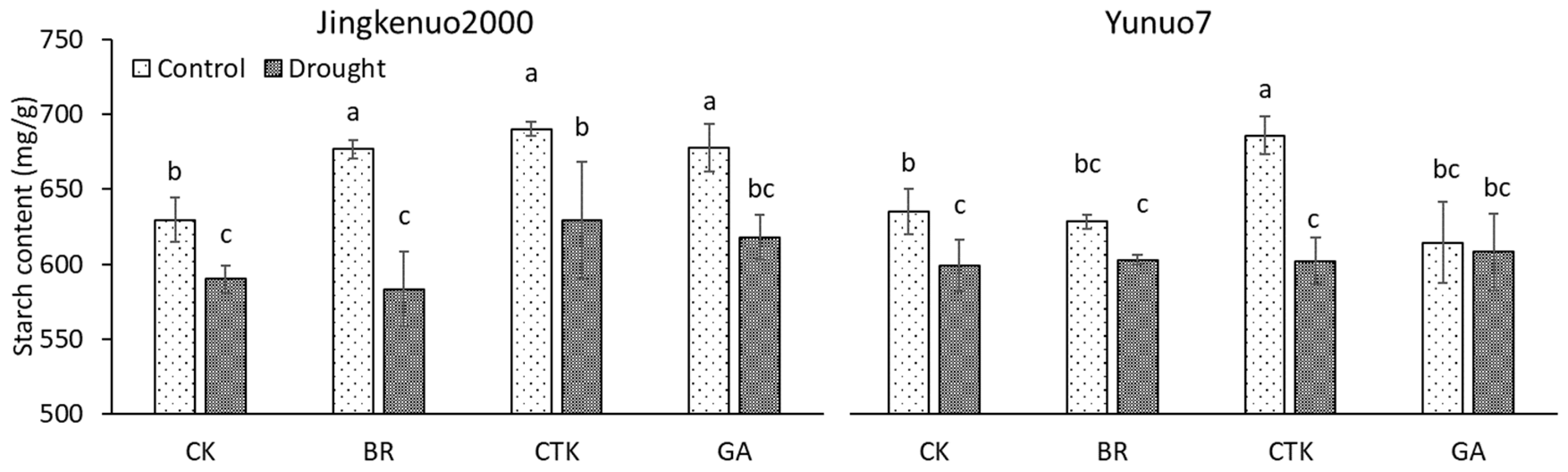
2.2. Starch Content
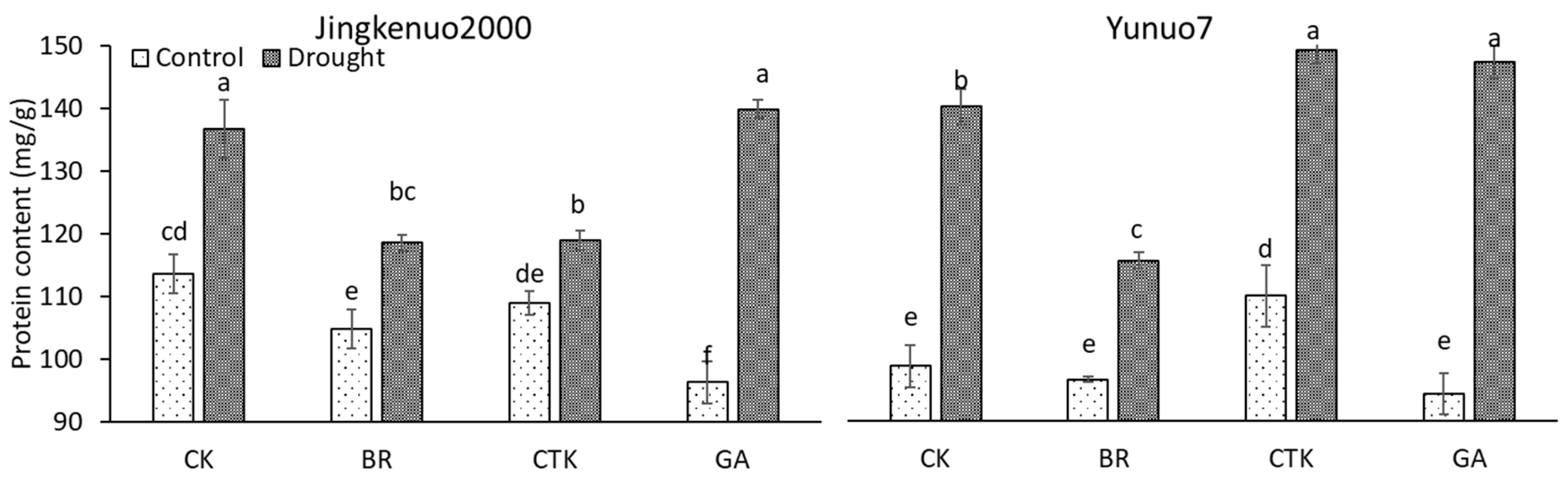
2.3. Protein Content
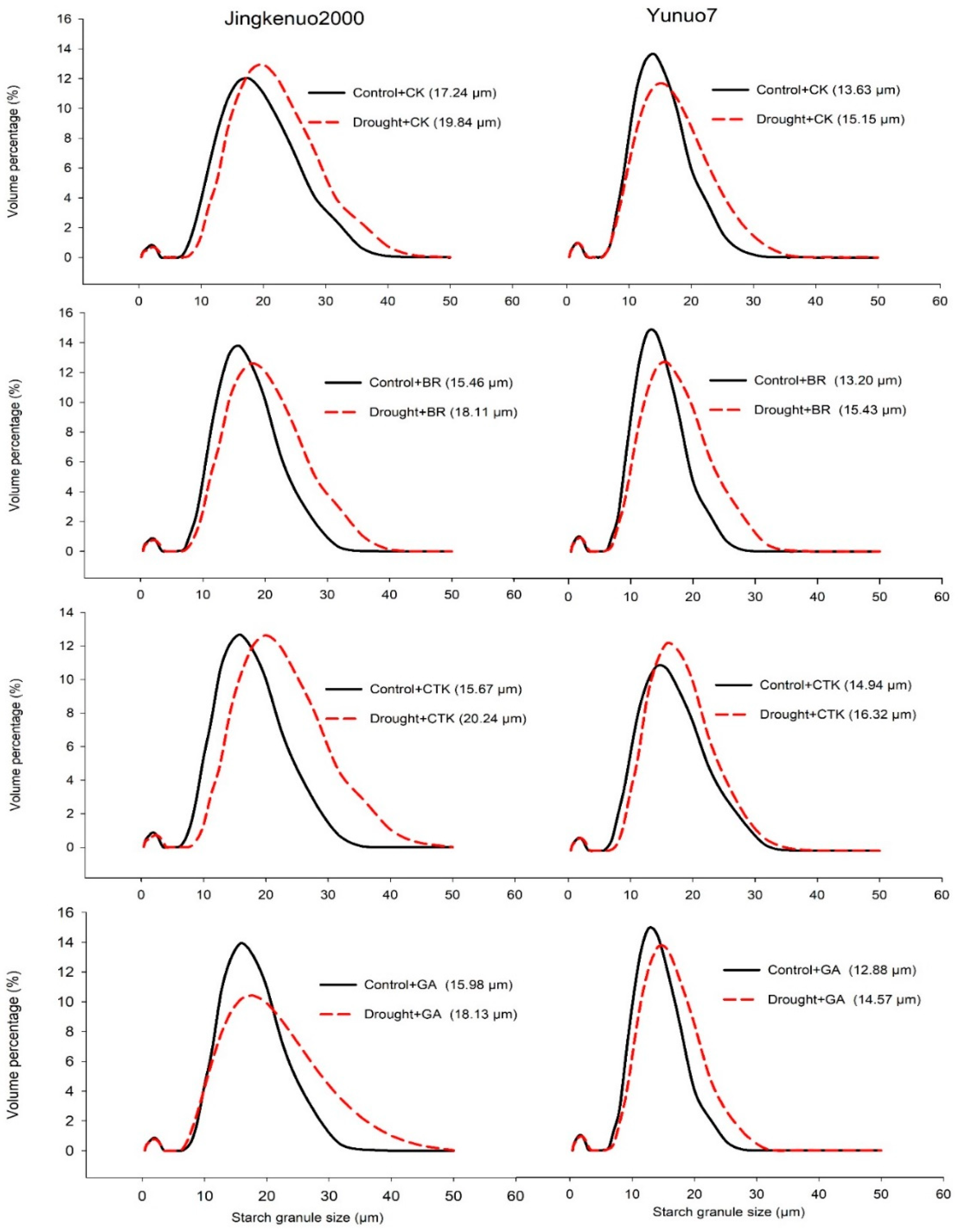
2.4. Starch Granule Size (SGS)
2.5. X-ray Diffraction (XRD) Pattern
2.6. Iodine Staining
2.7. Pasting Property
2.8. Thermal Property
3. Materials and Methods
3.1. Experimental Design
3.2. Grain Weight
3.3. Grain Starch and Protein Contents Determination
3.4. Starch Isolation
3.5. Granule Size Distribution
3.6. Iodine Staining
3.7. X-ray Diffraction Pattern
3.8. Pasting Properties
3.9. Thermal Properties
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boomsma, C.R.; Vyn, T.J. Maize drought tolerance: Potential improvements through arbuscular mycorrhizal symbiosis? Field Crop. Res. 2008, 108, 14–31. [Google Scholar] [CrossRef]
- Zaidi, P.H.; Rafique, S.; Singh, N.N. Response of maize (Zea mays L.) genotypes to excess soil moisture stress: Morpho-physiological effects and basis of tolerance. Eur. J. Agron. 2003, 19, 383–399. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.X.; Jacinthe, P.A. Global synthesis of drought effects on maize and wheat production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef] [PubMed]
- Daryanto, S.; Wang, L.X.; Jacinthe, P.A. Global synthesis of drought effects on cereal, legume, tuber and root crops production: A review. Agric. Water Manag. 2017, 179, 18–33. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.X.; Frei, M. Stressed food—The impact of abiotic environmental stresses on crop quality. Agric. Ecosyst. Environ. 2011, 141, 271–286. [Google Scholar] [CrossRef]
- Thitisaksakul, M.; Jimenez, R.C.; Arias, M.C.; Beckles, D.M. Effects of environmental factors on cereal starch biosynthesis and composition. J. Cereal Sci. 2012, 56, 67–80. [Google Scholar] [CrossRef]
- Beckles, D.M.; Thitisaksakul, M. How environmental stress affects starch composition and functionality in cereal endosperm. Starch Stärke 2014, 66, 58–71. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, M.; Gleadow, R.; Dalling, M. Effect of postanthesis drought on cell-division and starch accumulation in developing wheat grains. Ann. Bot. 1985, 55, 433–444. [Google Scholar] [CrossRef]
- Yang, W.B.; Li, Y.; Yin, Y.P.; Jiang, W.W.; Peng, D.L.; Cui, Z.Y.; Yang, D.Q.; Wang, Z.L. Ethylene and spermidine in wheat grains in relation to starch content and granule size distribution under water deficit. J. Integr. Agric. 2014, 13, 2141–2153. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.F.; Wang, C.Y.; Guo, T.C.; Xie, Y.X.; Feng, W.; Li, S.Y. Starch composition and its granules distribution in wheat grains in relation to post- anthesis high temperature and drought stress treatments. Starch Stärke 2014, 66, 419–428. [Google Scholar] [CrossRef]
- Yu, X.R.; Li, B.; Wang, L.L.; Chen, X.Y.; Wang, W.J.; Gu, Y.J.; Wang, Z.; Xiong, F. Effect of drought stress on the development of endosperm starch granules and the composition and physicochemical properties of starches from soft and hard wheat. J. Sci. Food Agric. 2016, 96, 2746–2754. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, C.Y.; Zhang, R.Q.; Liang, W.; Kang, X.L.; Jia, Y.; Liao, Y.C. Effects of drought on the morphological and physicochemical characteristics of starch granules in different elite wheat varieties. J. Cereal Sci. 2015, 66, 66–73. [Google Scholar] [CrossRef]
- Singh, S.; Singh, G.; Singh, P.; Singh, N. Effect of water stress at different stages of grain development on the characteristics of starch and protein of different wheat varieties. Food Chem. 2008, 108, 130–139. [Google Scholar] [CrossRef]
- Singh, S.; Gupta, A.K.; Gupta, S.K.; Kaur, N. Effect of sowing time on protein quality and starch pasting characteristics in wheat (Triticum aestivum L.) genotypes grown under irrigated and rain-fed conditions. Food Chem. 2010, 122, 559–565. [Google Scholar] [CrossRef]
- Xia, J.; Zhu, D.; Chang, H.M.; Yan, X.; Yan, Y.M. Effects of water-deficit and high-nitrogen treatments on wheat resistant starch crystalline structure and physicochemical properties. Carbohyd. Polym. 2020, 234, 115905. [Google Scholar] [CrossRef]
- Zhang, W.; Gu, J.; Wang, Z.; Wei, C.; Yang, J.; Zhang, J. Comparison of structural and functional properties of wheat starch under different soil drought conditions. Sci. Rep. 2017, 7, 12312. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, K.; Anand, K.G.V.; Vaghela, P.; Ghosh, A. Differential growth, yield and biochemical responses of maize to the exogenous application of Kappaphycus alvarezii seaweed extract, at grain-filling stage under normal and drought conditions. Algal. Res. 2018, 35, 236–244. [Google Scholar] [CrossRef]
- Ullah, A.; Manghwar, H.; Shaban, M.; Khan, A.H.; Akbar, A.; Ali, U.; Ali, E.; Fahad, S. Phytohormones enhanced drought tolerance in plants: A coping strategy. Environ. Sci. Pollut. R. 2018, 25, 33103–33118. [Google Scholar] [CrossRef]
- Anjum, S.A.; Wang, L.C.; Farooq, M.; Hussain, M.; Xue, L.L.; Zou, C.M. Brassinolide application improves the drought tolerance in maize through modulation of enzymatic antioxidants and leaf gas exchange. J. Agron. Crop Sci. 2011, 197, 177–185. [Google Scholar] [CrossRef]
- Kaya, C.; Tuna, A.L.; Alves, A.A.C. Gibberellic acid improves water deficit tolerance in maize plants. Acta Physiol. Plant 2006, 28, 331–337. [Google Scholar] [CrossRef]
- Xie, T.L.; Gu, W.R.; Li, C.F.; Li, J.; Wei, S. Exogenous DCPTA increases the tolerance of maize seedlings to PEG-simulated drought by regulating nitrogen metabolism-related enzymes. Agron. Basel 2019, 9, 676. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.X.; Guo, B.W.; Tang, J.; Xu, F.F.; Xu, K.; Hu, Y.J.; Xing, Z.P.; Zhang, H.C.; Dai, Q.G.; Huo, Z.Y.; et al. Combined effects of low temperature and weak light at grain-filling stage on rice grain quality. Acta Agron. Sin. 2019, 45, 1208–1220. [Google Scholar]
- Pei, L.M.; Che, R.H.; He, L.L.; Gao, X.X.; Li, W.J.; Li, H. Role of exogenous glutathione in alleviating abiotic stress in maize (Zea mays L.). J. Plant Growth Regul. 2019, 38, 199–215. [Google Scholar] [CrossRef]
- Li, L.J.; Gu, W.R.; Li, C.F.; Li, W.H.; Li, C.F.; Li, J.; Wei, S. Exogenous spermidine improves drought tolerance in maize by enhancing the antioxidant defence system and regulating endogenous polyamine metabolism. Crop. Pasture Sci. 2018, 69, 1076–1091. [Google Scholar] [CrossRef]
- Maswada, H.F.; Abd El-Razek, U.A.; El-Sheshtawy, A.N.A.; Elzaawely, A.A. Morpho-physiological and yield responses to exogenous moringa leaf extract and salicylic acid in maize (Zea mays L.) under water stress. Arch. Agron. Soil Sci. 2018, 64, 994–1010. [Google Scholar] [CrossRef]
- McMillan, T.; Tidemann, B.D.; O’Donovan, J.T.; Izydorczyk, M.S. Effects of plant growth regulator application on the malting quality of barley. J. Sci. Food Agric. 2020, 100, 2082–2089. [Google Scholar] [CrossRef]
- Xu, Y.J.; Qiu, M.T.; Li, Y.Y.; Qian, X.Y.; Gu, J.F.; Yang, J.C. Polyamines mediate the effect of post-anthesis soil drying on starch granule size distribution in wheat kernels. Crop J. 2016, 4, 444–458. [Google Scholar] [CrossRef] [Green Version]
- Pan, S.G.; Rasul, F.; Li, W.; Tian, H.; Mo, Z.W.; Duan, M.Y.; Tang, X.R. Roles of plant growth regulators on yield, grain qualities and antioxidant enzyme activities in super hybrid rice (Oryza sativa L.). Rice 2013, 6, 1–10. [Google Scholar] [CrossRef]
- Lu, D.L.; Lu, W.P. Effects of protein removal on the physicochemical properties of waxy maize flours. Starch Stärke 2012, 64, 874–881. [Google Scholar] [CrossRef]
- Hsieh, C.F.; Liu, W.C.; Whaley, J.K.; Shi, Y.C. Structure and functional properties of waxy starches. Food Hydrocolloid. 2019, 94, 238–254. [Google Scholar] [CrossRef]
- Yang, H.; Gu, X.T.; Ding, M.Q.; Lu, W.P.; Lu, D.L. Activities of starch synthetic enzymes and contents of endogenous hormones in waxy maize grains subjected to post-silking water deficit. Sci. Rep. 2019, 9, 7059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, D.L.; Cai, X.M.; Zhao, J.Y.; Shen, X.; Lu, W.P. Effects of drought after pollination on grain yield and quality of fresh waxy maize. J. Sci. Food Agric. 2015, 95, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.L.; Sun, S.L.; Wang, X.; Yan, F.B.; Lu, W.P. Effects of water stress during grain filling on physicochemical properties of waxy maize flour. Sci. Agric. Sin. 2013, 46, 30–36. [Google Scholar]
- El-Sabagh, A.; Barutcular, C.; Hossain, A.; Islam, M.S. Response of maize hybrids to drought tolerance in relation to grain weight. Fresen Environ. Bull. 2018, 27, 2476–2482. [Google Scholar]
- Rajala, A.; Hakala, K.; Makela, P.; Peltonen-Sainio, P. Drought effect on grain number and grain weight at spike and spikelet level in six-row spring barley. J. Agron. Crop. Sci. 2011, 197, 103–112. [Google Scholar] [CrossRef]
- Ahmad, I.; Kamran, M.; Meng, X.P.; Ali, S.; Bilegjargal, B.; Cai, T.; Liu, T.N.; Han, Q.F. Effects of plant growth regulators on seed filling, endogenous hormone contents and maize production in semiarid regions. J. Plant Growth Regul. 2019, 38, 1467–1480. [Google Scholar] [CrossRef]
- Panda, B.B.; Sekhar, S.; Dash, S.K.; Behera, L.; Shaw, B.P. Biochemical and molecular characterisation of exogenous cytokinin application on grain filling in rice. BMC Plant Biol. 2018, 18, 89. [Google Scholar] [CrossRef] [Green Version]
- Javid, M.G.; Sorooshzadeh, A.; Sanavy, S.A.M.M.; Allahdadi, I.; Moradi, F. Effects of the exogenous application of auxin and cytokinin on carbohydrate accumulation in grains of rice under salt stress. Plant Growth Regul. 2011, 65, 305–313. [Google Scholar] [CrossRef]
- Xie, Z.J.; Jiang, D.; Dai, T.B.; Jing, Q.; Cao, W.X. Effects of exogenous ABA and cytokinin on leaf photosynthesis and grain protein accumulation in wheat ears cultured in vitro. Plant Growth Regul. 2004, 44, 25–32. [Google Scholar] [CrossRef]
- Prathap, V.; Ali, K.; Singh, A.; Vishwakarma, C.; Krishnan, V.; Chinnusamy, V.; Tyagi, A. Starch accumulation in rice grains subjected to drought during grain filling stage. Plant Physiol. Bioch. 2019, 142, 440–451. [Google Scholar]
- Zhang, Q.; Zhang, L.Z.; Evers, J.C.; van der Werf, W.; Zhang, W.Q.; Duan, L.S. Maize yield and quality in response to plant density and application of a novel plant growth regulator. Field Crop. Res. 2014, 164, 82–89. [Google Scholar] [CrossRef]
- Chen, X.Y.; Li, B.; Shao, S.S.; Wang, L.L.; Zhu, X.W.; Yang, Y.; Wang, W.J.; Yu, X.R.; Xiong, F. Accumulation characteristic of protein bodies in different regions of wheat endosperm under drought stress. J. Integr. Agric. 2016, 15, 2921–2930. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, Z.; Yin, Y.; Cai, R.; Yan, S.; Li, W. Starch content and granule size distribution in grains of wheat in relation to post-anthesis water deficits. J. Agron. Crop Sci. 2010, 196, 1–8. [Google Scholar] [CrossRef]
- Li, L.; Blanco, M.; Jane, J. Physicochemical properties of endosperm and pericarp starches during maize development. Carbohyd. Polym. 2007, 67, 630–639. [Google Scholar] [CrossRef]
- Peng, D.L.; Cai, T.; Yin, Y.P.; Yang, W.B.; Ni, Y.L.; Yang, D.Q.; Wang, Z.L. Exogenous application of abscisic acid or gibberellin acid has different effects on starch granule size distribution in grains of wheat. J. Integr. Agric. 2013, 12, 1551–1559. [Google Scholar] [CrossRef]
- Yang, D.Q.; Li, Y.; Shi, Y.H.; Cui, Z.Y.; Luo, Y.L.; Zheng, M.J.; Chen, J.; Li, Y.X.; Yin, Y.P.; Wang, Z.L. Exogenous cytokinins increase grain yield of winter wheat cultivars by improving stay-green characteristics under heat stress. PLoS ONE 2016, 11, e0155437. [Google Scholar] [CrossRef]
- Basunia, M.A.; Nonhebel, H.M. Hormonal regulation of cereal endosperm development with a focus on rice (Oryza sativa). Funct. Plant Biol. 2019, 46, 493–506. [Google Scholar] [CrossRef]
- Almeida, V.O.; Batista, K.A.; Di-Medeiros, M.C.B.; Moraes, M.G.; Fernandes, K.F. Effect of drought stress on the morphological and physicochemical properties of starches from Trimezia juncifolia. Carbohyd. Polym. 2019, 212, 304–311. [Google Scholar] [CrossRef]
- Bertofta, E.; Annor, G.A.; Shen, X.Y.; Rumpagaporn, P.; Seetharaman, K.; Hamaker, B.R. Small differences in amylopectin fine structure may explain large functional differences of starch. Carbohyd. Polym. 2016, 140, 113–121. [Google Scholar] [CrossRef]
- Lu, D.L.; Sun, X.L.; Yan, F.B.; Wang, X.; Xu, R.C.; Lu, W.P. Effects of high temperature during grain filling under control conditions on the physicochemical properties of waxy maize flour. Carbohyd. Polym. 2013, 98, 302–310. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Saud, S.; Hassan, S.; Chauhan, B.S.; Khan, F.; Ihsan, M.Z.; Ullah, A.; Wu, C.; Bajwa, A.A.; et al. Responses of rapid viscoanalyzer profile and other rice grain qualities to exogenously applied plant growth regulators under high day and high night temperatures. PLoS ONE 2016, 11, e0159590. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.Z.; Hu, J.; Zhang, J.W.; Dong, S.T.; Liu, P.; Zhao, B. Spraying exogenous synthetic cytokinin 6-benzyladenine following the waterlogging improves grain growth of waterlogged maize in the field. J. Agron. Crop. Sci. 2019, 205, 616–624. [Google Scholar] [CrossRef]
- Gao, Z.; Liang, X.G.; Zhang, L.; Lin, S.; Zhao, X.; Zhou, L.L.; Shen, S.; Zhou, S.L. Spraying exogenous 6-benzyladenine and brassinolide at maize yield by enhancing source and sink capacity tasseling increases. Field Crop. Res. 2017, 211, 1–9. [Google Scholar] [CrossRef]
- Hansen, J.; Moller, I. Percolation of starch and soluble carbohydrates from plant tissue for quantitative determination with anthrone. Anal. Biochem. 1975, 68, 87–94. [Google Scholar] [CrossRef]
- Chemists, A.A.O.C. Approved Methods of the AACCI; The Association: St. Paul, MN, USA, 1999. [Google Scholar]
- Gu, X.T.; Huang, T.Q.; Ding, M.Q.; Lu, W.P.; Lu, D.L. Effects of short-term heat stress at the grain formation stage on physicochemical properties of waxy maize starch. J. Sci. Food Agric. 2018, 98, 1008–1015. [Google Scholar] [CrossRef]
- Tang, Q.Y.; Feng, M.G. Data Processing System: Experimental Design, Statistical Analysis and Data Mining; Science Press: Beijing, China, 2007. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hybrid | Soil Moisture | Exogenous Hormones | PV (cP) | TV (cP) | BD (cP) | FV (cP) | SB (cP) | Ptemp (°C) |
|---|---|---|---|---|---|---|---|---|
| JKN2000 | Control | CK | 413 c | 234 b | 179 c | 353 b | 119 cd | 76.8 c |
| BR | 701 a | 308 a | 389 a | 434 a | 123 bcd | 74.9 d | ||
| CTK | 418 c | 237 b | 182 c | 351 b | 114 d | 76.7 c | ||
| GA | 498 b | 204 b | 291 b | 320 b | 114 d | 75.1 d | ||
| Drought | CK | 246 d | 235 b | 85 de | 347 b | 138 a | 79.3 a | |
| BR | 402 c | 291 a | 138 cd | 421 a | 131 abc | 76.9 bc | ||
| CTK | 282 d | 212 b | 71 e | 332 b | 120 cd | 77.0 bc | ||
| GA | 381 c | 293 a | 91 de | 421 a | 135 ab | 78.3 ab | ||
| YN7 | Control | CK | 1516 b | 731 ef | 771 b | 913 c | 177 cd | 76.7 cd |
| BR | 1650 a | 811 c | 839 a | 1009 b | 198 a | 75.9 d | ||
| CTK | 1473 b | 880 ab | 593 c | 1062 a | 182 bc | 78.3 b | ||
| GA | 1512 b | 709 f | 803 ab | 883 c | 174 cd | 76.2 d | ||
| Drought | CK | 979 d | 770 de | 209 e | 962 b | 191 ab | 79.4 a | |
| BR | 1492 b | 882 a | 610 c | 1063 a | 181 bc | 77.5 bc | ||
| CTK | 981 d | 801 cd | 179 e | 999 b | 198 a | 79.7 a | ||
| GA | 1354 c | 841 bc | 513 d | 1007 b | 167 d | 78.3 b | ||
| F-value | ||||||||
| Hybrid | 6690.3 ** | 6865.8** | 2145.3 ** | 6025.3 ** | 722.3 ** | 19.1 ** | ||
| Soil moisture | 492.0 ** | 15.6 ** | 1037.9 ** | 13.0 ** | 11.5 ** | 95.4 ** | ||
| Hormone | 128.1 ** | 26.7 ** | 166.7 ** | 24.0 ** | 4.7 ** | 16.5 ** | ||
| Hybrid × Soil moisture | 45.3 ** | 4.7 ** | 157.7 ** | 2.5 | 7.1 ** | 0.0 | ||
| Hybrid × Hormone | 8.1 ** | 11.5 ** | 34.6 ** | 10.6 ** | 6.5 ** | 5.2 ** | ||
| Soil moisture × Hormone | 16.8 ** | 24.8 ** | 6.0 ** | 15.9 ** | 4.1 * | 4.5 * | ||
| Hybrid × Soil moisture × Hormone | 28.9 ** | 5.0 ** | 43.0 ** | 2.4 | 4.0 * | 1.3 | ||
| Hybrid | Soil Moisture | Exogenous Hormones | ΔHgel (J/g) | To (°C) | Tp (°C) | Tc (°C) | ΔHret (J/g) | %R (%) |
|---|---|---|---|---|---|---|---|---|
| JKN2000 | Control | CK | 9.2 a | 68.7 e | 73.9 e | 83.1 c | 3.6 a | 39.1 a |
| BR | 9.3 a | 69.5 cd | 74.8 d | 83.9 b | 3.2 b | 33.9 b | ||
| CTK | 9.0 a | 69.4 d | 74.7 d | 84.1 b | 2.1 e | 23.2 d | ||
| GA | 8.1 b | 69.9 c | 75.4 c | 84.4 b | 2.1 de | 26.3 c | ||
| Drought | CK | 7.3 c | 70.8 b | 76.1 b | 84.4 b | 2.5 c | 34.6 b | |
| BR | 7.0 c | 68.8 e | 74.7 d | 82.4 d | 2.3 de | 32.4 b | ||
| CTK | 6.9 c | 69.7 cd | 75.5 c | 84.3 b | 2.2 de | 32.3 b | ||
| GA | 7.1 c | 72.0 a | 77.1 a | 85.7 a | 2.3 cd | 32.8 b | ||
| YN7 | Control | CK | 7.4 bc | 68.1 f | 73.4 f | 81.5 d | 3.2 b | 42.9 d |
| BR | 7.8 ab | 68.6 e | 74.5 e | 82.5 c | 3.1 bc | 39.5 e | ||
| CTK | 7.8 ab | 69.6 d | 75.0 d | 82.4 c | 3.0 bc | 39.0 e | ||
| GA | 7.7 b | 68.8 e | 73.7 f | 82.1 cd | 4.0 a | 51.8 a | ||
| Drought | CK | 8.2 a | 71.4 b | 76.9 ab | 84.2 a | 4.1 a | 49.4 b | |
| BR | 7.8 ab | 70.1 c | 76.1 c | 83.4 b | 3.8 a | 48.4 b | ||
| CTK | 7.4 bc | 71.8 a | 77.2 a | 84.6 a | 2.8 c | 37.1 f | ||
| GA | 7.1 c | 71.1 b | 76.5 b | 84.8 a | 3.3 b | 46.0 c | ||
| F-value | ||||||||
| Hybrid | 15.0 ** | 1.3 | 5.6 * | 71.5 ** | 307.9 ** | 1332.4 ** | ||
| Soil moisture | 105.3 ** | 639.4 ** | 1052.2 ** | 159.9 ** | 7.2 * | 39.7 ** | ||
| Hormone | 7.3 ** | 62.2 ** | 39.9 ** | 31.5 ** | 49.2 ** | 115.4 ** | ||
| Hybrid × Soil moisture | 101.3 ** | 101.3 ** | 151.1 ** | 85.1 ** | 34.1 ** | 0.4 | ||
| Hybrid × Hormone | 0.3 | 48.7 ** | 65.5 ** | 7.9 ** | 14.8 ** | 46.4 ** | ||
| Soil moisture × Hormone | 2.9 | 64.3 ** | 69.0 ** | 33.0 ** | 0.9 | 6.6 ** | ||
| Hybrid × Soil moisture × Hormone | 7.2 ** | 11.8 ** | 1.3 | 1.6 | 54.9 ** | 89.9 ** | ||
| PC | SC | SGS | RC | λmax | ΔHgel | To | Tp | Tc | ΔHret | %R | PV | TV | BD | FV | SB | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SC | −0.56 * | |||||||||||||||
| SGS | 0.43 | −0.25 | ||||||||||||||
| RC | 0.11 | −0.08 | 0.02 | |||||||||||||
| λmax | 0.32 | −0.32 | 0.83 ** | 0.05 | ||||||||||||
| ΔHgel | 0.39 | −0.20 | −0.26 | 0.01 | −0.43 | |||||||||||
| To | −0.38 | 0.58 * | 0.32 | −0.21 | 0.34 | −0.28 | ||||||||||
| Tp | 0.86 ** | −0.31 | 0.36 | 0.25 | 0.31 | −0.34 | 0.97 ** | |||||||||
| Tc | 0.85 ** | −0.36 | 0.51 * | 0.18 | 0.46 | −0.07 | 0.85 ** | 0.82 ** | ||||||||
| ΔHret | 0.69 ** | −0.06 | −0.59 * | 0.33 | −0.52 * | 0.25 | −0.09 | −0.13 | −0.37 | |||||||
| %R | −0.04 | −0.27 | −0.52 * | −0.34 | −0.36 | −0.16 | 0.01 | 0.00 | −0.35 | 0.91 ** | ||||||
| PV | 0.10 | −0.49 * | −0.85 ** | −0.26 | −0.60 * | −0.11 | −0.22 | −0.19 | −0.54 * | 0.65 ** | 0.73 ** | |||||
| TV | −0.25 | −0.03 | −0.70 ** | −0.15 | −0.51 * | −0.22 | 0.08 | 0.12 | −0.34 | 0.65 ** | 0.76 ** | 0.92 ** | ||||
| BD | 0.10 | −0.21 | −0.86 ** | −0.18 | −0.57 * | 0.00 | −0.49 * | −0.49 * | −0.67 ** | 0.54 * | 0.58 * | 0.92 ** | 0.70 ** | |||
| FV | −0.56 * | 0.13 | −0.70 ** | −0.10 | −0.52 * | −0.22 | 0.08 | 0.12 | −0.34 | 0.65 ** | 0.76 ** | 0.92 ** | 1.00 ** | 0.70 ** | ||
| SB | 0.10 | −0.21 | −0.60 * | −0.18 | −0.43 | −0.29 | 0.14 | 0.18 | −0.31 | 0.60 * | 0.73 ** | 0.84 ** | 0.95 ** | 0.61 ** | 0.95 ** | |
| Ptemp | 0.86 ** | −0.54 * | 0.29 | −0.06 | 0.34 | −0.43 | 0.72 ** | 0.70 ** | 0.37 | 0.10 | 0.27 | −0.01 | 0.33 | −0.34 | 0.33 | 0.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Yan, Y.; Lu, W.; Lu, D. Application of Exogenous Phytohormones at Silking Stage Improve Grain Quality under Post-Silking Drought Stress in Waxy Maize. Plants 2021, 10, 48. https://doi.org/10.3390/plants10010048
Wang L, Yan Y, Lu W, Lu D. Application of Exogenous Phytohormones at Silking Stage Improve Grain Quality under Post-Silking Drought Stress in Waxy Maize. Plants. 2021; 10(1):48. https://doi.org/10.3390/plants10010048
Chicago/Turabian StyleWang, Longfei, Yini Yan, Weiping Lu, and Dalei Lu. 2021. "Application of Exogenous Phytohormones at Silking Stage Improve Grain Quality under Post-Silking Drought Stress in Waxy Maize" Plants 10, no. 1: 48. https://doi.org/10.3390/plants10010048
APA StyleWang, L., Yan, Y., Lu, W., & Lu, D. (2021). Application of Exogenous Phytohormones at Silking Stage Improve Grain Quality under Post-Silking Drought Stress in Waxy Maize. Plants, 10(1), 48. https://doi.org/10.3390/plants10010048