Single and Combined Abiotic Stress in Maize Root Morphology




Abstract
:1. Introduction
2. Results
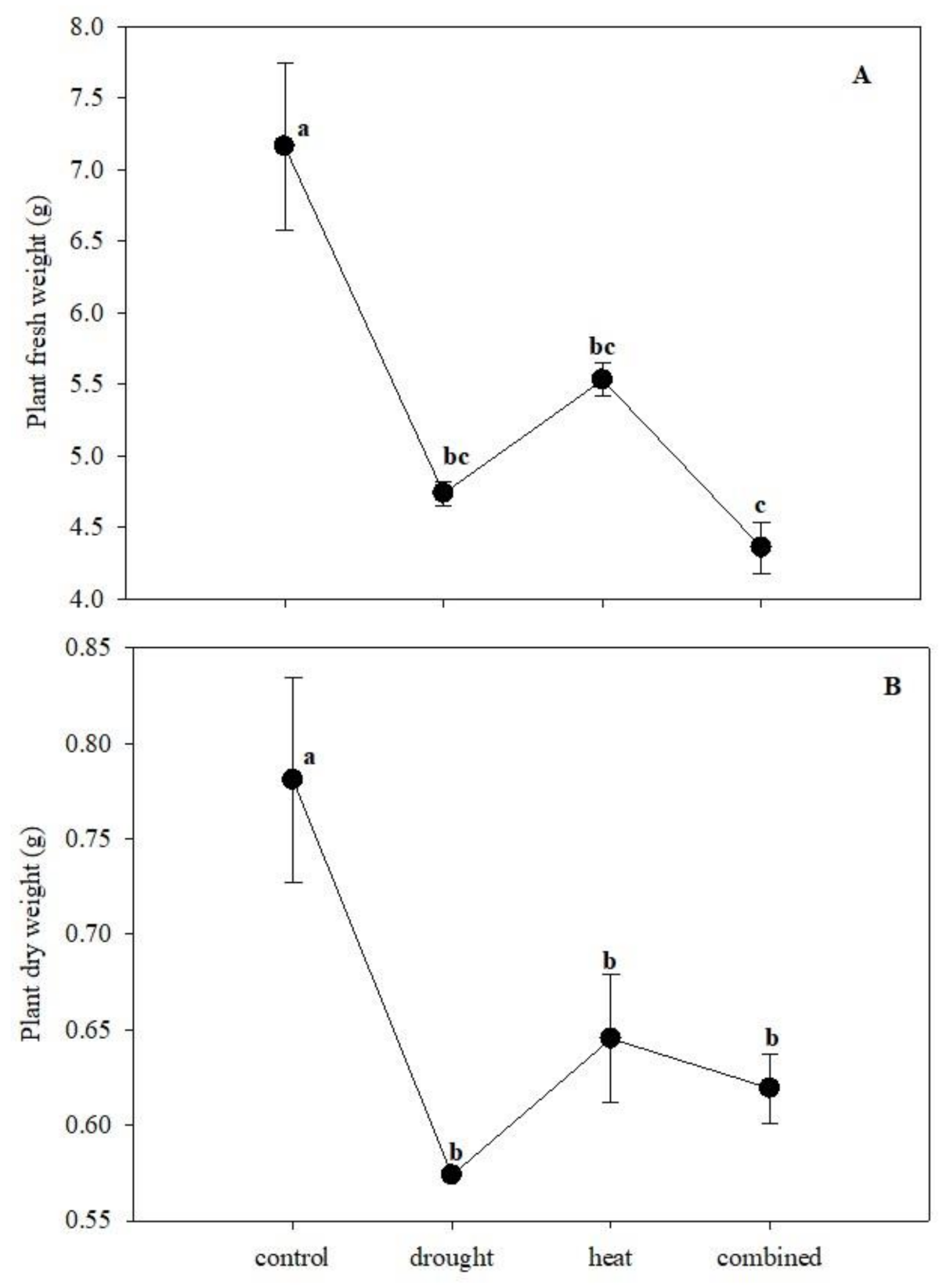
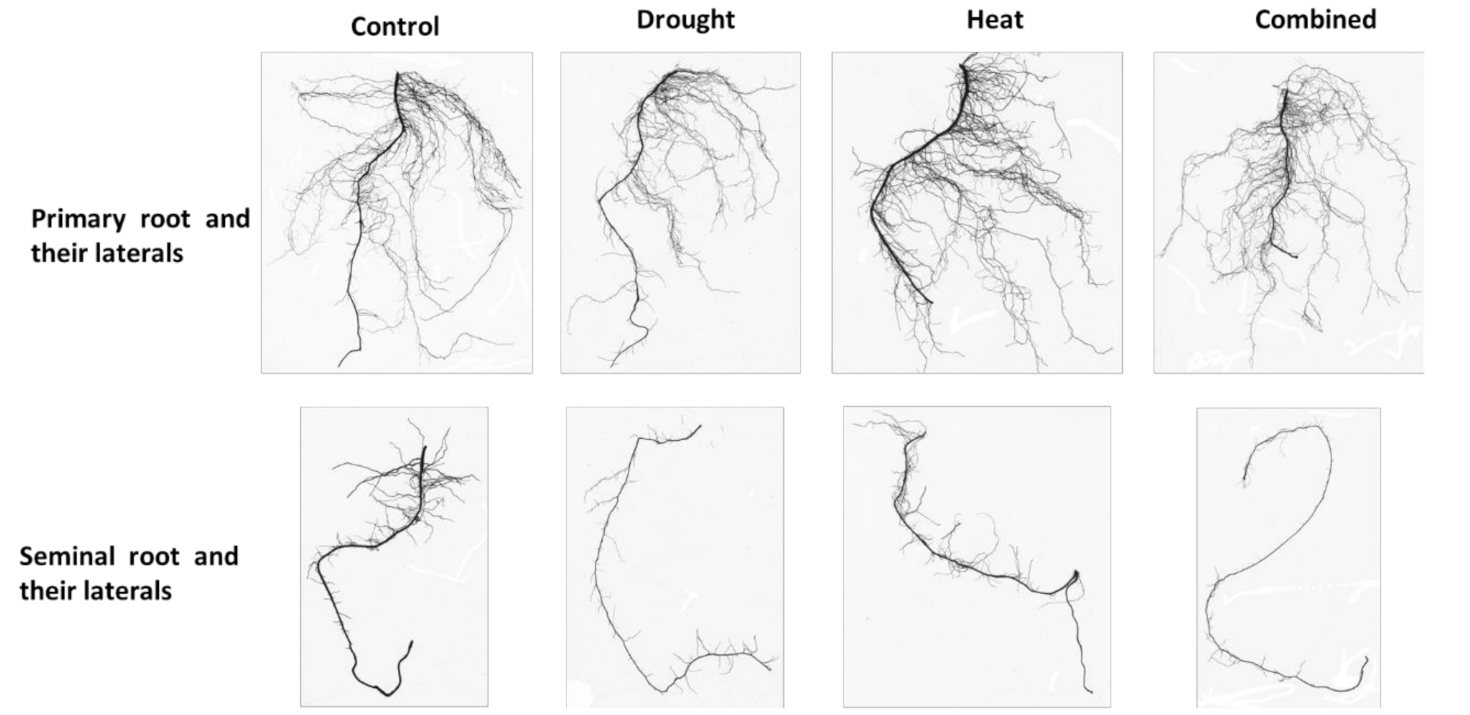
2.1. Univariate Analysis of the Root Morphological Data
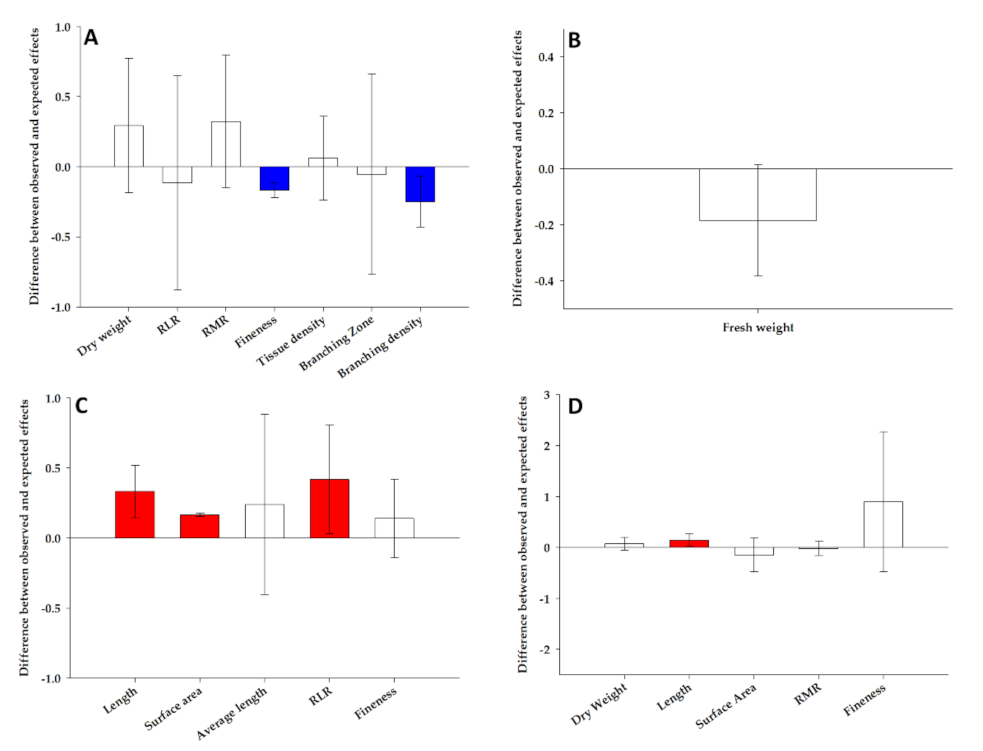
2.2. Additive, Synergistic and Antagonistic Effect of Combined Stress
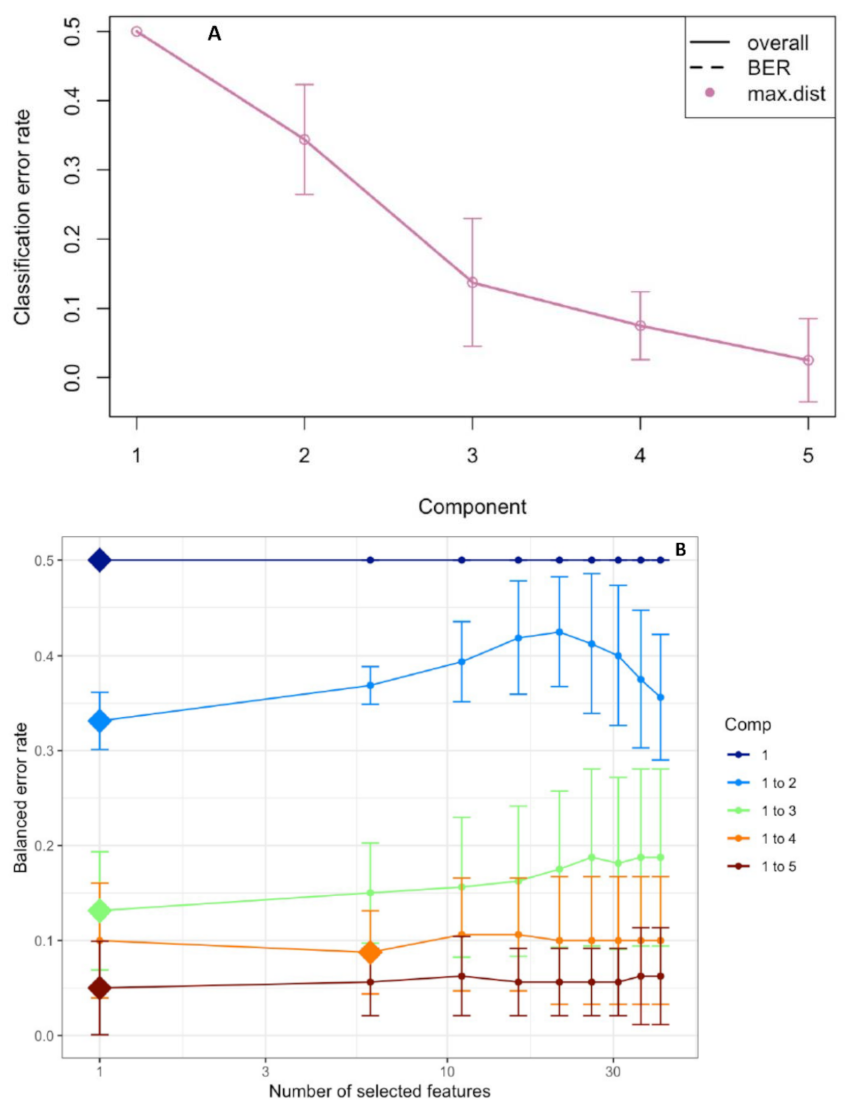
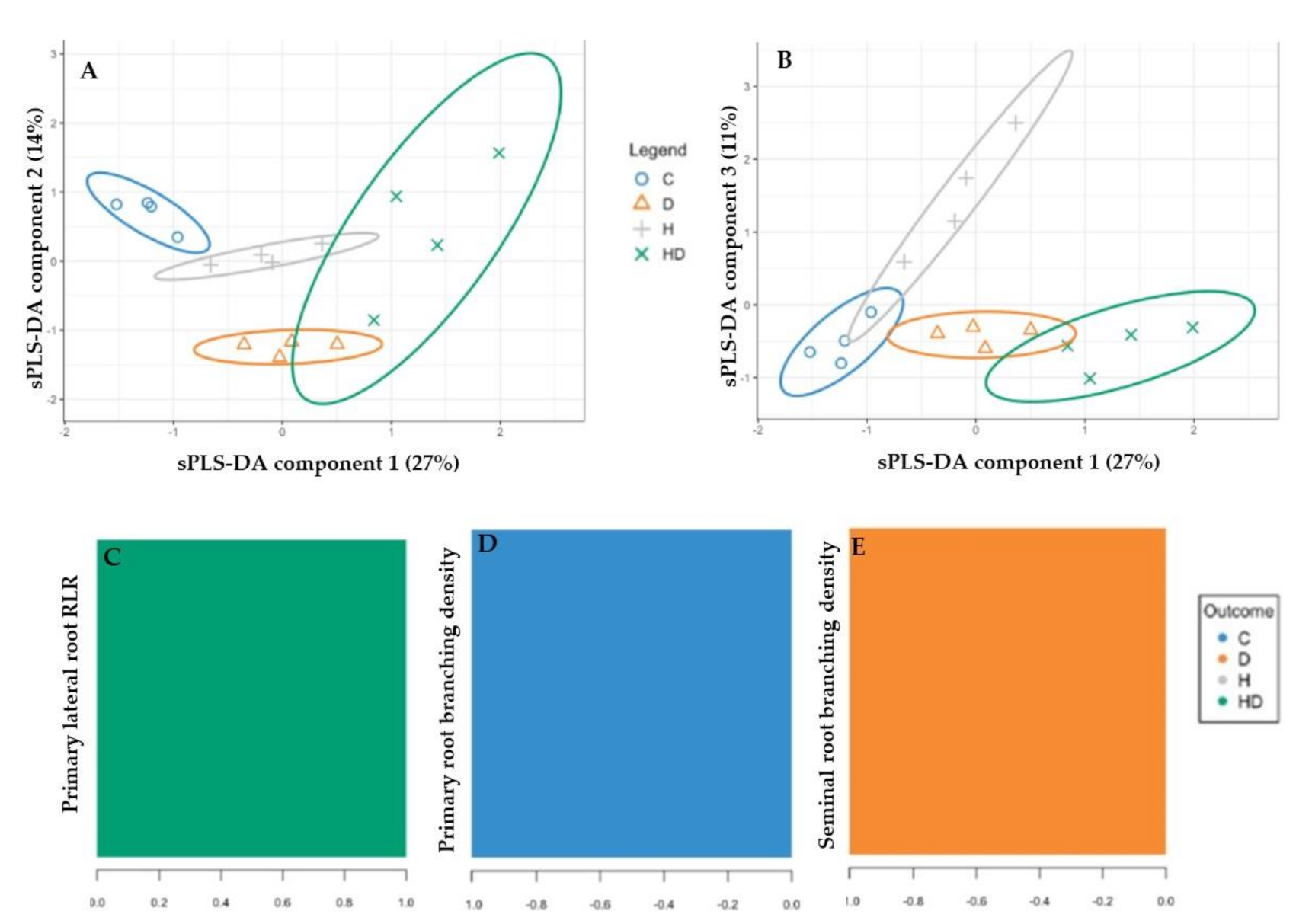
2.3. Root Responses to the Single and Combined Stress: A Supervised Analysis with PLS-DA
3. Discussion
3.1. Single Stress Determined Different Root Type-Related Morphological Responses
3.2. Combined Stress Caused Different Root Type-Related Morphological Response with Respect to the Single Stress with Nonadditive Effects
3.3. Primary Lateral RLR and Primary and Seminal Root Branching Density Discriminated the Root Phenotypes in Drought and Heat Stress and Their Combination
4. Materials and Methods
4.1. Plant Material and Growth Condition and Treatment
4.2. Treatments
4.3. Morphological Root Analysis
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| sPLS-DA | Partial Least Squares-Discriminant Analysis |
| PCA | Principal Component Analysis |
| D | Drought stress |
| H | Heat stress |
| Comb | Combined stress |
| FC | Field Capacity |
| Con | Control |
| ShFW | Shoot Fresh Weight |
| ShDW | Shoot Dry Weight |
| L | Length |
| SA | Surface Area |
| V | Volume |
| S | Seminal root |
| Pr | Primary root |
| SL | Seminal lateral root |
| PL | Primary Lateral roots |
| RZF | Root Zone Formation |
| BZF | Branching Zone Formation |
| BD | Branching Density |
| PrFW | Primary Fresh Weight |
| PrDW | Primary Dry Weight |
| SFW | Seminal Fresh weight |
| SDW | Seminal Dry Weight |
| PrLFW | Primary Lateral Fresh Weight |
| PrLDW | Primary Lateral Dry Weight |
| SLFW | Seminal Lateral Fresh Weight |
| SLDW | Seminal Lateral Dry Weight |
| RFW | Root Fresh Weight |
| RDW | Root Dry Weight |
| PFW | Plant Fresh Weight |
| PDW | Plant Dry Weight |
| RLR | Root Length Ratio |
| RMR | Root Mass Ratio |
| RF | Root Fineness |
| RTD | Root Tissue Density |
| FDR | False Discovery Rate |
References
- Cardell, M.F.; Amengual, A.; Romero, R.; Ramis, C. Future extremes of temperature and precipitation in Europe derived from a combination of dynamical and statistical approaches. Int. J. Climatol. 2020, 40, 4800–4827. [Google Scholar] [CrossRef]
- Boisvenue, C.; Running, S.W. Impacts of climate change on natural forest productivity—Evidence since the middle of the 20th century. Glob. Chang. Biol. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Niinemets, U. Responses of forest trees to single and multiple environmental stresses from seedlings to mature plants: Past stress history, stress interactions, tolerance and acclimation. For. Ecol. Manag. 2010, 260, 1623–1639. [Google Scholar] [CrossRef]
- Lobell, D.B.; Gourdji, S.M. The influence of climate change on global crop productivity. Plant Physiol. 2012, 160, 1686–1697. [Google Scholar] [CrossRef] [Green Version]
- Tito, R.; Vasconcelos, H.L.; Feeley, K.J. Global climate change increases risk of crop yield losses and food insecurity in the tropical Andes. Glob. Chang. Biol. 2018, 24, e592–e602. [Google Scholar] [CrossRef]
- Sultan, B.; Defrance, D.; Iizumi, T. Evidence of crop production losses in West Africa due to historical global warming in two crop models. Sci. Rep. 2019, 9, 12834. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Liang, S.; Peng, S. Climate change affects forest productivity in a typical climate transition region of China. Sustainability 2019, 11, 2856. [Google Scholar] [CrossRef] [Green Version]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Wu, L.; Zhao, F.; Zhang, D.; Li, N.; Zhu, G.; Li, C.; Wang, W. Phosphoproteomic analysis of the response of maize leaves to drought, heat and their combination stress. Front. Plant Sci. 2015, 6, 298. [Google Scholar] [CrossRef] [Green Version]
- Sah, R.P.; Chakraborty, M.; Prasad, K. Impact of water deficit stress in maize: Phenology and yield components. Sci. Rep. 2020, 10, 2944. [Google Scholar] [CrossRef]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Rivero, R.M.; Mestre, T.C.; Mittler, R.; Rubio, F.; Garcia-Sanchez, F.; Martinez, V. The combined effect of salinity and heat reveals a specific physiological, biochemical and molecular response in tomato plants. Plant Cell Environ. 2013, 37, 1059–1073. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Ramegowda, V.; Senthil-Kumar, M. Shared and unique responses of plants to multiple individual stresses and stress combinations: Physiological and molecular mechanisms. Front. Plant Sci. 2015, 6, 723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, H.A.; Men, S.; Hussain, S. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obata, T.; Witt, S.; Lisec, J.; Palacios-Rojas, N.; Florez-Sarasa, I.; Yousfi, S.; Araus, J.L.; Cairns, J.E.; Fernie Alisdair, R. Metabolite profiles of maize leaves in drought, heat, and combined stress field trials reveal the relationship between metabolism and grain yield. Plant Physiol. 2015, 169, 2665–2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killi, D.; Bussotti, F.; Raschi, A.; Haworth, M. Adaptation to high temperature mitigates the impact of water deficit during combined heat and drought stress in C3 sunflower and C4 maize varieties with contrasting drought tolerance. Physiol. Plant. 2017, 159, 130–147. [Google Scholar] [CrossRef]
- Zhao, F.; Zhang, D.; Zhao, Y.; Wang, W.; Yang, H.; Tai, F.; Li, C.; Hu, X. The difference of physiological and proteomic changes in maize leaves adaptation to drought, heat, and combined both stresses. Front. Plant Sci. 2016, 7, 1471. [Google Scholar] [CrossRef] [Green Version]
- Hochholdinger, F.; Yu, P.; Marcon, C. Genetic control of root system development in maize. Trends Plant Sci. 2018, 23, 79–88. [Google Scholar] [CrossRef]
- Zhan, A.; Schneider, H.; Lynch, J.P. Reduced lateral root branching density improves drought tolerance in maize. Plant Physiol. 2015, 168, 1603–1615. [Google Scholar] [CrossRef] [Green Version]
- Hund, A.; Fracheboud, Y.; Soldati, A.; Stamp, P. Cold tolerance of maize seedlings as determined by root morphology and photosynthetic traits. Eur. J. Agron. 2008, 28, 178–185. [Google Scholar] [CrossRef]
- Abenavoli, M.R.; Sorgonà, A.; Albano, S.; Cacco, G. Coumarin differentially affects the morphology of different root types of maize seedlings. J. Chem. Ecol. 2004, 30, 1871–1883. [Google Scholar] [CrossRef] [PubMed]
- Lupini, A.; Sorgonà, A.; Princi, M.P.; Sunseri, F.; Abenavoli, M.R. Morphological and physiological effects of trans-cinnamic acid and its hydroxylated derivatives on maize root types. Plant Growth Regul. 2016, 78, 263–273. [Google Scholar] [CrossRef]
- Rubio, G.; Sorgonà, A.; Lynch, J.P. Spatial mapping of phosphorus influx in bean root systems using digital autoradiography. J. Exp. Bot. 2004, 55, 2269–2280. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [Green Version]
- York, L.M.; Nord, E.A.; Lynch, J.P. Integration of root phenes for soil resource acquisition. Front. Plant Sci. 2013, 4, 355. [Google Scholar] [CrossRef] [Green Version]
- Miguel, M.A.; Postma, J.A.; Lynch, J.P. Phene synergism between root hair length and basal root growth angle for phosphorus acquisition. Plant Physiol. 2015, 167, 1430–1439. [Google Scholar] [CrossRef]
- Rangarajan, H.; Postma, J.A.; Lynch, J.P. Co-optimization of axial root phenotypes for nitrogen and phosphorus acquisition in common bean. Ann. Bot. 2018, 122, 485–499. [Google Scholar] [CrossRef]
- Bodner, G.; Leitner, D.; Nakhforoosh, A.; Sobotik, M.; Moder, K.; Kaul, H.P. A statistical approach to root system classification. Front. Plant Sci. 2013, 4, 292. [Google Scholar] [CrossRef] [Green Version]
- Lê Cao, K.A.; Boitard, S.; Besse, P. Sparse PLS discriminant analysis: Biologically relevant feature selection and graphical displays for multiclass problems. BMC Bioinform. 2011, 12, 253. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.P.; Xu, W.W.; Velten, J.; Xin, Z.G.; Stout, J. Characterization of maize inbred lines for drought and heat tolerance. J. Soil Water Conserv. 2012, 67, 354–364. [Google Scholar] [CrossRef] [Green Version]
- Darling, E.S.; McClanahan, T.R.; Côté, I.M. Combined effects of two stressors on Kenyan coral reefs are additive or antagonistic, not synergistic. Conserv. Lett. 2010, 3, 122–130. [Google Scholar] [CrossRef]
- Jiang, M.; Wang, C.; Zhang, Y.; Feng, Y.; Wang, Y.; Zhu, Y. Sparse Partial-least-squares Discriminant Analysis for Different Geographical Origins of Salvia miltiorrhiza by 1H-NMR-based Metabolomics. Phytochem. Anal. 2014, 25, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Landi, P.; Albrecht, B.; Giuliani, M.; Sanguineti, M.C. Seedling characteristics in hydroponic culture and field performance of maize genotypes with different resistance to root lodging. Maydica 1998, 43, 111–116. [Google Scholar]
- Uga, Y.; Sugimoto, K.; Ogawa, S. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat. Genet. 2013, 45, 1097–1102. [Google Scholar] [CrossRef]
- Wasson, A.P.; Richards, R.A.; Chatrath, R.; Misra, S.C.; Prasad, S.V.; Rebetzke, G.J.; Kirkegaard, J.A.; Christopher, J.; Watt, M. Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J. Exp. Bot. 2012, 63, 3485–3498. [Google Scholar] [CrossRef] [Green Version]
- Ryser, P. Intra- and interspecific variation in root length, root turnover and the underlying parameters. In Inherent Variation in Plant Growth: Physiological Mechanisms and Ecological Consequences; Lambers, H., Poorter, H., Van Vuuren, M.M.I., Eds.; Backhuys Publishers: Leiden, The Netherlands, 1998; pp. 441–465. [Google Scholar]
- Ryser, P.; Lambers, H. Root and leaf attributes accounting for the performance of fast- and slow-growing grasses at different nutrient supply. Plant Soil 1995, 170, 251–265. [Google Scholar] [CrossRef]
- Ciamporova, M.; Dekankova, K.; Ovecka, M. Root morphology and anatomy of fast- and slow-growing grass species. In Variation in Plant Growth. Physiological Mechanisms and Ecological Consequences; Lambers, H., Poorter, H., Van Vuuren, M.M.I., Eds.; Backhuys Publishers: Leiden, The Netherlands, 1998; pp. 57–69. [Google Scholar]
- Wahl, S.; Ryser, P. Root tissue structure is linked to ecological strategies of grasses. New Phytol. 2000, 148, 459–471. [Google Scholar] [CrossRef]
- Hummel, I.; Vile, D.; Violle, C.; Devaux, J.; Ricci, B.; Blanchard, A.; Garnier, E.; Roumet, C. Relating root structure and anatomy to whole-plant functioning in 14 herbaceous Mediterranean species. New Phytol. 2007, 173, 313–321. [Google Scholar] [CrossRef]
- Chimungu, J.G.; Loades, K.W.; Lynch, J.P. Root anatomical phenes predict root penetration ability and biomechanical properties in maize (Zea Mays). J. Exp. Bot. 2015, 66, 3151–3162. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M.; Sharp, R.E. Complexity and coordination of root growth at low water potentials: Recent advances from transcriptomic and proteomic analyses. Plant Cell Environ. 2010, 33, 590–603. [Google Scholar] [CrossRef]
- Voothuluru, P.; Mäkelä, P.; Zhu, J.; Yamaguchi, M.; Cho, I.-J.; Oliver, M.J.; Simmonds, J.; Sharp, R.E. Apoplastic hydrogen peroxide in the growth zone of the maize primary root. Increased levels differentially modulate root elongation under well-watered and water-stressed conditions. Front. Plant Sci. 2020, 11, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, P.; Hochholdinger, F.; Li, C. Plasticity of Lateral Root Branching in Maize. Front. Plant Sci. 2019, 10, 363. [Google Scholar] [CrossRef] [PubMed]
- Prasad, P.V.V.; Pisipati, S.R.; Momcilovic, I.; Ristic, Z. Independent and combined effects of high temperature and drought stress during grain filling on plant yield and chloroplast EF-Tu expression in spring wheat. J. Agron. Crop. Sci. 2011, 197, 430–441. [Google Scholar] [CrossRef]
- Liu, H.; Able, A.J.; Able, J.A. Genotypic performance of Australian durum under single and combined water-deficit and heat stress during reproduction. Sci. Rep. 2019, 9, 14986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vile, D.; Pervent, M.; Belluau, M.; Vasseur, F.; Bresson, J.; Muller, B.; Granier, C.; Simonneau, T. Arabidopsis growth under prolonged high temperature and water deficit: Independent or interactive effects? Plant Cell Environ. 2012, 35, 702–718. [Google Scholar] [CrossRef]
- Orians, C.M.; Schweiger, R.; Dukes, J.S.; Scott, E.R.; Müller, C. Combined impacts of prolonged drought and warming on plant size and foliar chemistry. Ann. Bot. 2019, 124, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Tricker, P.J.; ElHabti, A.; Schmidt, J.; Fleury, D. The physiological and genetic basis of combined drought and heat tolerance in wheat. J. Exp. Bot. 2018, 69, 3195–3210. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Boon, N.J.; Webb, A.A.; Tanaka, R.J. Synergistic activation of RD29A via integration of salinity stress and Abscisic Acid in Arabidopsis thaliana. Plant Cell Physiol. 2016, 57, 2147–2160. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.; Rieu, I.; Mariani, C. How plants handle multiple stresses: Hormonal interactions underlying responses to abiotic stress and insect herbivory. Plant Mol. Biol. 2016, 91, 727–740. [Google Scholar] [CrossRef] [Green Version]
- Bengough, A.G.; McKenzie, B.M.; Hallett, P.D.; Valentine, T.A. Root elongation, water stress, and mechanical impedance: A review of limiting stresses and beneficial root tip traits. J. Exp. Bot. 2011, 62, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Nosalewicz, A.; Lipiec, J. The effect of compacted soil layers on vertical root distribution and water uptake by wheat. Plant Soil 2014, 375, 229–240. [Google Scholar] [CrossRef]
- Lynch, J.P. Rightsizing root phenotypes for drought resistance. J. Exp. Bot. 2018, 69, 3279–3292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tellah, S.; Badiani, M.; Trifilò, P.; Lo Gullo, M.A.; Ounane, G.; Ounane, S.M.; Sorgonà, A. Morpho-physiological traits contributing to water stress tolerance in a peanut (Arachis hypogaea L.) landraces collection from the Algerian Maghreb. Agrochimica 2014, 58, 126–147. [Google Scholar]
- Romano, A.; Sorgonà, A.; Lupini, A.; Araniti, F.; Stevanato, P.; Cacco, G.; Abenavoli, M.R. Morpho-physiological responses of sugar beet (Beta vulgaris L.) genotypes to drought stress. Acta Physiol. Plant. 2013, 35, 853–865. [Google Scholar] [CrossRef]
- Abenavoli, M.R.; Leone, M.; Sunseri, F.; Bacchi, M.; Sorgonà, A. Root phenotyping for drought tolerance in bean landraces from Calabria (Italy). J. Agron. Crop. Sci. 2016, 202, 1–12. [Google Scholar] [CrossRef]
- Zuffo, A.M.; Steiner, F.; Aguilera, J.G.; Teodoro, P.E.; Teodoro, L.P.R.; Busch, A. Multi-trait stability index: A tool for simultaneous selection of soya bean genotypes in drought and saline stress. J. Agron. Crop. Sci. 2020, 206, 815–822. [Google Scholar] [CrossRef]
- Chung, D.; Keles, S. Sparse Partial Least Squares Classification for High Dimensional Data. Stat. Appl. Genet. Mol. Boil. 2010, 9, 17. [Google Scholar] [CrossRef]
- Ryser, P.; Eek, L. Consequences of phenotypic plasticity vs. interspecific differences in leaf and root traits for acquisition of aboveground and belowground resources. Am. J. Bot. 2000, 87, 402–411. [Google Scholar] [CrossRef]
- Sorgonà, A.; Abenavoli, M.R.; Gringeri, P.G.; Cacco, G. Comparing morphological plasticity of root orders in slow- and fast-growing citrus rootstocks supplied with different nitrate levels. Ann. Bot. 2007, 100, 1287–1296. [Google Scholar] [CrossRef] [Green Version]
- Freschet, G.T.; Swart, E.M.; Cornelissen, J.H.C. Integrated plant phenotypic responses to contrasting above- and below-ground resources: Key roles of specific leaf area and root mass fraction. New Phytol. 2015, 206, 1247–1260. [Google Scholar] [CrossRef]
- Gelsomino, A.; Tortorella, D.; Cianci, V.; Petrovičová, B.; Sorgonà, A.; Piccolo, A.; Abenavoli, M.R. Effects of a biomimetic iron-porphyrin on soil respiration and maize root morphology as by a microcosm experiment. Z. Pflanz. Bodenkd. 2010, 173, 399–406. [Google Scholar] [CrossRef]
- Dubrovsky, J.G.; Forde, B.G. Quantitative Analysis of Lateral Root Development: Pitfalls and How to Avoid Them. Plant Cell 2012, 24, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bansal, S.; Hallsby, G.; Löfvenius, M.O.; Nilsson, M.C. Synergistic, additive and antagonistic impacts of drought and herbivory on Pinus sylvestris: Leaf, tissue and whole-plant responses and recovery. Tree Physiol. 2013, 33, 451–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.A. mixOmics: An R package for `omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Parameters | Treatments | |||
|---|---|---|---|---|---|
| Control | Drought | Heat | Combined | ||
| biometric | Fresh weight (g) | 0.128 (0.008) a | 0.131 (0.001) a | 0.147 (0.001) a | 0.162 (0.021) a |
| Dry weight (g) | 0.0111 (0.0009) b | 0.0116 (0.0006) b | 0.0131 (0.0003) b | 0.0168 (0.0017) a | |
| geometric | Length (cm) | 49 (1) a | 71 (6) a | 57 (4) a | 66 (12) a |
| Surface area (cm2) | 9.460 (0.009) a | 10.669 (0.896) a | 9.723 (0.587) a | 10.415 (1.766) a | |
| Length components | RLR (cm g−1) | 63 (4) b | 123 (12) a | 83 (2) b | 117 (15) a |
| RMR (g g−1) | 0.01381 (0.00016) c | 0.01840 (0.00002) b | 0.02004 (0.00051) b | 0.02702 (0.00205) a | |
| Fineness (cm cm−3) | 336 (11) c | 531 (11) a | 396 (8) b | 500 (6) a | |
| Tissue density (g cm−3) | 0.075 (0.007) c | 0.097 (0.006) b | 0.097 (0.003) b | 0.117 (0.007) a | |
| branching | Root zone formation (cm) | 9 (1) a,b | 3 (1) b | 9 (3) a | 7 (1) a,b |
| Branching zone formation (cm) | 42 (2) b | 65 (5) a | 48 (1) a,b | 64 (11) a | |
| Branching density (n cm−1) | 4.961 (0.101) a | 3.389 (0.051) c | 4.276 (0.027) b | 4.152 (0.284) b | |
| Category | Parameters | Treatments | |||
|---|---|---|---|---|---|
| Control | Drought | Heat | Combined | ||
| biometric | Fresh weight (g) | 0.285 (0.031) a | 0.172 (0.029) b | 0.205 (0.043) a,b | 0.176 (0.018) b |
| Dry weight (g) | 0.021 (0.004) a | 0.015 (0.003) a | 0.014 (0.004) a | 0.018 (0.002) a | |
| geometric | Length (cm) | 64 (7) a | 64 (7) a | 63 (8) a | 51 (2) a |
| Surface area(cm2) | 11 (1) a | 10 (1) a | 12 (2) a | 10 (1) a | |
| Length components | RLR(cm g−1) | 82 (13) a | 103 (14) a | 97 (14) a | 83 (1) a |
| RMR (g g−1) | 0.027 (0.002) a | 0.023 (0.002) a | 0.027 (0.007) a | 0.031 (0.003) a | |
| Fineness (cm cm−3) | 294 (47) b | 463 (21) a | 435 (55) a | 362 (24) a,b | |
| Tissue density (g cm−3) | 0.094 (0.007) a,b | 0.064 (0.023) b | 0.091 (0.003) a,b | 0.124 (0.001) a | |
| branching | Root zone formation (cm) | 7 (1) a,b | 8 (2) a,b | 12 (3) a | 6 (1) b |
| Branching zone formation (cm) | 54 (5) a | 55 (6) a | 57 (14) a | 44 (3) a | |
| Branching density (n cm−1) | 2.8 (0.1) b | 2.9 (0.1) b | 4.5 (0.4) a | 3.0 (0.2) b | |
| Category | Parameters | Treatments | |||
|---|---|---|---|---|---|
| Control | Drought | Heat | Combined | ||
| biometric | Fresh weight (g) | 0.212 (0.012) b | 0.280 (0.047) a,b | 0.315 (0.040) a | 0.225 (0.001) a,b |
| Dry weight (g) | 0.022 (0.004) a | 0.016 (0.001) a | 0.021 (0.003) a | 0.020 (0.001) a | |
| geometric | Length (cm) | 557 (8) c | 691 (35) b,c | 757(82) b | 1028 (33) a |
| Surface area (cm2) | 34.6 (0.7) b | 37.2 (2.7) b | 45.8 (4.8) a | 53.3 (0.1) a | |
| number (n) | 208 (16) a | 225 (14) a | 200 (2) a | 247 (27) a | |
| Average length (cm) | 2.860 (0.136) b | 3.076 (0.039) b | 3.640 (0.384) a,b | 4.486 (0.578) a | |
| Length components | RLR (cm g−1) | 736 (41) c | 1195 (63) b | 1125 (75) b | 1650 (90) a |
| RMR (g g−1) | 0.0324 (0.0079) a | 0.0220 (0.0003) a | 0.0308 (0.0031) a | 0.0330 (0.0016) a | |
| Fineness (cm cm−3) | 3257 (44) c | 4018 (56) b | 3512 (76) c | 4670 (288) a | |
| Tissue density (g cm−3) | 0.134 (0.027) a | 0.093 (0.005) a | 0.097 (0.005) a | 0.093 (0.006) a | |
| Category | Parameters | Treatments | |||
|---|---|---|---|---|---|
| Control | Drought | Heat | Combined | ||
| biometric | Fresh weight (g) | 0.0600 (0.0114) a,b | 0.0355 (0.0048) b | 0.1323 (0.0459) a | 0.0149 (0.0002) b |
| Dry weight (g) | 0.0051 (0.0010) a | 0.0035 (0.0006) a,b | 0.0025 (0.0008) b | 0.0013 (0.0002) b | |
| geometric | Length (cm) | 113 (7) a | 104 (1) a,b | 105 (13) a,b | 79 (14) b |
| Surface area (cm2) | 8.5 (1.3) a | 7.5 (0.1) a | 3.7 (0.6) b | 4.5 (0.9) b | |
| number (n) | 149 (21) a,b | 169 (18) a,b | 202 (23) a | 134 (5) b | |
| Average length (cm) | 0.872 (0.151) a | 0.707 (0.081) a | 0.563 (0.136) a | 0.530 (0.094) a | |
| Length components | RLR (cm g−1) | 147 (1) a,b | 178 (2) a | 151 (12) a,b | 128 (19) b |
| RMR (g g−1) | 0.0066 (0.0010) a | 0.0032 (0.0001) b | 0.0041 (0.0015) a,b | 0.0021 (0.0003) b | |
| Fineness (cm cm−3) | 2728 (48) b | 2416 (0) b | 3630 (376) b | 6283 (1176) a | |
| Tissue density (g cm−3) | 0.095 (0.004) a | 0.085 (0.015) a | 0.101 (0.036) a | 0.102 (0.022) a | |
| Factor | df | R2 | F | p | ||
|---|---|---|---|---|---|---|
| Stress treatment | 3 | 0.6693 | 8.0955 | <0.001 | ||
| Residual | 12 | 0.3307 | ||||
| Total | 15 | 1.0000 | ||||
| Pairwise contrasts | ||||||
| F model | R2 | p | ||||
| C vs. D | 9.262202 | 0.6068719 | 0.031 | |||
| C vs. H | 3.877963 | 0.3925873 | 0.053 | |||
| C vs. HD | 15.925573 | 0.7263469 | 0.027 | |||
| D vs. H | 7.056074 | 0.5404438 | 0.034 | |||
| D vs. HD | 13.677155 | 0.6950779 | 0.023 | |||
| H vs. HD | 7.751958 | 0.5636985 | 0.054 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vescio, R.; Abenavoli, M.R.; Sorgonà, A. Single and Combined Abiotic Stress in Maize Root Morphology. Plants 2021, 10, 5. https://doi.org/10.3390/plants10010005
Vescio R, Abenavoli MR, Sorgonà A. Single and Combined Abiotic Stress in Maize Root Morphology. Plants. 2021; 10(1):5. https://doi.org/10.3390/plants10010005
Chicago/Turabian StyleVescio, Rosa, Maria Rosa Abenavoli, and Agostino Sorgonà. 2021. "Single and Combined Abiotic Stress in Maize Root Morphology" Plants 10, no. 1: 5. https://doi.org/10.3390/plants10010005
APA StyleVescio, R., Abenavoli, M. R., & Sorgonà, A. (2021). Single and Combined Abiotic Stress in Maize Root Morphology. Plants, 10(1), 5. https://doi.org/10.3390/plants10010005