The Impact of Human Pressure and Climate Change on the Habitat Availability and Protection of Cypripedium (Orchidaceae) in Northeast China

 , and
, and
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Species
2.2. Study Area
2.3. Environmental Niche Modeling
2.3.1. Data Preparation and Selection
- Kira’s warmth index (WI) [62],
- Holdridge’s annual biotemperature (ABT) [63],
- Kira’s coldness index (CI) [62],
- Seasonality of precipitation (PSD),
- |WI-CI|,|WI-CI| = |warmth index − coldness index|.
2.3.2. Model Evaluation
2.3.3. Threshold Selection
3. Results
3.1. Model Performance and Key Environmental Variables
3.2. Suitable Habitats of Cypripedium in Northeast China
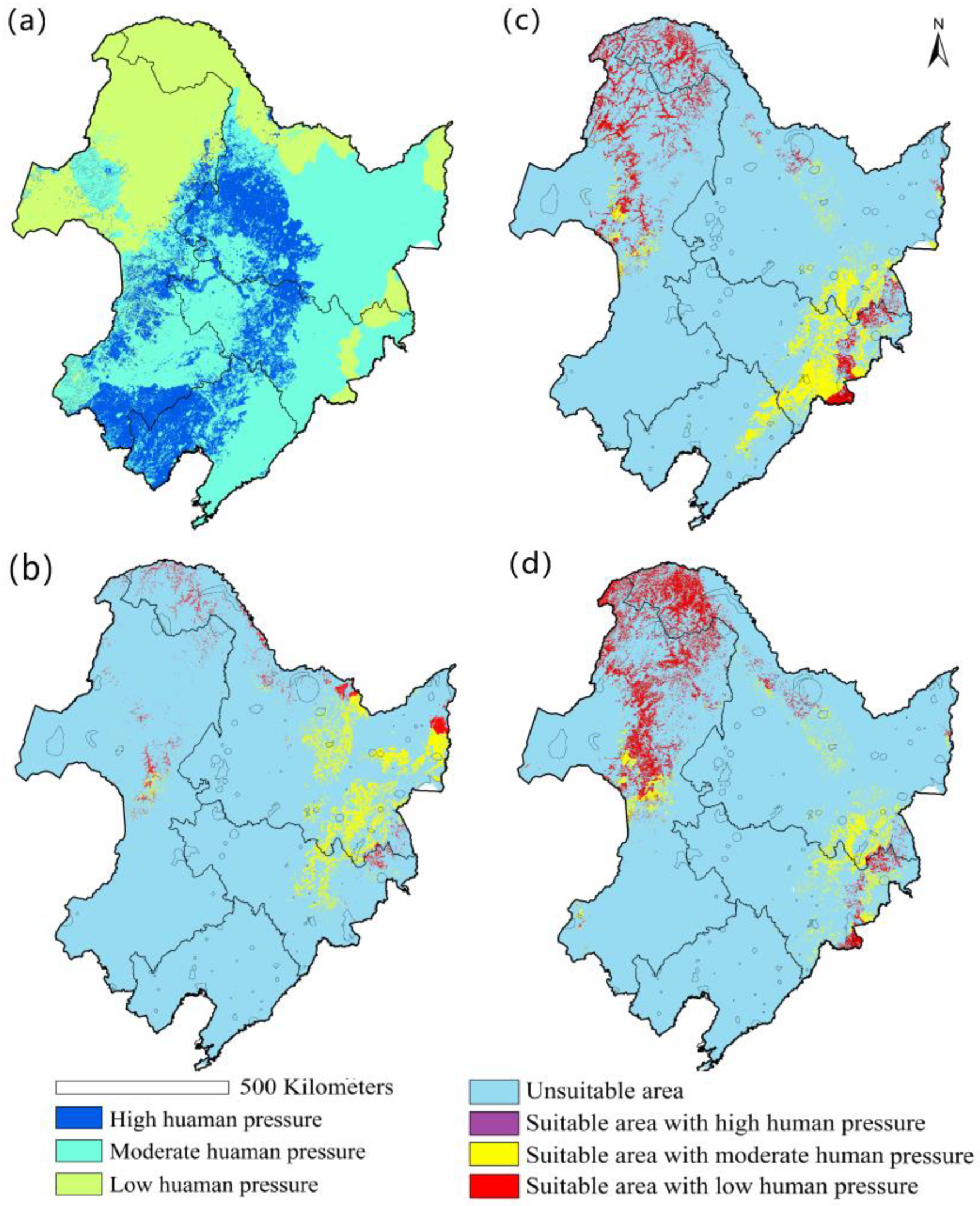
3.3. The Impact of Human Pressure on the Distribution of Cypripedium
3.4. The Suitable Area of Cypripedium within Nature Reserve Area
3.5. Future Suitable Habitats Prediction of Cypripedium
4. Discussion
4.1. Ecological Niche of Cypripedium
4.2. Future Protection under Climate Change and Human Pressure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adhikari, D.; Barik, S.K.; Upadhaya, K. Habitat distribution modelling for reintroduction of Ilex khasiana Purk, a critically endangered tree species of northeastern India. Ecol. Eng. 2012, 40, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time Annual Review of Ecology Evolution and Systematics. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Fei, S.L.; Liang, L.; Paillet, F.L.; Steiner, K.C.; Fang, J.Y.; Shen, Z.H.; Wang, Z.H.; Hebard, V.F. Modelling chestnut biogeography for American chestnut restoration. Divers. Distrib. 2012, 18, 754–768. [Google Scholar] [CrossRef] [Green Version]
- Garcia, K.; Lasco, R.; Ines, A.; Lyon, B.; Pulhin, F. Predicting geographic distribution and habitat suitability due to climate change of selected threatened forest tree species in the Philippines. Appl. Geogr. 2013, 44, 12–22. [Google Scholar] [CrossRef]
- Yi, Y.J.; Cheng, X.; Yang, Z.F.; Zhang, S.H. Maxent modeling for predicting the potential distribution of endangered medicinal plant (H, riparia Lour) in Yunnan China. Ecol. Eng. 2016, 92, 260–269. [Google Scholar] [CrossRef]
- Stipkova, Z.; Romportl, D.; Cernocka, V.; Kindlmann, P. Factors associated with the distributions of orchids in the Jeseníky mountains, Czech Republic. Eur. J. Environ. Sci. 2017, 7, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Li, X.; Zhao, Z.; Nawaz, Z. Predicting the impacts of climate change soils and vegetation types on the geographic distribution of Polyporus umbellatus in China. Sci. Total. Environ. 2019, 648, 1–11. [Google Scholar] [CrossRef]
- Rolland, V.; Weimerskirch, H.; Barbraud, C. Relative influence of fisheries and climate on the demography of four albatross species. Glob. Chang. Biol. 2010, 16, 1910–1922. [Google Scholar] [CrossRef]
- Lorenzen, E.D.; Nogues-Bravo, D.; Orlando, L.; Weinstock, J.; Binladen, J.; Marske, K.A.; Ugan, A.; Borregaard, M.K.; Gilbert, M.T.P.; Nielsen, R.; et al. Species-specific responses of Late Quaternary megafauna to climate and humans. Nature 2011, 479, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Edwards, H.H. Potential impacts of climate change on warmwater megafauna: The Florida manatee example (Trichechus manatus latirostris). Clim. Chang. 2013, 121, 727–738. [Google Scholar] [CrossRef]
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Borger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Wang, W.H.; Qu, Y.K.; Fu, J.; Jiang, S.Q.; Wang, D.L.; Kaplan, I.; Ren, B.Z. Resource-mediated effects of grazing and irrigation on insect diversity in a meadow steppe. Insect. Conserv. Divers. 2019, 12, 29–38. [Google Scholar] [CrossRef] [Green Version]
- McKinney, M.L.; Lockwood, J.L. Biotic homogenization: A few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 1999, 14, 450–453. [Google Scholar] [CrossRef]
- Zhang, W.G.; Hu, Y.M.; Hu, J.C.; Yu, C.; Jing, Z.; Miao, L. Impacts of land-use change on mammal diversity in the upper reaches of Minjiang River China: Implications for biodiversity conservation planning. Landsc. Urban Plan. 2008, 85, 195–204. [Google Scholar] [CrossRef]
- Kubiszewski, I.; Costanza, R.; Dorji, L.; Thoennes, P.; Tshering, K. An initial estimate of the value of ecosystem services in Bhutan. Ecosyst. Serv. 2013, 3, E11–E21. [Google Scholar] [CrossRef]
- Malavasi, M.; Santoro, R.; Cutini, M.; Acosta, A.T.R.; Carranza, M.L. The impact of human pressure on landscape patterns and plant species richness in Mediterranean coastal dunes. Plant Biosyst. 2016, 150, 73–82. [Google Scholar] [CrossRef]
- Hoffmann, M.; Hilton-Taylor, C.; Angulo, A.; Bohm, M.; Brooks, T.M.; Butchart, S.H.M.; Carpenter, K.E.; Chanson, J.; Collen, B.; Cox, N.A.; et al. The Impact of Conservation on the Status of the World’s Vertebrates. Science 2010, 330, 1503–1509. [Google Scholar] [CrossRef] [Green Version]
- Di Marco, M.; Boitani, L.; Mallon, D.; Hoffmann, M.; Iacucci, A.; Meijaard, E.; Visconti, P.; Schipper, J.; Rondinini, C. A Retrospective Evaluation of the Global Decline of Carnivores and Ungulates. Conserv. Biol. 2014, 28, 1109–1118. [Google Scholar] [CrossRef]
- Allan, J.R.; Watson, J.E.M.; Di Marco, M.; O’Bryan, C.J.; Possingham, H.P.; Atkinson, S.C.; Venter, O. Hotspots of human impact on threatened terrestrial vertebrates. PLoS Biol. 2019, 17, e3000158. [Google Scholar] [CrossRef] [Green Version]
- Venter, O.; Sanderson, E.W.; Magrach, A.; Allan, J.R.; Beher, J.; Jones, K.R.; Possingham, H.P.; Laurance, W.F.; Wood, P.; Fekete, B.M.; et al. Sixteen years of change in the global terrestrial human footprint and implications for biodiversity conservation. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, J.E.M.; Evans, T.; Venter, O.; Williams, B.; Tulloch, A.; Stewart, C.; Thompson, I.; Ray, J.C.; Murray, K.; Salazar, A.; et al. The exceptional value of intact forest ecosystems. Nat. Ecol. Evol. 2018, 5, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.R.; Venter, O.; Fuller, R.A.; Allan, J.R.; Maxwell, S.L.; Negret, P.J.; Watson, J.E.M. Protected land: Threat of invasive species Response. Science 2018, 361, 562–563. [Google Scholar] [CrossRef] [PubMed]
- Di Marco, M.; Venter, O.; Possingham, H.P.; Watson, J.E.M. Changes in human footprint drive changes in species extinction risk. Nat. Commu. 2018, 9. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Harley, C.D.G. Climate Change Keystone Predation and Biodiversity Loss. Science 2011, 334, 1124–1127. [Google Scholar] [CrossRef]
- Feeley, K.J.; Malhi, Y.; Zelazowski, P.; Silman, M.R. The relative importance of deforestation precipitation change and temperature sensitivity in determining the future distributions and diversity of Amazonian plant species. Glob. Chang. Biol. 2012, 18, 2636–2647. [Google Scholar] [CrossRef]
- Wang, W.J.; Thompson, F.R.; He, H.S.; Fraser, J.S.; Dijak, W.D.; Spetich, M.A. Population dynamics has greater effects than climate change on tree species distribution in a temperate forest region. J. Biogeogr. 2018, 45, 2766–2778. [Google Scholar] [CrossRef]
- Duffy, K.J.; Jacquemyn, H. Climate change increases ecogeographical isolation between closely related plants. J. Ecol. 2019, 107, 167–177. [Google Scholar] [CrossRef]
- Mantyka-Pringle, C.S.; Martin, T.G.; Rhodes, J.R. Interactions between climate and habitat loss effects on biodiversity: A systematic review and meta-analysis. Glob. Chang. Biol. 2012, 18, 1239–1252. [Google Scholar] [CrossRef] [Green Version]
- Gong, M.; Guan, T.; Hou, M.; Liu, G.; Zhou, T. Hopes and challenges for giant panda conservation under climate change in the Qinling Mountains of China. Ecol. Evol. 2017, 7, 596–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Summers, D.M.; Bryan, B.A.; Crossman, N.D.; Meyer, A.S. Species vulnerability to climate change: Impacts on spatial conservation priorities and species representation. Glob. Change Biol. 2012, 18, 3268. [Google Scholar] [CrossRef]
- Wang, C.J.; Liu, C.Z.; Wan, J.Z.; Zhang, Z.X. Climate change may threaten habitat suitability of threatened plant species within Chinese nature reserves. PeerJ 2016, 4, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, E.S.; Lischke, H.; Schmatz, D.R.; Zimmermann, N.E. Climate competition and connectivity affect future migration and ranges of European trees. Glob. Ecol. Biogeogr. 2012, 21, 164–178. [Google Scholar] [CrossRef]
- Barber, Q.E.; Nielsen, S.E.; Hamann, A. Assessing the vulnerability of rare plants using climate change velocity habitat connectivity and dispersal ability: A case study in Alberta Canada. Reg. Environ. Chang. 2016, 16, 1433–1441. [Google Scholar] [CrossRef]
- Moran, E.V.; Hartig, F.; Bell, D.M. Intraspecific trait variation across scales: Implications for understanding global change responses. Glob. Chang. Biol. 2016, 22, 137–150. [Google Scholar] [CrossRef] [Green Version]
- Llewelyn, J.; Macdonald, S.L.; Hatcher, A.; Moritz, C.; Phillips, B.L. Intraspecific variation in climate-relevant traits in a tropical rainforest lizard. Divers. Distrib. 2016, 22, 1000–1012. [Google Scholar] [CrossRef]
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Tsiftsis, S.; Stipkova, Z.; Kindlmann, P. Role of way of life latitude elevation and climate on the richness and distribution of orchid species. Biodivers. Conserv. 2019, 28, 75–96. [Google Scholar] [CrossRef]
- Diez, J.M.; Pulliam, H.R. Hierarchical analysis of species distributions and abundance across environmental gradients. Ecology 2007, 88, 3144–3152. [Google Scholar] [CrossRef]
- McCormick, M.K.; Jacquemyn, H. What constrains the distribution of orchid populations? New Phytol. 2014, 202, 392–400. [Google Scholar] [CrossRef]
- Duffy, K.J.; Johnson, S.D. Effects of distance from models on the fitness of floral mimics. Plant Biol. 2017, 19, 438–443. [Google Scholar] [CrossRef]
- Joffard, N.; Massol, F.; Grenie, M.; Montgelard, C.; Schatz, B. Effect of pollination strategy phylogeny and distribution on pollination niches of Euro-Mediterranean orchids. J. Ecol. 2019, 107, 478–490. [Google Scholar] [CrossRef] [Green Version]
- Stipkova, Z.; Tsiftsis, S.; Kindlmann, P. Pollination Mechanisms are Driving Orchid Distribution in Space. Sci. Rep. 2020, 10, 850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquemyn, H.; Brys, R.; Hermy, M.; Willems, J.H. Does nectar reward affect rarity and extinction probabilities of orchid species? An assessment using historical records from Belgium and the Netherlands. Biol. Conserv. 2005, 121, 257–263. [Google Scholar] [CrossRef]
- Swarts, N.D.; Dixon, K.W. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izawa, F.; Brzosko, E.; Till-Bottraud, I. Population viability analysis of Cypripedium calceolus in a protected area: Longevity stability and persistence. J. Ecol. 2005, 93, 716–726. [Google Scholar] [CrossRef]
- Izawa, T.; Kawahara, T.; Takahashi, H. Genetic diversity of an endangered plant Cypripedium macranthos var, rebunense (Orchidaceae): Background genetic research for future conservation. Conserv. Genet. 2007, 8, 1369–1376. [Google Scholar] [CrossRef]
- Minasiewicz, J.; Znaniecka, J.M.; Gorniak, M.; Kawinski, A. Spatial genetic structure of an endangered orchid Cypripedium calceolus (Orchidaceae) at a regional scale: Limited gene flow in a fragmented landscape. Conserv. Genet. 2018, 19, 1449–1460. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.Y.; Peter, H.R.; Hong, D. Folra of China; Science Press: Beijing, China, 2004. [Google Scholar]
- Guo, J.L.; Cao, W.J.; Li, Z.M.; Zhang, Y.H.; Volis, S. Conservation implications of population genetic structure in a threatened orchid Cypripedium tibeticum. Plant Divers. 2019, 41, 13–18. [Google Scholar] [CrossRef]
- Kull, T. Fruit-set and recruitment in populations of Cypripedium calceolus L in Estonia. Bot. J. Linn. Soc. 1998, 126, 27–38. [Google Scholar] [CrossRef]
- Weng, E.S.; Hu, H.; Li, S.Y.; Huang, J.L. Differentiation of Flower Bud of Cypripedium flavum. Acta Bot. Yunnanica 2002, 24, 222–228. [Google Scholar]
- Zhang, Y.; Wang, M.M.; Zhang, X.; Zhao, S.W. The Study on Numerical Taxonomy of Cypripedium in North and Northeast China. Acta Hortic. Sin. 2013, 40, 933–942. [Google Scholar] [CrossRef]
- Zhang, G.Y. Studies on Investigation and Genetic Diversity of Cypriprdium in Changbai Mountains. Master’s Thesis, Jilin Agricultural University, Changchun, China, 2013. (In Chinese). [Google Scholar]
- Yin, W.P.; Li, G.F.; Zhao, Z.Z.; Chen, F.Y.; Qiu, G.L.; Liu, W.S.; Mu, L.Q. Species diversity and distribution of orchids in Heilongjiang Zhongyangzhan Black-billed Capercaillie National Nature Reserve. J. Qiqihar. Univ. (Nat. Sci. Ed.) 2019, 35, 52–56. [Google Scholar]
- Gong, H.L.; Meng, D.; Li, X.; Zhu, F. Soil degradation and food security coupled with global climate change in northeastern China. Chin. Geogr. Sci. 2013, 23, 562–573. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.W.; Zhang, Z.X.; Jin, Y.H.; Wang, P.; Wang, X.Z. Regionalization and distribution pattern of vegetation of Northeast China. Chinese J. Plant Ecol. 2010, 34, 1359–1368. [Google Scholar] [CrossRef]
- Venter, O.; Sanderson, E.W.; Magrach, A.; Allan, J.R.; Beher, J.; Jones, K.R.; Possingham, H.P.; Laurance, W.F.; Wood, P.; Fekete, B.M.; et al. Data from: Global terrestrial Human Footprint maps for 1993 and 2009. Dryad. Digit. Repos. 2016. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2013: The Physical Science Basis, Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: New York, NY, USA, 2013. [Google Scholar]
- Science On a Sphere. Available online: https://sos.noaa.gov/datasets/climate-model-temperature-change-rcp-45-2006-2100/ (accessed on 7 May 2020).
- Kira’s warmth, T. Forest ecosystems of east and southeaat-asia in a global perspective. Ecol. Res. 1991, 6, 185–200. [Google Scholar] [CrossRef]
- Holdridge, L.R. Determination of world plant formations from simple cliamte data. Science 1947, 105, 367–3682. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudik, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of Maxent for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Cribb, P.; Sandison, M.S. A preliminary assessment of the conservation status of Cypripedium species in the wild. Bot. J. Linn. Soc. 1998, 126, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.R.; Zhang, S.B.; Hu, H.; Li, D.Z. Photosynthetic performance along a light gradient as related to leaf characteristics of a naturally occurring Cypripedium flavum. J. Plant Res. 2008, 121, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liu, Z.; Chen, L.; Li, Q. The Genus Cypripedium in China; Science Press: Beijing, China, 2012. [Google Scholar]
- Zweig, M.H.; Campbell, G. Receiver-opreating characteristic (ROC) plots—A fundamental evaluation tool in clinical medcine. Clin. Chem. 1993, 39, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Ramirez-Barahona, S.; Torres-Miranda, A.; Palacios-Rios, M.; Luna-Vega, I. Historical biogeography of the Yucatan Peninsula Mexico: A perspective from ferns (Monilophyta) and lycopods (Lycophyta). Bot. J. Linn. Soc. 2009, 98, 775–786. [Google Scholar] [CrossRef] [Green Version]
- Song, C.Y.; Liu, H.M. Habitat differentiation and conservation gap of Magnolia biondii M-denudata and M-sprengeri in China. PeerJ 2019, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Brito, J.C.; Acosta, A.L.; Alvares, F.; Cuzin, F. Biogeography and conservation of taxa from remote regions: An application of ecological-niche based models and GIS to North-African Canids. Biol. Conserv. 2009, 142, 3020–3029. [Google Scholar] [CrossRef]
- Liu, C.R.; White, M.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Wan, J.; Wang, C.; Han, S.; Yu, J. Planning the priority protected areas of endangered orchid species in northeastern China. Biodivers. Conserv. 2014, 23, 1395–1409. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudik, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurskainen, S.; Jakalaniemi, A.; Kaitala, V.; Kull, T.; Motlep, M.; Ramula, S.; Tuomi, J. Temporal cycles and spatial asynchrony in the reproduction and growth of a rare nectarless orchid Cypripedium calceolus. Bot. J. Linn. Soc. 2017, 183, 316–326. [Google Scholar] [CrossRef] [Green Version]
- Bunch, W.D.; Cowden, C.C.; Wurzburger, N.; Shefferson, R.P. Geography and soil chemistry drive the distribution of fungal associations in lady’s slipper orchid Cypripedium acaule. Botany 2013, 91, 850–856. [Google Scholar] [CrossRef]
- Suggitt, A.J.; Wilson, R.J.; Isaac, N.J.B.; Beale, C.M.; Auffret, A.G.; August, T.; Bennie, J.J.; Crick, H.Q.P.; Duffield, S.; Fox, R.; et al. Extinction risk from climate change is reduced by microclimatic buffering. Nat. Clim. Chang. 2018, 8, 713. [Google Scholar] [CrossRef]
- Opedal, O.H.; Armbruster, W.S.; Graae, B.J. Linking small-scale topography with microclimate plant species diversity and intra-specific trait variation in an alpine landscape. Plant. Ecol. Divers. 2015, 8, 305–315. [Google Scholar] [CrossRef] [Green Version]
- McCormick, M.K.; Whigham, D.F.; Canchani-Viruet, A. Mycorrhizal fungi affect orchid distribution and population dynamics. New Phytol. 2018, 219, 1207–1215. [Google Scholar] [CrossRef] [Green Version]
- Jacquemyn, H.; Waud, M.; Brys, R.; Lallemand, F.; Courty, P.E.; Robionek, A.; Selosse, M.A. Mycorrhizal Associations and Trophic Modes in Coexisting Orchids: An Ecological Continuum between Auto- and Mixotrophy. Front. Plant Sci. 2017, 8, 1497. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Classification | Description | Data Source | Details |
|---|---|---|---|---|
| Bio1 | Climate | Annual Mean Temperature | Worldclim database | |
| Bio2 | Climate | Mean Diurnal Range | Worldclim database | |
| Bio3 | Climate | Isothermality | Worldclim database | |
| Bio4 | Climate | Temperature Seasonality | Worldclim database | |
| Bio5 | Climate | Maximum Temperature of Warmest Month | Worldclim database | |
| Bio6 | Climate | Minimum Temperature of Coldest Month | Worldclim database | |
| Bio7 | Climate | Temperature Annual Range | Worldclim database | |
| Bio8 | Climate | Mean Temperature of Wettest Quarter | Worldclim database | |
| Bio9 | Climate | Mean Temperature of Driest Quarter | Worldclim database | |
| Bio10 | Climate | Mean Temperature of Warmest Quarter | Worldclim database | |
| Bio11 | Climate | Mean Temperature of Coldest Quarter | Worldclim database | |
| Bio12 | Climate | Annual Precipitation | Worldclim database | |
| Bio13 | Climate | Precipitation of Wettest Month | Worldclim database | |
| Bio14 | Climate | Precipitation of Driest Month | Worldclim database | |
| Bio15 | Climate | Precipitation Seasonality | Worldclim database | |
| Bio16 | Climate | Precipitation of Wettest Quarter | Worldclim database | |
| Bio17 | Climate | Precipitation of Driest Quarter | Worldclim database | |
| Bio18 | Climate | Precipitation of Warmest Quarter | Worldclim database | |
| Bio19 | Climate | Precipitation of Coldest Quarter | Worldclim database | |
| ABT | Climate | Annual Biotemperature | Worldclim database | |
| CI | Climate | Coldness Index | Worldclim database | |
| WI | Climate | Warmth Index | Worldclim database | |
| |WI-CI| | Climate | Absolute Value of Warmth Index - Coldness Index | Worldclim database | |
| Ele | Topography | Elevation | Geospatial Data Cloud | |
| Slo | Topography | Slope | Geospatial Data Cloud | |
| Asp | Topography | Aspect | Geospatial Data Cloud | |
| OC | Soil | Topsoil Organic Carbon Content | Harmonized World Soil Database | The percentage of organic carbon in topsoil |
| pH | Soil | Topsoil pH | Harmonized World Soil Database | Measured in a soil–water solution, it is a measure for the acidity and alkalinity of the soil |
| Veg | Vegetation | Vegetation Type | Data Center for Resources and Environmental Sciences | Distribution of 11 vegetation types in China |
| Built | Human activity | Built Environments | Human Footprint maps | All areas mapped as built given a score of 10 |
| Pop | Human activity | Population Density | Human Footprint maps | Pressure score = 3.333 × log (population density + 1) |
| Nig | Human activity | Night-time Density | Human Footprint maps | Equal quintile bins |
| Crop | Human activity | Croplands | Human Footprint maps | All areas mapped as crops given a score of 7 |
| Pas | Human activity | Pasture | Human Footprint maps | All areas mapped as pasture given a score of 4 |
| Roa | Human activity | Roads | Human Footprint maps | 500 m either side of roads given a direct pressure score of 8 Starting 500 m out from a road, a pressure score of 4 exponentially decaying out to 15 km |
| Rail | Human activity | Railways | Human Footprint maps | 500 m either side of railways given a direct pressure score of 8 Starting 500 m out from a road, a |
| Nav | Human activity | Navigable Waterways | Human Footprint maps | pressure score of 4 exponentially decaying out to 15 km |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Jacquemyn, H.; He, X.; Chen, W.; Huang, Y.; Yu, S.; Lu, Y.; Zhang, Y. The Impact of Human Pressure and Climate Change on the Habitat Availability and Protection of Cypripedium (Orchidaceae) in Northeast China. Plants 2021, 10, 84. https://doi.org/10.3390/plants10010084
Liu H, Jacquemyn H, He X, Chen W, Huang Y, Yu S, Lu Y, Zhang Y. The Impact of Human Pressure and Climate Change on the Habitat Availability and Protection of Cypripedium (Orchidaceae) in Northeast China. Plants. 2021; 10(1):84. https://doi.org/10.3390/plants10010084
Chicago/Turabian StyleLiu, Huanchu, Hans Jacquemyn, Xingyuan He, Wei Chen, Yanqing Huang, Shuai Yu, Yupeng Lu, and Yue Zhang. 2021. "The Impact of Human Pressure and Climate Change on the Habitat Availability and Protection of Cypripedium (Orchidaceae) in Northeast China" Plants 10, no. 1: 84. https://doi.org/10.3390/plants10010084
APA StyleLiu, H., Jacquemyn, H., He, X., Chen, W., Huang, Y., Yu, S., Lu, Y., & Zhang, Y. (2021). The Impact of Human Pressure and Climate Change on the Habitat Availability and Protection of Cypripedium (Orchidaceae) in Northeast China. Plants, 10(1), 84. https://doi.org/10.3390/plants10010084