The Habitat of the Neglected Independent Protonemal Stage of Buxbaumia viridis

Abstract
:1. Introduction
2. Results
2.1. Distribution of Buxbaumia viridis
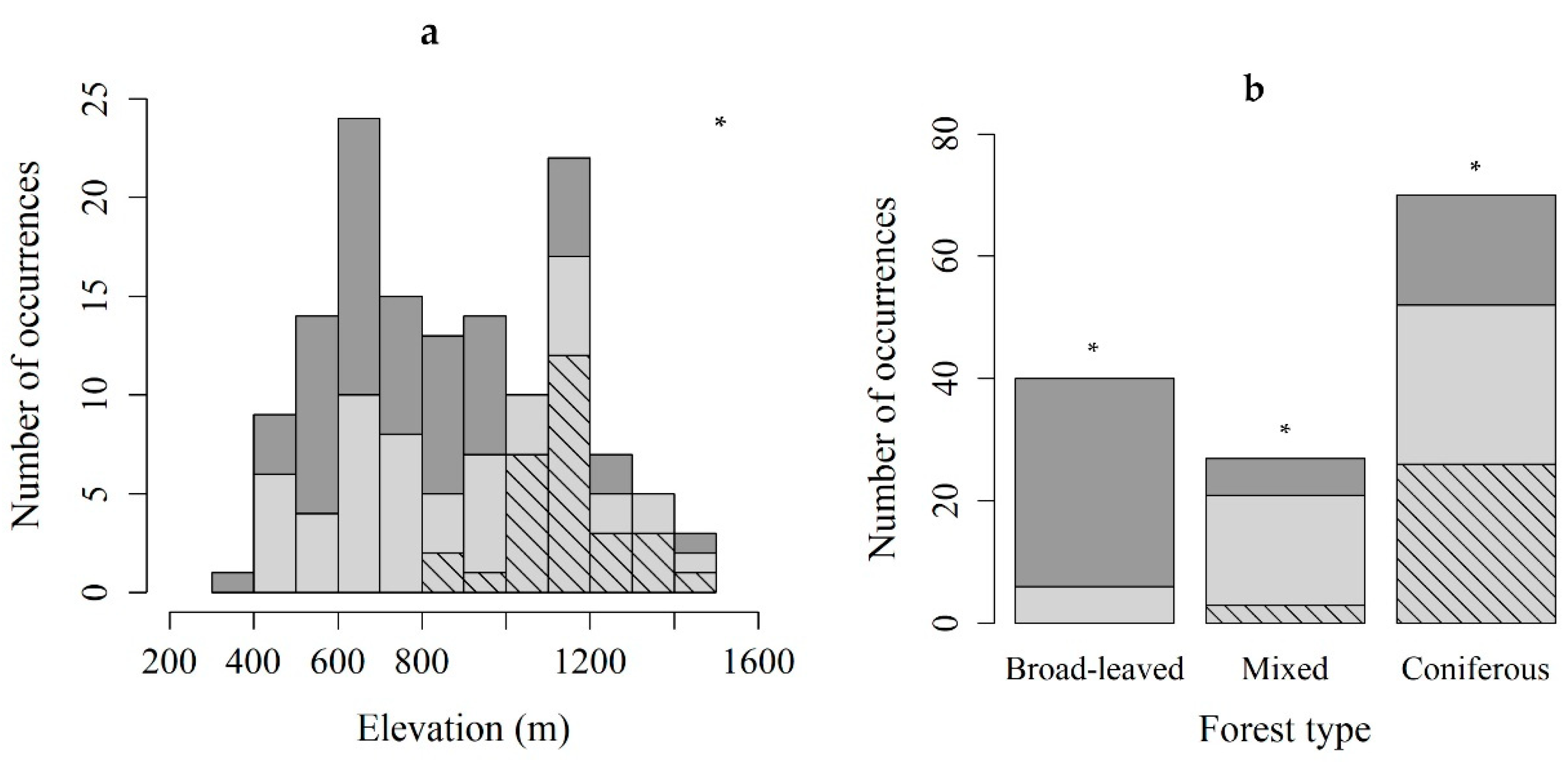
2.2. Habitat

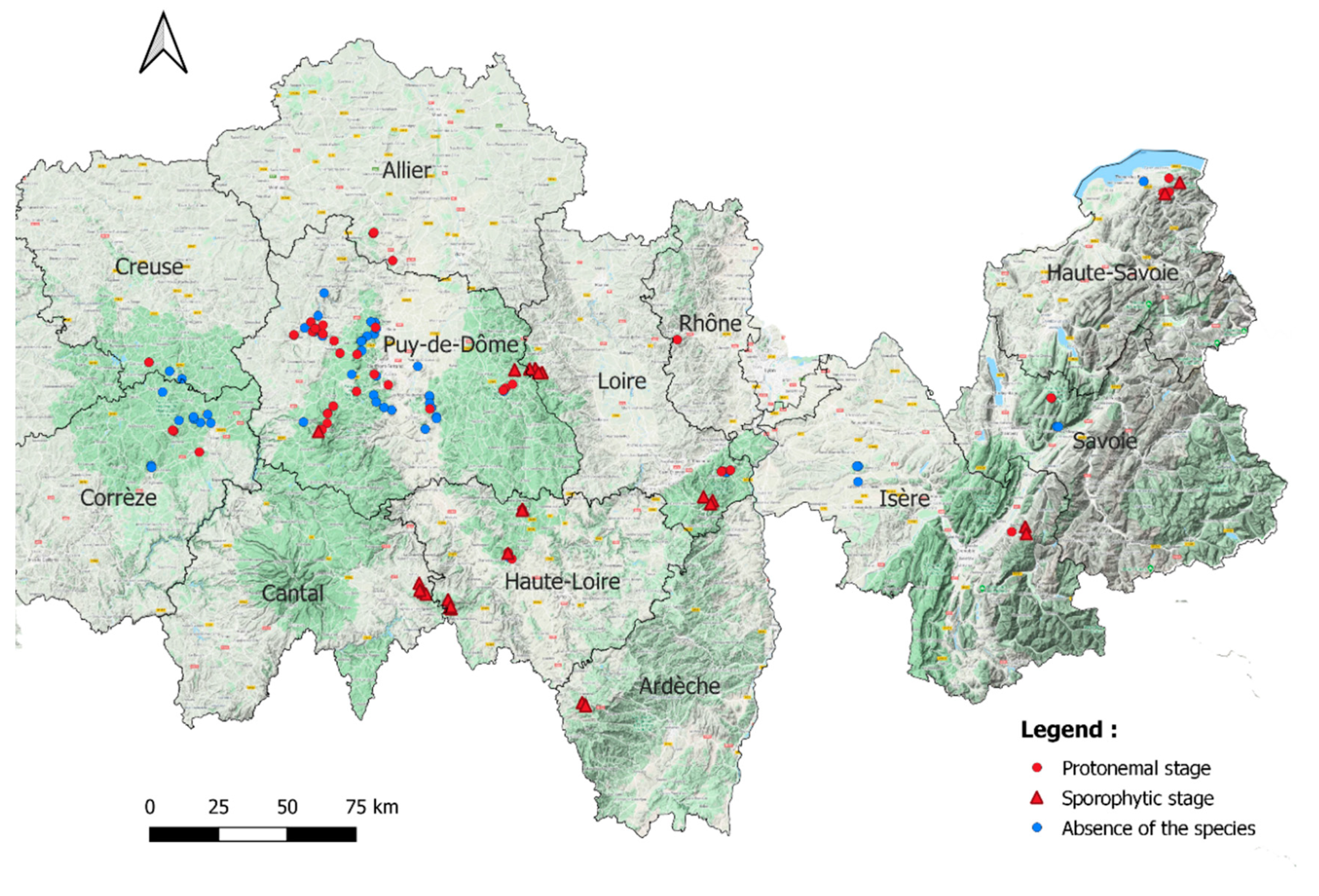{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | ||||
|---|---|---|---|---|
| Variable | Mean ± SD | Min | Max | |
| Elevation (m a.s.l) | All stands | 869 ± 273 | 313 | 1452 |
| with B. viridis | 946 ± 276 | 421 | 1452 | |
| with sporophytes | 1142 ± 135 | 840 | 1452 | |
| Slope (degrees) | All stands | 17 ± 13 | 0 | 50 |
| With B. viridis | 18 ± 14 | 0 | 50 | |
| With sporophytes | 17 ± 14 | 2 | 45 | |
| Distance to the nearest watercourse (m) | All stands | 366 ± 593 | 8 | 3330 |
| With B. viridis | 279 ± 418 | 8 | 3190 | |
| With sporophytes | 224 ± 222 | 10 | 1156 | |
| Deadwood surface (%) | All stands | 14 ± 7 | 3 | 33 |
| With B. viridis | 18 ± 6 | 3 | 33 | |
| With sporophytes | 19 ± 6 | 5 | 33 | |
| Northness | All stands | 0.29 ± 0.67 | −1 | 1 |
| With B. viridis | 0.40 ± 0.61 | −1 | 1 | |
| With sporophytes | 0.55 ± 0.48 | −0.71 | 1 | |
| (b) | ||||
| Forest Type | No All stands | No with B. viridis | No with sporophytes | |
| Broad-leaved | 40 | 6 | 0 | |
| Mixed | 27 | 21 | 3 | |
| Coniferous | 70 | 52 | 26 | |
| Total | 137 | 79 | 29 | |
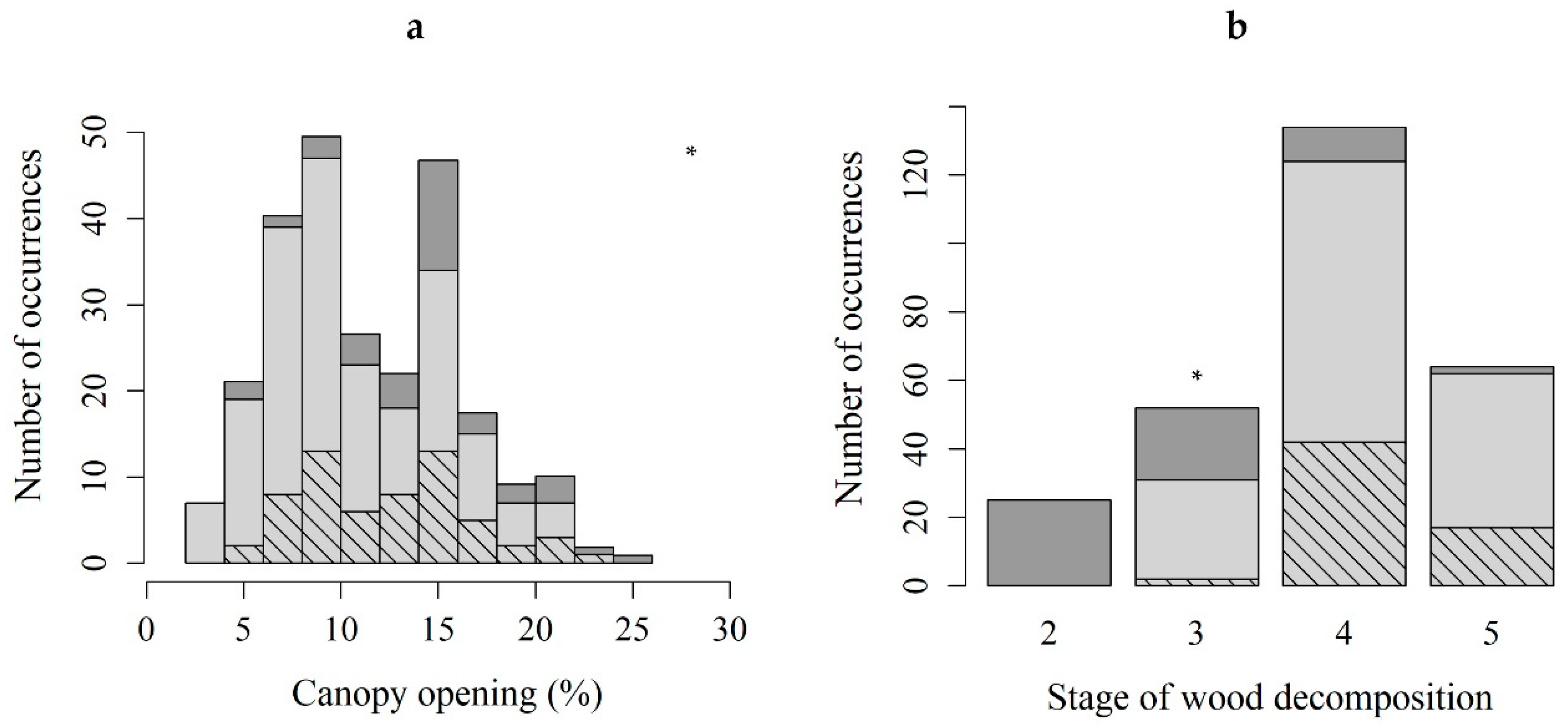
2.3. Microhabitat
3. Discussion
3.1. The Independent Protonemal Populations
3.2. Habitat and Microhabitat Comparison between Protonemal and Sporophyte Sites
3.3. Conservation Implications
4. Materials and Methods
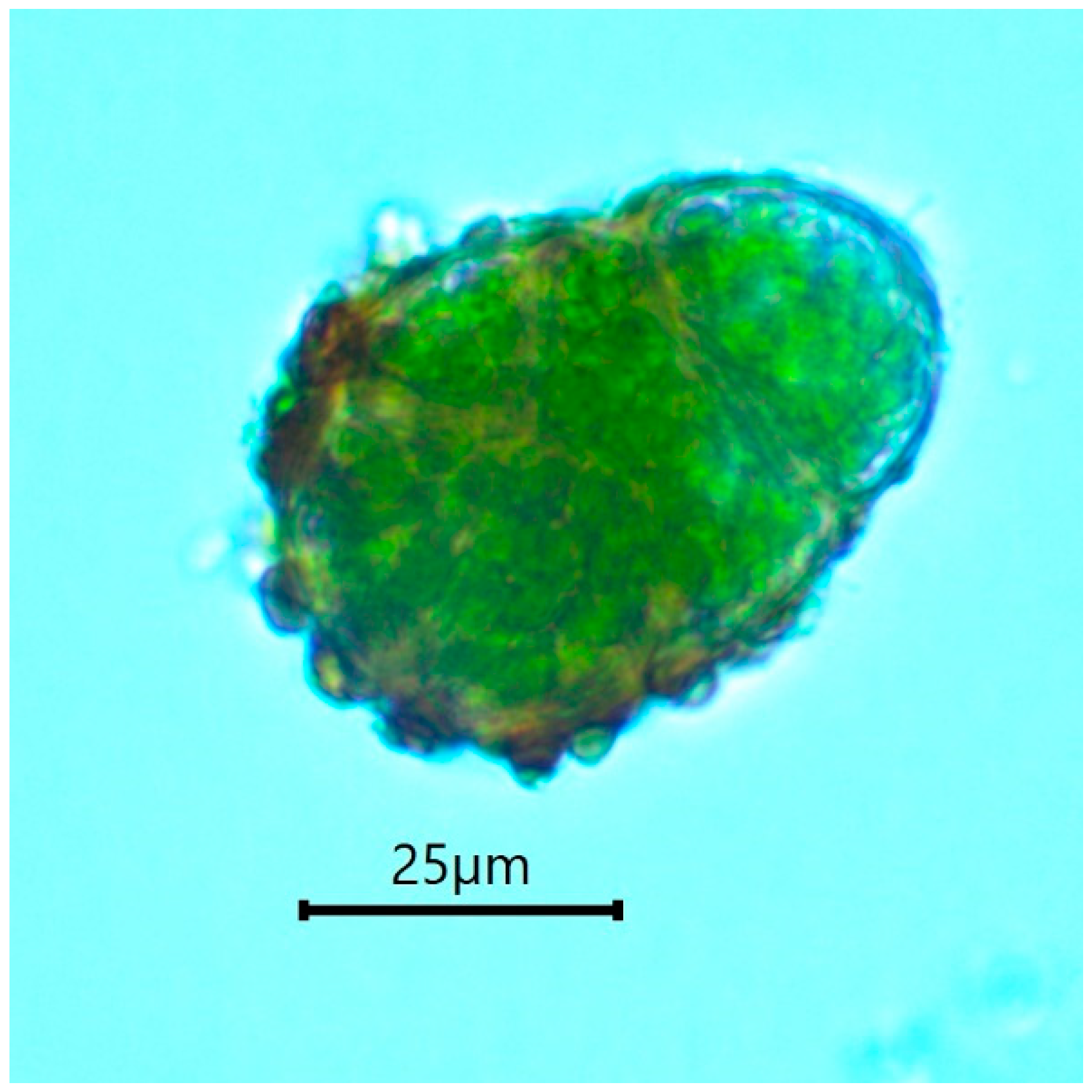
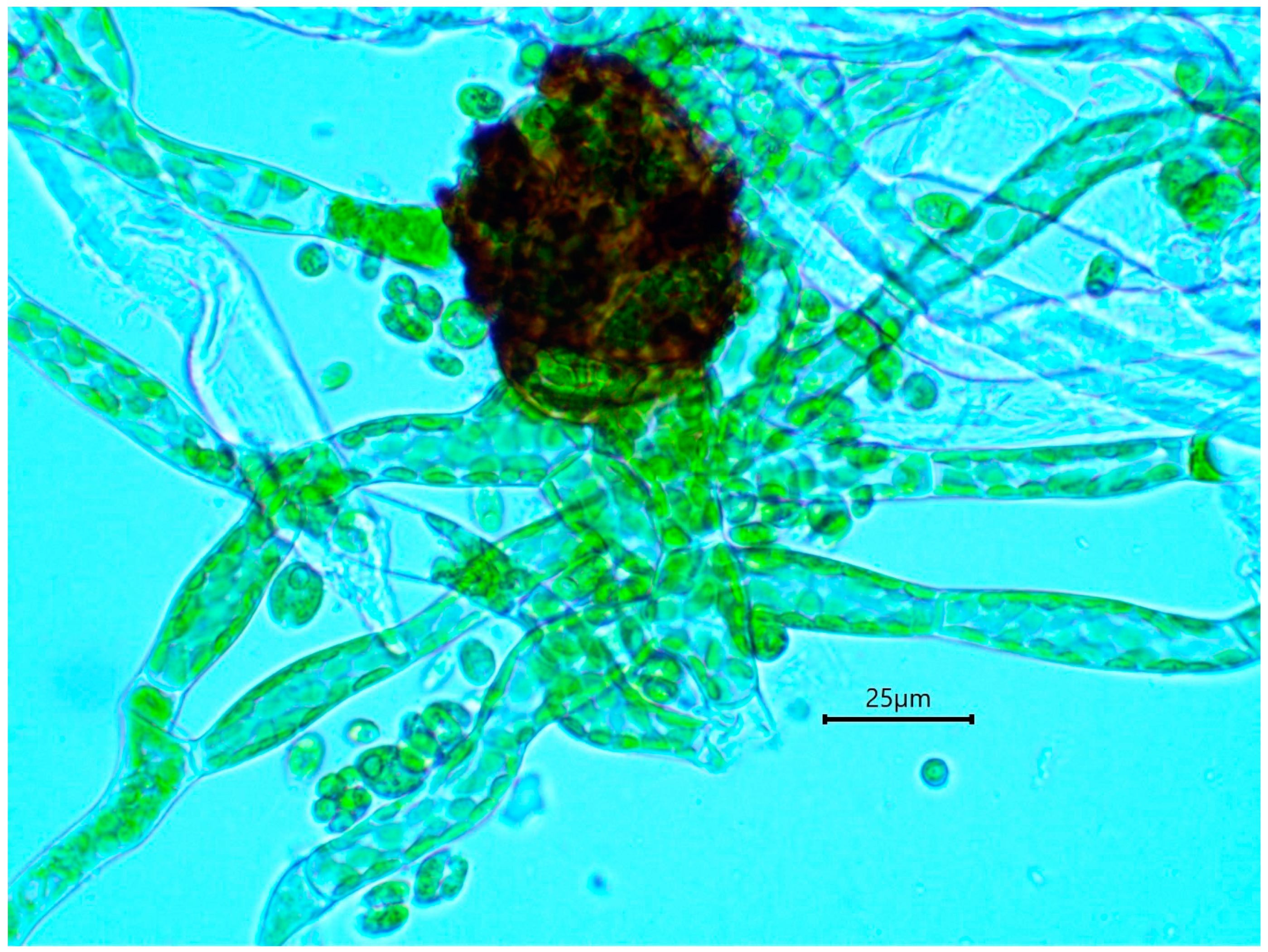
4.1. Practical Recognition of Buxbaumia viridis Non-Sporophytic Stage
4.2. Study Area
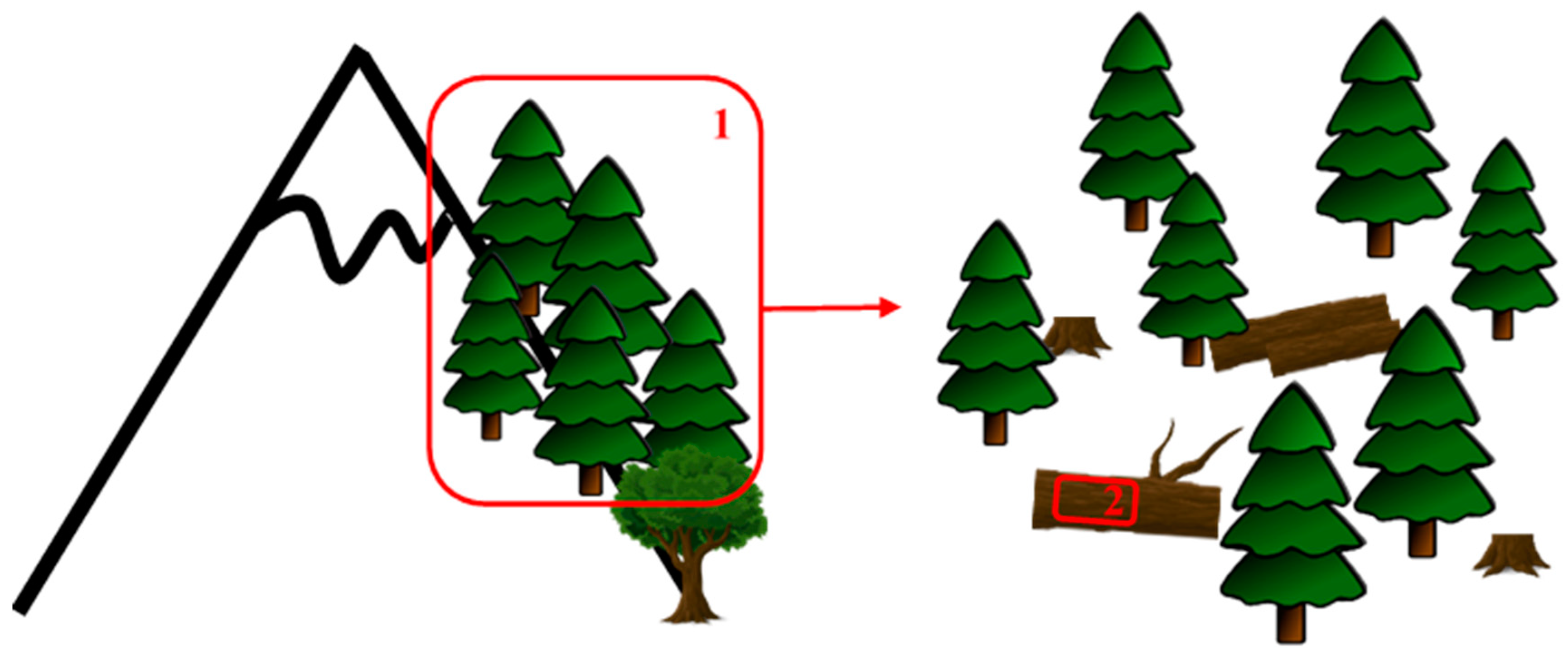
4.3. Habitat Scale
4.4. Microhabitat Scale
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oittinen, V. The distribution and ecology of Buxbaumia aphylla and B. viridis in Finland. Ann. Bot. Fenn. 1967, 4, 81–86. Available online: https://www.jstor.org/stable/23724310 (accessed on 5 May 2020).
- Dragićević, S.; Papp, B.; Erzberger, P. Distribution of Buxbaumia viridis (Moug. ex Lam. et DC.) Brid. ex Moug. et Nestl. (Bryophyta) in Montenegro. Acta Bot. Croat. 2012, 71, 365–370. Available online: https://hrcak.srce.hr/87490 (accessed on 5 May 2020).
- Spitale, D.; Mair, P. Predicting the distribution of a rare species of moss: The case of Buxbaumia viridis (Bryopsida, Buxbaumiaceae). Plant Biosyst. 2017, 151, 9–19. [Google Scholar] [CrossRef]
- Wiklund, K. Substratum preference, spore output and temporal variation in sporophyte production of the epixylic moss Buxbaumia viridis. J. Bryol. 2002, 24, 187–195. [Google Scholar] [CrossRef]
- Holá, E.; Vrba, J.; Linhartová, R.; Novozámská, E.; Zmrhalová, M.; Plášek, V.; Kučera, J. Thirteen years on the hunt for Buxbaumia viridis in the Czech Republic: Still on the tip of the iceberg? Acta Soc. Bot. Pol. 2014, 83, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Council of Europe. Bern Convention—Convention on the Conservation of European Wildlife and Natural Habitats. In Appendix I—Strictly Protected Flora Species; European Treaty Series No. 104; Council of Europe: Bern, Switzerland, 1979. [Google Scholar]
- Fudali, E.; Zubel, R.; Stebel, A.; Rusińska, A.; Górski, P.; Vončina, G.; Rosadziński, S.; Cykowska-Marzencka, B.; Staniaszek-Kik, M.; Wierzcholska, S.; et al. Contribution to the bryoflora of the roztocze national park (se poland)–bryophytes of the świerszcz river valley. Steciana 2015, 19, 39–54. [Google Scholar] [CrossRef]
- Callaghan, D.; Taylor, S. Classification of sporophyte stages in Buxbaumia viridis. Field Bryol. 2017, 117, 5–7. [Google Scholar]
- Gawryś, R.; Szulc, A. New location of the moss Buxbaumia viridis in the Białowieża Forest. J. For. Res. 2017, 78, 248–250. [Google Scholar] [CrossRef] [Green Version]
- Hodgetts, N.; Cálix, M.; Englefield, E.; Fettes, N.; García Criado, M.; Patin, L.; Nieto, A.; Bergamini, A.; Bisang, I.; Baisheva, E.; et al. A miniature World in Decline: European Red List of Mosses, Liverworts and Hornworts; IUCN: Brussels, Belgium, 2019; ISBN 978-2-8317-1993-1. [Google Scholar] [CrossRef] [Green Version]
- Siitonen, J. Forest Management, Coarse Woody Debris and Saproxylic Organisms: Fennoscandian Boreal Forests as an Example. Ecol. Bull. 2001, 49, 11–41. Available online: http://www.jstor.org/stable/20113262 (accessed on 5 May 2020).
- Benítez, Á.; Prieto, M.; Aragón, G. Large trees and dense canopies: Key factors for maintaining high epiphytic diversity on trunk bases (bryophytes and lichens) in tropical montane forests. Int. J. For. Res. 2015, 88, 521–527. [Google Scholar] [CrossRef] [Green Version]
- Wolf, T. Untersuchungen zu den Entwicklungsstadien von Buxbaumia viridis (Lam. & DC.) Moug. & Nestl. (Grünes Koboldmoos). Carolinea 2015, 73, 5–15. Available online: https://www.zobodat.at/pdf/Carolinea_73_0005-0015.pdf (accessed on 5 May 2020).
- Duckett, J.G.; Burch, J.; Fletcher, P.; Matcham, H.; Read, D.; Russell, A.; Pressel, S. In vitro cultivation of bryophytes: A review of practicalities, problems, progress and promise. J. Bryol. 2004, 26, 3–20. [Google Scholar] [CrossRef]
- Ebihara, A.; Nitta, J.H.; Matsumoto, Y.; Fukazawa, Y.; Kurihara, M.; Yokote, H.; Sakuma, K.; Azakami, O.; Hirayama, Y.; Imaichi, R. Growth dynamics of independent gametophytes of Pleurosoriopsis makinoi (Polypodiaceae). Bull. Nat. Mus. Nat. Sci. Ser. B (Bot.) 2019, 45, 77–86. [Google Scholar]
- Rumsey, F.J.; Jermy, A.C.; Sheffield, E. The independent gametophytic stage of Trichomanes speciosum Willd. (Hymenophyllaceae), the Killarney Fern and its distribution in the British Isles. Watsonia 1998, 22, 1–19. [Google Scholar]
- Herben, T. The role of reproduction for persistence of bryophyte populations in transient and stable habitats. J. Hattori Bot. Lab. 1994, 76, 115–126. [Google Scholar] [CrossRef]
- Mackensen, J.; Bauhus, J.; Webber, E. Decomposition rates of coarse woody debris—A review with particular emphasis on Australian tree species. Austr. J. Bot. 2003, 51, 27–37. [Google Scholar] [CrossRef]
- Kimmerer, R.W. Gathering Moss: A Natural and Cultural History of Mosses; Oregon State University: Corvallis, OR, USA, 2003; 168p, ISBN 9780870716416. [Google Scholar]
- Frey, W.; Kürschner, H. Asexual reproduction, habitat colonization and habitat maintenance in bryophytes. Flora-Morphol. Distrib. Funct. Ecol. Plants 2011, 206, 173–184. [Google Scholar] [CrossRef]
- Pohjamo, M.; Laaka-Lindberg, S.; Ovaskainen, O.; Korpelainen, H. Dispersal potential of spores and asexual propagules in the epixylic hepatic Anastrophyllum hellerianum. Evol. Ecol. 2006, 20, 415–430. [Google Scholar] [CrossRef]
- Rumsey, F.; Barrett, J.A.; Gibby, M.; Russell, S.; Vogel, J. Reproductive strategies and population structure in the endangered pteridophyte Trichomanes speciosum (Hymenophyllaceae: Pteridophyta). Fern Gaz. 2005, 17, 205–215. [Google Scholar]
- Chmielewski, M.W.; Eppley, S.M. Forest Passerines as a Novel Dispersal Vector of Viable Bryophyte Propagules. R. Soc. Lond. B Biol. Sci. 2019, 286, 20182253. [Google Scholar] [CrossRef] [Green Version]
- Táborská, M.; Přívětivý, T.; Vrška, T.; Ódor, P. Bryophytes associated with two tree species and different stages of decay in a natural fir-beech mixed forest in the Czech Republic. Preslia 2015, 87, 387–401. Available online: http://www.preslia.cz/P154Taborska.pdf (accessed on 5 May 2020).
- Wiklund, K. Phosphorus concentration and pH in decaying wood affect establishment of the red-listed moss Buxbaumia viridis. Can. J. Bot. 2003, 81, 541–549. [Google Scholar] [CrossRef]
- Goia, I.; Gafta, D. Beech versus spruce deadwood as forest microhabitat: Does it make any difference to bryophytes? Plant Biosyst. 2018, 153, 1–8. [Google Scholar] [CrossRef]
- Müller, J.; Boch, S.; Blaser, S.; Fischer, M.; Prati, D. Effects of forest management on bryophyte communities on deadwood. Nova Hedwig. 2015, 100, 423–438. [Google Scholar] [CrossRef]
- McCain, C.; Grytnes, J.A. Elevational gradients in species richness. In Encyclopedia of Life Sciences (ELS); John Wiley & Sons: Hoboken, NJ, USA, 2010. [Google Scholar] [CrossRef]
- Přívětivý, T.; Baldrian, P.; Šamonil, P.; Vrška, T. Deadwood Density and Moisture Variation in a Natural Temperate Spruce-Fir-Beech Forest. Preprints 2017. [Google Scholar] [CrossRef]
- Philippe, M. Occurrence of the moss Buxbaumia viridis (Bryopsida, Buxbaumiaceae) in Tatras National Park (Poland). In Bryophytes of the Polish Carpathians; Stebel, A., Ochyra, R., Eds.; Sorus: Poznań, Poland, 2004; pp. 29–36. [Google Scholar]
- Vončina, G.; Cykowska-Marzencka, B.; Chachuła, P. Rediscovery of Buxbaumia viridis (Bryophyta, Buxbaumiaceae) in the Tatra and Gorce in the Polish Western Carpathians; Sorus: Poznań, Poland, 2012; pp. 171–176. [Google Scholar]
- Kimmerer, R.W.; Young, C. Effect of Gap Size and Regeneration Niche on Species Coexistence in Bryophyte Communities. Bull. Torrey Bot. Club 1996, 123, 16–24. [Google Scholar] [CrossRef]
- Deme, J.; Erzberger, P.; Kovács, D.; Tóth, I.Z.; Csiky, J. Buxbaumia viridis (Moug. ex Lam. & DC.) Brid. Ex Moug. & Nestl. in Hungary predominantly terricolous and found in managed forests. Cryptogam. Bryol. 2020, 41, 89–103. [Google Scholar] [CrossRef]
- Bardat, J.; Hugonnot, V. Aperçu de la flore et de la végétation bryophytiques du site des Narces d’Issanlas (Ardèche-France), témoin exceptionnel d’une zone humide de moyenne montagne. Cryptogam. Bryol. 2002, 23, 51–72. [Google Scholar] [CrossRef]
- Philippe, M. Un cas de rareté paradoxale: Buxbaumia viridis (Musci, Bryales) en Franche-Comté. Nouv. Arch. Flore Jura. 2005, 3, 23–28. [Google Scholar]
- Lesica, P.; McCune, B.; Cooper, S.V.; Hong, W.S. Differences in lichen and bryophyte communities between old-growth and managed second-growth forests in the Swan Valley, Montana. Can. J. Bot. 1991, 69, 1745–1755. [Google Scholar] [CrossRef]
- Bunnell, F.L.; Houde, I. Down wood and biodiversity—Implications to forest practices. Environ. Rev. 2010, 18, 397–421. [Google Scholar] [CrossRef]
- Agresti, A. An Introduction to Categorical Data Analysis; Wiley: Hoboken, NJ, USA, 2007; ISBN 978-0471226185. [Google Scholar]








| (a) Buxbaumia viridis (Sterile Protonema + Sporophyte) | (b) Sporophyte | |||||
|---|---|---|---|---|---|---|
| Predictor | β | SE | p (Wald Test) | β | SE | p (Wald Test) |
| (Intercept) | −5.282 | 0.946 | <0.001 | −10.023 | 1.853 | <0.001 |
| Mixed forest | 3.426 | 0.790 | <0.001 | |||
| Coniferous forest | 3.432 | 0.699 | <0.001 | |||
| Deadwood surface | 0.253 | 0.049 | <0.001 | 0.096 | 0.043 | 0.026 |
| Elevation | 0.007 | 0.001 | <0.001 | |||
| Northness | 1.250 | 0.522 | 0.017 | |||
| Observations | 135 | 135 | ||||
| R² Tjur | 0.555 | 0.418 | ||||
| Hoslem–Lemeshow p (Goodness-of-fit test) | 0.190 | 0.854 | ||||
| (a) | ||||
|---|---|---|---|---|
| Variable | Mean ± SD | Min | Max | |
| Canopy opening (%) | ||||
| All substrates | 12.0 ± 4.7 | 2.5 | 24.5 | |
| with B. viridis | 11.0 ± 4.6 | 2.5 | 22.5 | |
| with sporophytes | 12.8 ± 4.3 | 5.5 | 22.5 | |
| Slope (°) | ||||
| All substrates | 35 ± 29 | 0 | 90 | |
| With B. viridis | 34 ± 29 | 0 | 90 | |
| With sporophytes | 32 ± 30 | 0 | 90 | |
| Northness | ||||
| All substrates | 0.28 ± 0.66 | −1 | 1 | |
| With B. viridis | 0.29 ± 0.66 | −1 | 1 | |
| With sporophytes | 0.34 ± 0.61 | −0.92 | 1 | |
| Bryophyte cover (%) | ||||
| All substrates | 26 ± 21 | 0 | 98 | |
| With B. viridis | 28 ± 21 | 0 | 88 | |
| With sporophytes | 30 ± 22 | 0 | 88 | |
| Diameter (cm) | ||||
| All substrates | 18.5 ± 11.2 | 10.0 | 90.0 | |
| With B. viridis | 19.0 ± 11.8 | 10.0 | 90.0 | |
| With sporophytes | 16.4 ± 6.4 | 10.0 | 90.0 | |
| (b) | ||||
| Wood Decomposition | No All substrates | No with B. viridis | No with sporophytes | |
| Stage 2 | 25 | 0 | 0 | |
| Stage 3 | 52 | 31 | 2 | |
| Stage 4 | 134 | 124 | 42 | |
| Stage 5 | 64 | 62 | 17 | |
| Total | 275 | 217 | 61 | |
| (a) Gemmae Occupancy | (b) Number of Sporophytes | |||||
|---|---|---|---|---|---|---|
| Predictor | β | SE | p (t-Test) | β | SE | p (t-Test) |
| (Intercept) | 0.465 | 0.348 | <0.001 | 0.020 | 1.025 | <0.001 |
| Decomposition stage 3 | 1.539 | 0.317 | <0.001 | |||
| Decomposition stage 4 | 2.654 | 0.287 | <0.001 | 15.288 | 0.940 | 0.004 |
| Decomposition stage 5 | 2.862 | 0.310 | <0.001 | 14.457 | 0.956 | 0.006 |
| Canopy opening | −0.034 | 0.017 | 0.045 | 1.164 | 0.031 | <0.001 |
| Observations | 275 | 217 | ||||
| R² Nagelkerke | 0.602 | 0.487 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guillet, A.; Hugonnot, V.; Pépin, F. The Habitat of the Neglected Independent Protonemal Stage of Buxbaumia viridis. Plants 2021, 10, 83. https://doi.org/10.3390/plants10010083
Guillet A, Hugonnot V, Pépin F. The Habitat of the Neglected Independent Protonemal Stage of Buxbaumia viridis. Plants. 2021; 10(1):83. https://doi.org/10.3390/plants10010083
Chicago/Turabian StyleGuillet, Ameline, Vincent Hugonnot, and Florine Pépin. 2021. "The Habitat of the Neglected Independent Protonemal Stage of Buxbaumia viridis" Plants 10, no. 1: 83. https://doi.org/10.3390/plants10010083
APA StyleGuillet, A., Hugonnot, V., & Pépin, F. (2021). The Habitat of the Neglected Independent Protonemal Stage of Buxbaumia viridis. Plants, 10(1), 83. https://doi.org/10.3390/plants10010083