
Genome-Wide Transcriptomic and Proteomic Exploration of Molecular Regulations in Quinoa Responses to Ethylene and Salt Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material Treatment and Sample Collection
2.2. Transcriptome Sequencing and Data Analysis
2.3. Protein Extraction, TMT Labeling, and Proteomics Analysis
2.4. Correlation Analysis between Proteomic and Transcriptomic Results
2.5. Quantitative Real Time PCR (qRT-PCR) Analysis
2.6. Physiological Indexes Detection
2.7. Statistical Analyses
3. Results
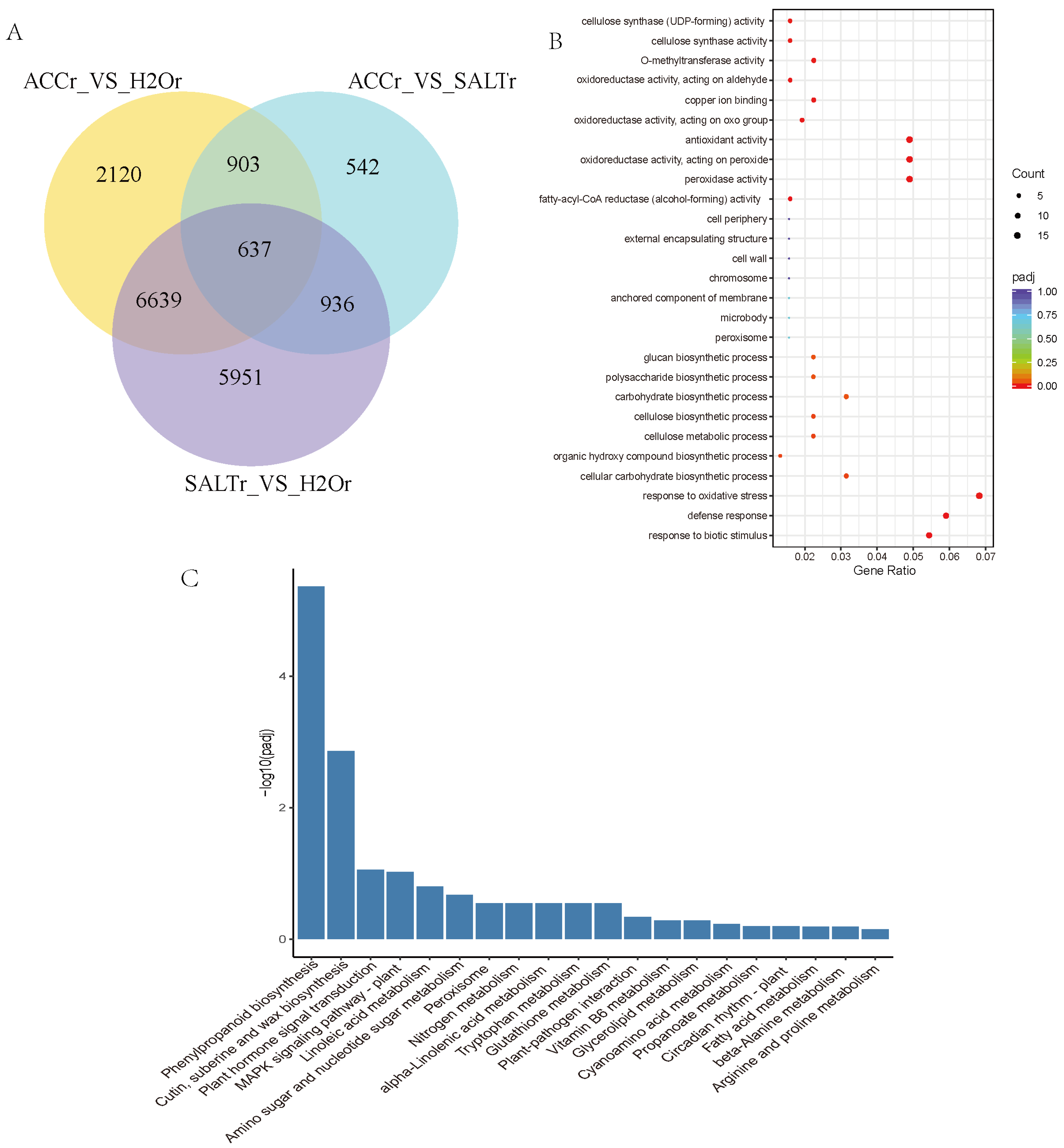
3.1. Gene Identification and DEGs Analysis in Transcriptome
3.2. DEGs Detection in Ethylene and Salt Responses of Quinoa
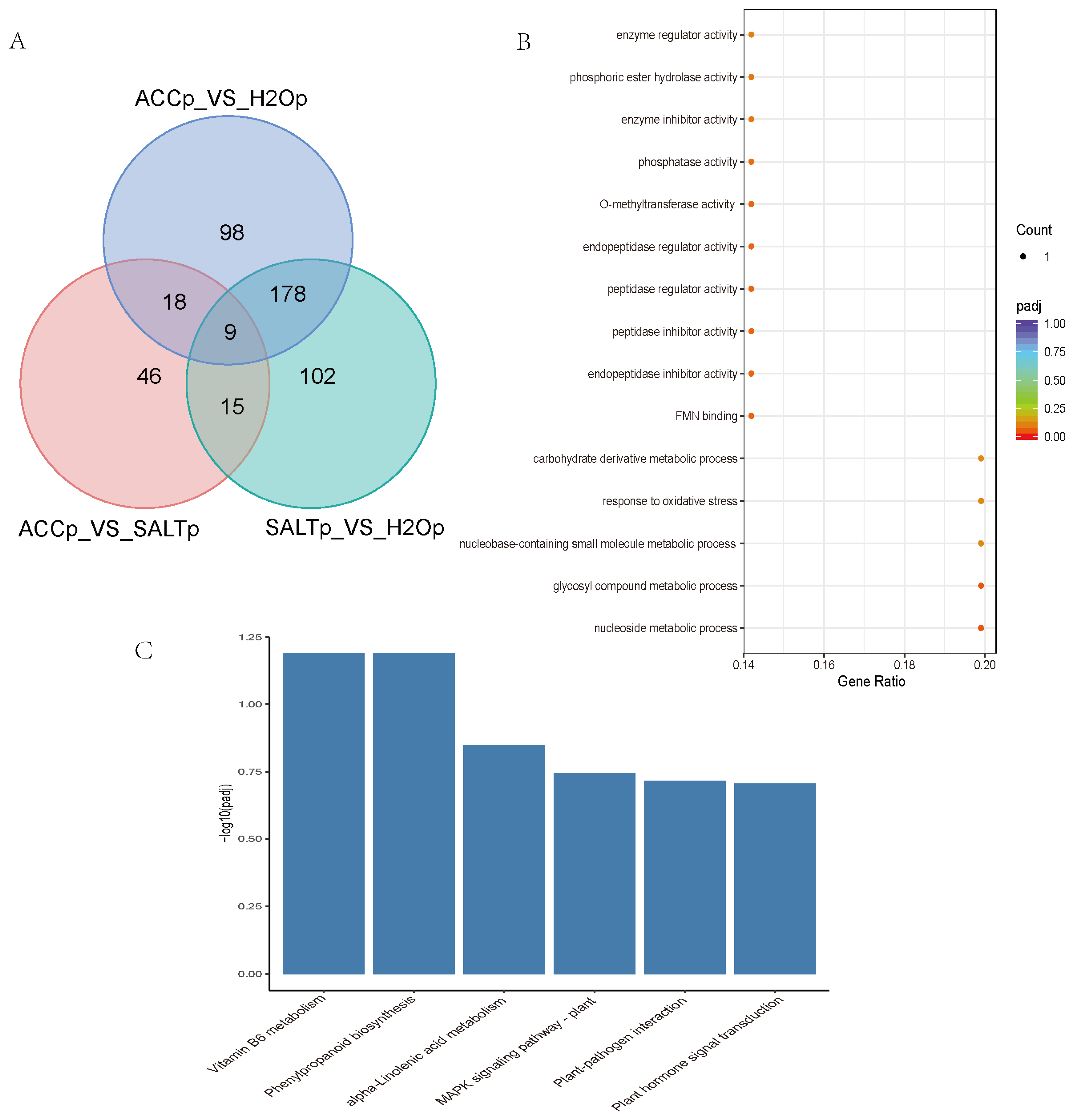
3.3. Protein Identification and DEPs Analysis
3.4. DEPs Analysis in Ethylene Regulated Salt Responses
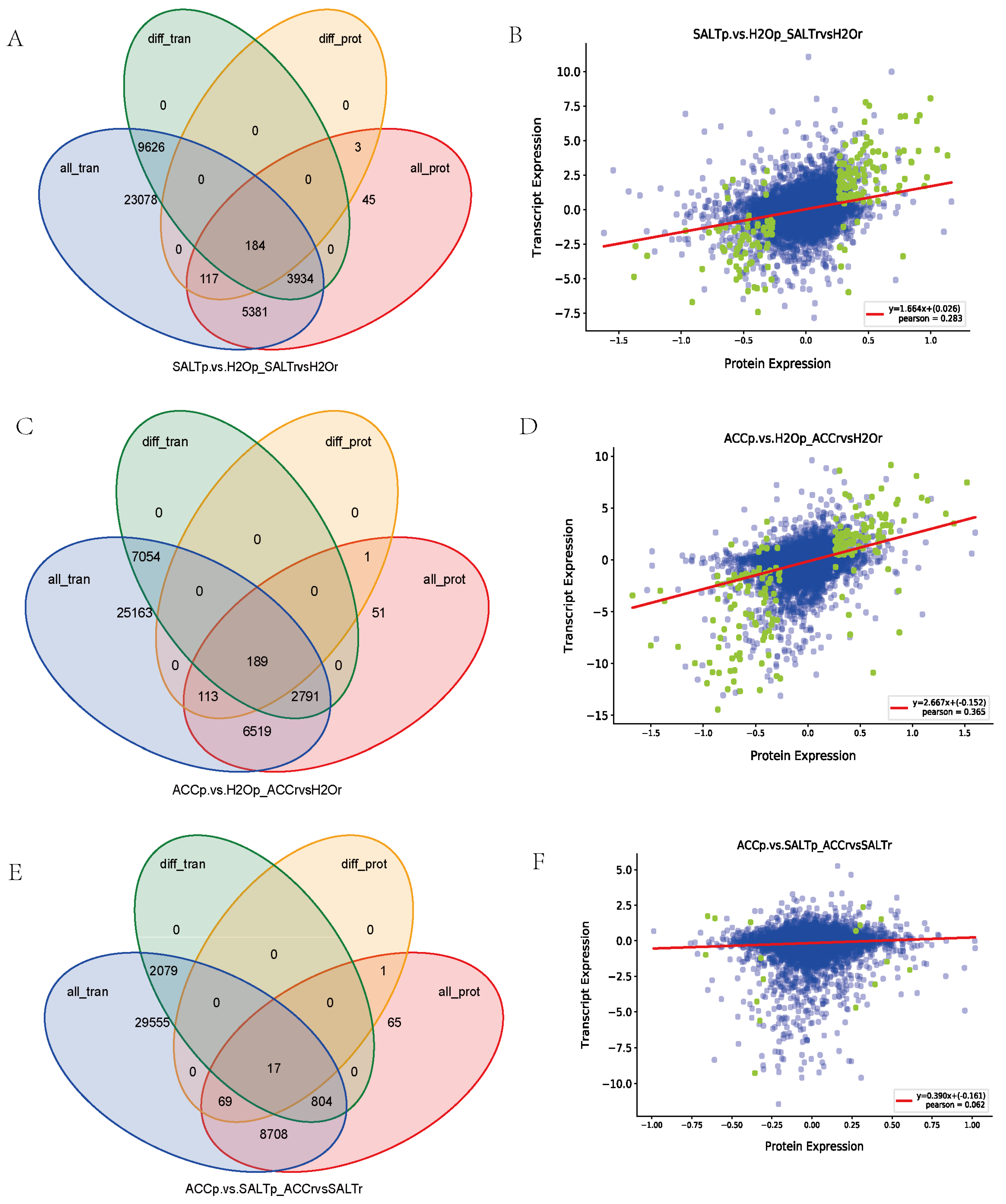
3.5. Correlation between the Proteomic and Transcriptomic Results
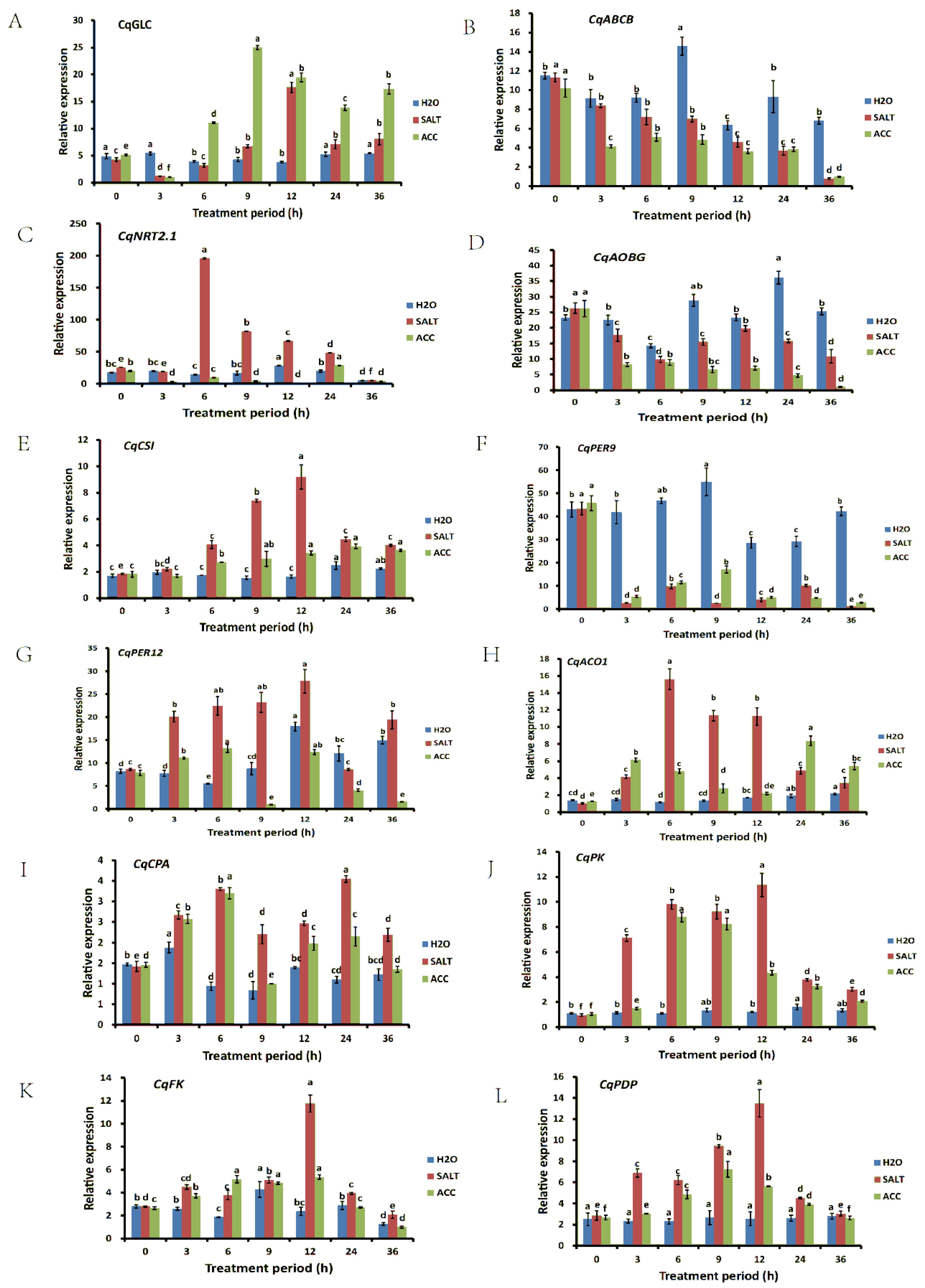
3.6. Verification of RNA-seq Results by qRT-PCR
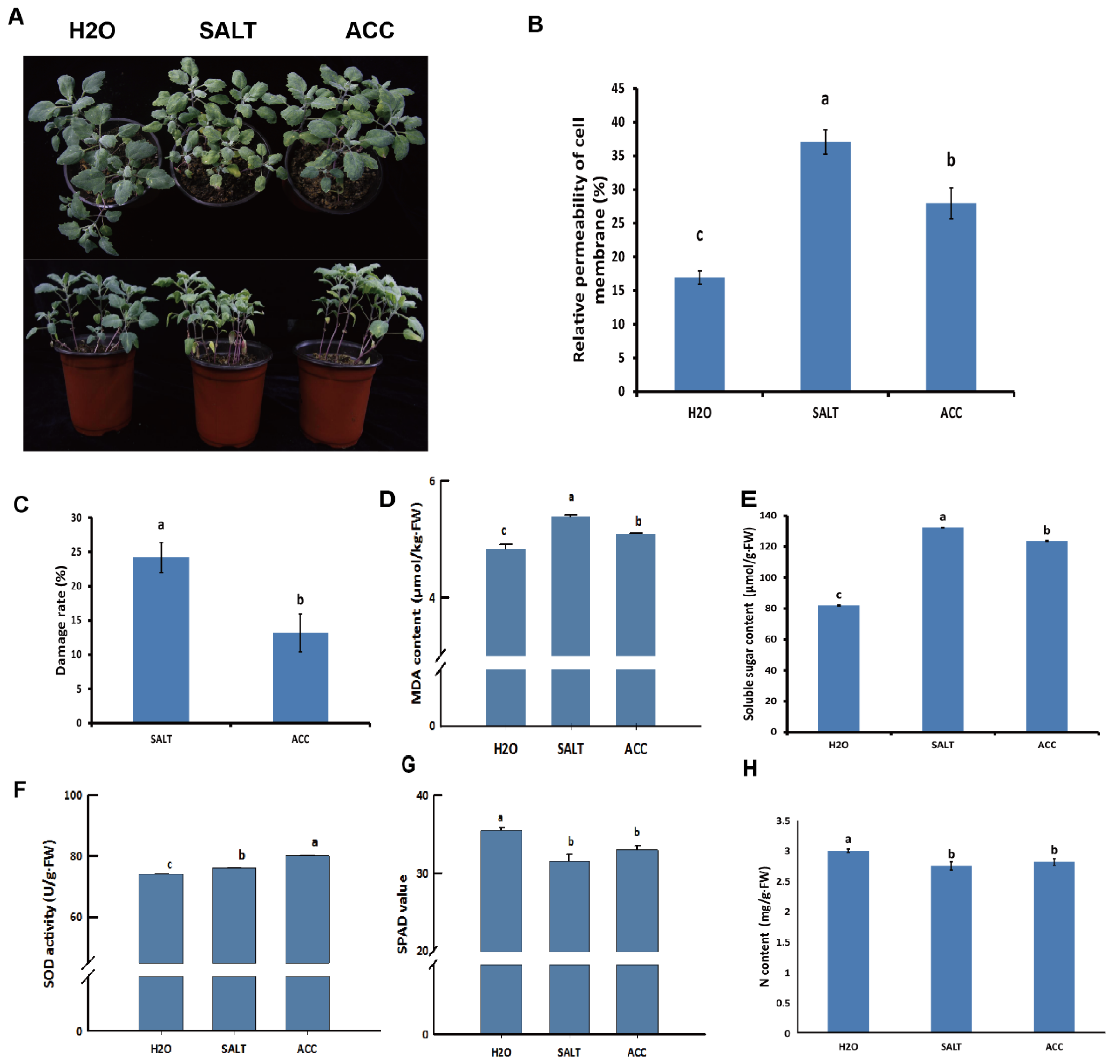
3.7. Physiological Alterations by Ethylene and Salt Stress
4. Discussion
4.1. Plant Hormones Play Regulatory Roles in Quinoa Responses to Ethylene and Salt Stress
4.2. ROS Scavenging Enzymes Function in Quinoa Responses to Ethylene and Salt Stress
4.3. Osmotic Adjustment Is Important for Quinoa Responses to Ethylene and Salt Stress
4.4. Cell Wall Structural Proteins Respond to Ethylene and Salt Stress in Quinoa
4.5. Secondary Metabolism-Associated Proteins Respond to Ethylene and Salt Stress in Quinoa
4.6. Early Response Genes and Late Response Genes in Quinoa
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jacobsen, S.E.; Mujica, A.; Jensen, C.R. The resistance of quinoa (Chenopodium quinoa Willd.) to adverse abiotic factors. Food Rev. Int. 2003, 19, 99–109. [Google Scholar] [CrossRef]
- Kolano, B.; McCann, J.; Orzechowska, M.; Siwinska, D.; Temsch, E.; Weiss-Schneeweiss, H. Molecular and cytogenetic evidence for an allotetraploid origin of Chenopodium quinoa and C. berlandieri (Amaranthaceae). Mol. Phylogenet. Evol. 2016, 100, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Zurita-Silva, A.; Fuentes, F.; Zamora, P.; Jacobsen, S.E.; Schwember, A.R. Breeding quinoa (Chenopodium quinoa Willd.): Potential and perspectives. Mol. Breed. 2014, 34, 13–30. [Google Scholar] [CrossRef]
- Vega-Galvez, A.; Miranda, M.; Vergara, J.; Uribe, E.; Puente, L.; Martinez, E.A. Nutrition facts and functional potential of quinoa (Chenopodium quinoa willd.) an ancient Andean grain: A review. J. Sci. Food Agric. 2010, 90, 2541–2547. [Google Scholar] [CrossRef]
- Bhargava, A.; Shukla, S.; Ohri, D. Chenopodium quinoa. An indian perspective. Ind. Crops Prod. 2006, 23, 73–87. [Google Scholar] [CrossRef]
- Ruiz, K.B.; Biondi, S.; Oses, R.; Acuna-Rodriguez, I.S.; Antognoni, F.; Martinez-Mosqueira, E.A.; Coulibaly, A.; Canahua-Murillo, A.; Pinto, M.; Zurita-Silva, A.; et al. Quinoa biodiversity and sustainability for food security under climate change. Agron. Sustain. Dev. 2014, 34, 349–359. [Google Scholar] [CrossRef] [Green Version]
- Simnadis, T.G.; Tapsell, L.C.; Beck, E.J. Physiological effects associated with quinoa consumption and implications for research involving humans: A review. Plant Foods Hum. Nutr. 2015, 70, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Angeli, V.; Silva, P.M.; Massuela, D.C.; Khan, M.W.; Hamar, A.; Khajehei, F.; Graeff-Honninger, S.; Piatti, C. Quinoa (Chenopodium quinoa Willd.): An overview of the potentials of the "golden grain" and socio-economic and environmental aspects of its cultivation and marketization. Foods 2020, 9, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazile, D.; Jacobsen, S.E.; Verniau, A. The global expansion of quinoa: Trends and limits. Front. Plant Sci. 2016, 7, 622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, C.; Chen, A.; Xiao, L.; Muller, H.M.; Ache, P.; Haberer, G.; Zhang, M.; Jia, W.; Deng, P.; Huang, R.; et al. A high-quality genome assembly of quinoa provides insights into the molecular basis of salt bladder-based salinity tolerance and the exceptional nutritional value. Cell Res. 2017, 27, 1327–1340. [Google Scholar] [CrossRef]
- Rozema, J.; Flowers, T. Ecology. Crops for a salinized world. Science 2008, 322, 1478–1480. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity tolerance of crops—What is the cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [Green Version]
- van Zelm, E.; Zhang, Y.X.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, K.B.; Aloisi, I.; Del Duca, S.; Canelo, V.; Torrigiani, P.; Silva, H.; Biondi, S. Salares versus coastal ecotypes of quinoa: Salinity responses in Chilean landraces from contrasting habitats. Plant Physiol. Biochem. 2016, 101, 1–13. [Google Scholar] [CrossRef]
- Iqbal, S.; Basra, S.M.A.; Afzal, I.; Wahid, A. Exploring potential of well adapted quinoa lines for salt tolerance. Int. J. Agric. Biol. 2017, 19, 933–940. [Google Scholar] [CrossRef]
- Saleem, M.A.; Basra, S.M.A.; Afzal, I.; Hafeez-ur-Rehman; Iqbal, S.; Saddiq, M.S.; Naz, S. Exploring the potential of quinoa accessions for salt tolerance in soilless culture. Int. J. Agric. Biol. 2017, 19, 233–240. [Google Scholar] [CrossRef]
- Panuccio, M.R.; Jacobsen, S.E.; Akhtar, S.S.; Muscolo, A. Effect of saline water on seed germination and early seedling growth of the halophyte quinoa. AoB Plants 2014, 6, plu047. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.Q.; Gao, Q. Comparative physiological and biochemical mechanisms of salt tolerance in five contrasting highland quinoa cultivars. BMC Plant Biol. 2020, 20, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, S.; Ma, J.; Zhang, L.; Chen, F.; Sang, Z.; Jia, Z.; Ma, L. De novo transcriptome sequencing and gene expression profiling of Magnolia wufengensis in response to cold stress. BMC Plant Biol. 2019, 19, 321. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Yan, Y.; Hou, X.; He, Y.; Wei, Y.; Yang, G.; He, G.; Peng, M. TaPP2C1, a group F2 protein phosphatase 2C gene, confers resistance to salt stress in transgenic tobacco. PLoS ONE 2015, 10, e0129589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, P.; Gu, M. Transcriptome analysis and differential gene expression profiling of two contrasting quinoa genotypes in response to salt stress. BMC Plant Biol. 2020, 20, 568. [Google Scholar] [CrossRef]
- Achard, P.; Cheng, H.; De Grauwe, L.; Decat, J.; Schoutteten, H.; Moritz, T.; Van Der Straeten, D.; Peng, J.; Harberd, N.P. Integration of plant responses to environmentally activated phytohormonal signals. Science 2006, 311, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.R.; Chen, S.Y.; Zhang, J.S. Ethylene signaling regulates salt stress response: An overview. Plant Signal. Behav. 2008, 3, 761–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, T.; Li, K.; Li, X. Genetic analysis of involvement of ETR1 in plant response to salt and osmotic stress. Plant Growth Regul. 2008, 54, 261–269. [Google Scholar] [CrossRef]
- Ma, Q.; Zhou, H.; Sui, X.; Su, C.; Yu, Y.; Yang, Y.; Dong, C. Generation of new salt-tolerant wheat lines and transcriptomic exploration of the responsive genes to ethylene and salt stress. Plant Growth Regul. 2021, 94, 33–48. [Google Scholar] [CrossRef]
- Wang, H.; Wang, F.; Zheng, F.; Wang, L.; Pei, H.; Dong, C. Ethylene-insensitive mutants of Nicotiana tabacum exhibit drought stress resistance. Plant Growth Regul. 2016, 79, 107–117. [Google Scholar] [CrossRef]
- Riyazuddin, R.; Verma, R.; Singh, K.; Nisha, N.; Keisham, M.; Bhati, K.K.; Kim, S.T.; Gupta, R. Ethylene: A master regulator of salinity stress tolerance in plants. Biomolecules 2020, 10, 959. [Google Scholar] [CrossRef]
- Sun, Y.; Lindberg, S.; Shabala, L.; Morgan, S.; Shabala, S.; Jacobsen, S.E. A comparative analysis of cytosolic Na+ changes under salinity between halophyte quinoa (Chenopodium quinoa) and glycophyte pea (Piswn sativwn). Environ. Exp. Bot. 2017, 141, 154–160. [Google Scholar] [CrossRef]
- Eisa, S.S.; Eid, M.A.; Abd, E.S.; Hussin, S.A.; Abdel-Ati, A.A.; El-Bordeny, N.E.; Ali, S.H.; Al-Sayed, H.M.A.; Lotfy, M.E.; Masoud, A.M.; et al. Chenopodium quinoa Willd. A new cash crop halophyte for saline regions of Egypt. Aust. J. Crop. Sci. 2017, 11, 343–351. [Google Scholar] [CrossRef]
- Ruiz, K.B.; Maldonado, J.; Biondi, S.; Silva, H. RNA-seq analysis of salt-stressed versus non salt-sressed transcriptomes of Chenopodium quinoa Landrace R49. Genes 2019, 10, 1042. [Google Scholar] [CrossRef] [Green Version]
- Schmockel, S.M.; Lightfoot, D.J.; Razali, R.; Tester, M.; Jarvis, D.E. Identification of putative transmembrane proteins involved in salinity tolerance in Chenopodium quinoa by integrating physiological data, RNAseq, and SNP analyses. Front. Plant Sci. 2017, 8, 1023. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, D.E.; Ho, Y.S.; Lightfoot, D.J.; Schmockel, S.M.; Li, B.; Borm, T.J.; Ohyanagi, H.; Mineta, K.; Michell, C.T.; Saber, N.; et al. The genome of Chenopodium quinoa. Nature 2017, 542, 307–312. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Shi, C.H.; Su, C.X.; Liu, Y.G. Complementary analyses of the transcriptome and iTRAQ proteome revealed mechanism of ethylene dependent salt response in bread wheat (Triticum aestivum L.). Food Chem. 2020, 325, 126866. [Google Scholar] [CrossRef]
- Ma, J.; Chen, T.; Wu, S.; Yang, C.; Bai, M.; Shu, K.; Li, K.; Zhang, G.; Jin, Z.; He, F.; et al. iProX: An integrated proteome resource. Nucleic Acids Res. 2019, 47, D1211–D1217. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.P.; Yu, Y.C.; Zhang, T.; Ma, Q.; Yang, H.B. Effects of ozone water irrigation and spraying on physiological characteristics and gene expression of tomato seedlings. Hortic. Res. 2021, 8, 180. [Google Scholar] [CrossRef] [PubMed]
- Graf, B.L.; Rojas-Silva, P.; Rojo, L.E.; Delatorre-Herrera, J.; Baldeon, M.E.; Raskin, I. Innovations in health value and functional food development of quinoa (Chenopodium quinoa Willd.). Compr. Rev. Food Sci. Food Saf. 2015, 14, 431–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Jiao, Y.; Xie, N.K.; Guo, Y.M.; Zhang, F.; Xiang, Z.P.; Wang, R.; Wang, F.; Gao, Q.M.; Tian, L.F.; et al. OsNCED5, a 9-cis-epoxycarotenoid dioxygenase gene, regulates salt and water stress tolerance and leaf senescence in rice. Plant Sci. 2019, 287, 110188. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, K.B.; Rapparini, F.; Bertazza, G.; Silva, H.; Torrigiani, P.; Biondi, S. Comparing salt-induced responses at the transcript level in a salares and coastal-lowlands landrace of quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 2017, 140, 150. [Google Scholar] [CrossRef]
- Mortier, V.; Wasson, A.; Jaworek, P.; De Keyser, A.; Decroos, M.; Holsters, M.; Tarkowski, P.; Mathesius, U.; Goormachtig, S. Role of LONELY GUY genes in indeterminate nodulation on Medicago truncatula. New Phytol. 2014, 202, 582–593. [Google Scholar] [CrossRef]
- Almagro, L.; Gomez Ros, L.V.; Belchi-Navarro, S.; Bru, R.; Ros Barcelo, A.; Pedreno, M.A. Class III peroxidases in plant defence reactions. J. Exp. Bot. 2009, 60, 377–390. [Google Scholar] [CrossRef] [Green Version]
- Passardi, F.; Cosio, C.; Penel, C.; Dunand, C. Peroxidases have more functions than a Swiss army knife. Plant Cell Rep. 2005, 24, 255–265. [Google Scholar] [CrossRef]
- Shigeto, J.; Tsutsumi, Y. Diverse functions and reactions of class III peroxidases. New Phytol. 2016, 209, 1395–1402. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Han, X.M.; Ren, L.L.; Yang, H.L.; Zeng, Q.Y. Functional divergence of the glutathione S-transferase supergene family in Physcomitrella patens reveals complex patterns of large gene family evolution in land plants. Plant Physiol. 2013, 161, 773–786. [Google Scholar] [CrossRef] [Green Version]
- Pereira, E.; Cadavez, V.; Barros, L.; Encina-Zelada, C.; Stojkovic, D.; Sokovic, M.; Calhelha, R.C.; Gonzales-Barron, U.; Ferreira, I. Chenopodium quinoa Willd. (quinoa) grains: A good source of phenolic compounds. Food Res. Int. 2020, 137, 109574. [Google Scholar] [CrossRef]
- Bohm, J.; Messerer, M.; Muller, H.M.; Scholz-Starke, J.; Gradogna, A.; Scherzer, S.; Maierhofer, T.; Bazihizina, N.; Zhang, H.; Stigloher, C.; et al. Understanding the molecular basis of salt sequestration in epidermal bladder cells of Chenopodium quinoa. Curr. Biol. 2018, 28, 3075–3085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarabi, B.; Bolandnazar, S.; Ghaderi, N.; Ghashghaie, J. Genotypic differences in physiological and biochemical responses to salinity stress in melon (Cucumis melo L.) plants: Prospects for selection of salt tolerant landraces. Plant Physiol. Biochem. 2017, 119, 294–311. [Google Scholar] [CrossRef] [PubMed]
- Ruffino, A.; Rosa, M.; Hilal, M.; Gonzalez, J.; Prado, F. The role of cotyledon metabolism in the establishment of quinoa (Chenopodium quinoa) seedlings growing under salinity. Plant Soil 2010, 326, 213–224. [Google Scholar] [CrossRef]
- Touchette, B.W. Seagrass-salinity interactions: Physiological mechanisms used by submersed marine angiosperms for a life at sea. J. Exp. Mar. Boil. Ecol. 2007, 350, 194–215. [Google Scholar] [CrossRef]
- Wang, G.L.; Ren, X.Q.; Liu, J.X.; Yang, F.; Wang, Y.P.; Xiong, A.S. Transcript profiling reveals an important role of cell wall remodeling and hormone signaling under salt stress in garlic. Plant Physiol. Biochem. 2019, 135, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, V.; Selbig, J.; Scheible, W.R. Involvement of TBL/DUF231 proteins into cell wall biology. Plant Signal. Behav. 2010, 5, 1057–1059. [Google Scholar] [CrossRef] [Green Version]
- Endler, A.; Kesten, C.; Schneider, R.; Zhang, Y.; Ivakov, A.; Froehlich, A.; Funke, N.; Persson, S. A mechanism for sustained cellulose synthesis during salt stress. Cell 2015, 162, 1353–1364. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Zhang, Z.; Chen, Z.; Cao, B.; Xu, K. Comparative transcriptome analysis of two contrasting Chinese cabbage (Brassica rapa L.) genotypes reveals that ion homeostasis is a crucial biological pathway involved in the rapid adaptive response to salt stress. Front. Plant Sci. 2021, 12, 683891. [Google Scholar] [CrossRef]
- Imamura, T.; Takagi, H.; Miyazato, A.; Ohki, S.; Mizukoshi, H.; Mori, M. Isolation and characterization of the betalain biosynthesis gene involved in hypocotyl pigmentation of the allotetraploid Chenopodium quinoa. Biochem. Biophys. Res. Commun. 2018, 496, 280–286. [Google Scholar] [CrossRef]
- Jain, G.; Schwinn, K.E.; Gould, K.S. Betalain induction by l-DOPA application confers photoprotection to saline-exposed leaves of Disphyma australe. New Phytol 2015, 207, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Koduri, P.K.; Gordon, G.S.; Barker, E.I.; Colpitts, C.C.; Ashton, N.W.; Suh, D.Y. Genome-wide analysis of the chalcone synthase superfamily genes of Physcomitrella patens. Plant Mol. Biol. 2010, 72, 247–263. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.P.; Huang, L.M.; Chen, L.O.; Chan, M.T.; Shaw, J.F. Genome-wide analysis of GDSL-type esterases/lipases in Arabidopsis. Plant Mol. Biol. 2017, 95, 181–197. [Google Scholar] [CrossRef]
- Qin, H.; Wang, Y.Y.; Wang, J.; Liu, H.; Zhao, H.; Deng, Z.A.; Zhang, Z.L.; Huang, R.F.; Zhang, Z.J. Knocking down the expression of GMPase gene OsVTC1-1 decreases salt tolerance of rice at seedling and reproductive stages. PLoS ONE 2016, 11, e0168650. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.G.; Wang, B.; Jin, S.H.; Qu, X.X.; Li, Y.J.; Hou, B.K. Ectopic expression of Arabidopsis glycosyltransferase UGT85A5 enhances salt stress tolerance in tobacco. PLoS ONE 2013, 8, e59924. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Carrozzi, V.R.; Nazarian, A.A.; Li, C.C.; Gore, S.L.; Sridharan, R.; Imbalzano, A.N.; Smale, S.T. Selective and antagonistic functions of SWI/SNF and Mi-2beta nucleosome remodeling complexes during an inflammatory response. Genes Dev. 2006, 20, 282–296. [Google Scholar] [CrossRef] [Green Version]
- Smale, S.T. Selective transcription in response to an inflammatory stimulus. Cell 2010, 140, 833–844. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviations | Detailed Information | Omics Used in |
|---|---|---|
| H2Op | Water-treated seedlings | proteomics |
| SALTp | Salt-treated seedlings | proteomics |
| ACCp | Salt- and ACC-treated seedlings | proteomics |
| H2Or | Water-treated seedlings | transcriptomics |
| SALTr | Salt-treated seedlings | transcriptomics |
| ACCr | Salt- and ACC-treated seedlings | transcriptomics |
| Gene ID | Symbol | Description | Function Category | Classification |
|---|---|---|---|---|
| LOC110729845 | ERF | ethylene-responsive transcription factor | plant hormone signaling | late response gene |
| LOC110730331 | ERF104 | ethylene-responsive transcription factor 104 | plant hormone signaling | late response gene |
| LOC110719638 | ERF2 | ethylene-responsive transcription factor 2 | plant hormone signaling | late response gene |
| LOC110719716 | ARF9 | auxin response factor 9 | plant hormone signaling | late response gene |
| LOC110715799 | ABP19a | auxin-binding protein 19a | plant hormone signaling | late response gene |
| LOC110710755 | PYL4 | abscisic acid receptor PYL4 | plant hormone signaling | late response gene |
| LOC110715081 | JIP | jasmonate-induced protein | plant hormone signaling | late response gene |
| LOC110711071 | JIP | jasmonate-induced protein | plant hormone signaling | late response gene |
| LOC110733576 | JIP | jasmonate-induced protein | plant hormone signaling | late response gene |
| LOC110738584 | LOG5 | cytokinin riboside 5′-monophosphate phosphoribohydrolase | plant hormone biosynthesis | late response gene |
| LOC110724460 | GST | glutathione S-transferase | ROS scavenging | late response gene |
| LOC110696392 | GST | glutathione S-transferase | ROS scavenging | late response gene |
| LOC110724461 | GST | glutathione S-transferase | ROS scavenging | late response gene |
| LOC110728060 | GST | glutathione S-transferase | ROS scavenging | late response gene |
| LOC110711174 | GST3 | microsomal glutathione S-transferase 3 | ROS scavenging | late response gene |
| LOC110739278 | GST | glutathione S-transferase | ROS scavenging | early response gene |
| LOC110713696 | GST | glutathione S-transferase | ROS scavenging | post-transcriptional modification |
| LOC110727188 | GST | glutathione S-transferase | ROS scavenging | post-transcriptional modification |
| LOC110682117 | POD64 | peroxidase 64 | ROS scavenging | post-transcriptional modification |
| LOC110682546 | POD72 | peroxidase 72 | ROS scavenging | late response gene |
| LOC110685850 | POD72 | peroxidase 72 | ROS scavenging | late response gene |
| LOC110692926 | POD5 | peroxidase 5 | ROS scavenging | late response gene |
| LOC110699378 | POD4 | peroxidase 4 | ROS scavenging | late response gene |
| LOC110724764 | POD9 | peroxidase 9 | ROS scavenging | late response gene |
| LOC110735668 | POD12 | peroxidase 12 | ROS scavenging | late response gene |
| LOC110694635 | POD55 | peroxidase 55 | ROS scavenging | late response gene |
| LOC110735670 | POD12 | peroxidase 12 | ROS scavenging | late response gene |
| LOC110681844 | POD55 | peroxidase 55 | ROS scavenging | late response gene |
| LOC110687369 | POD42 | peroxidase 42 | ROS scavenging | late response gene |
| LOC110690635 | POD1 | cationic peroxidase 1 | ROS scavenging | late response gene |
| LOC110727528 | POD5 | peroxidase 5 | ROS scavenging | late response gene |
| LOC110699380 | POD4 | peroxidase 4 | ROS scavenging | late response gene |
| LOC110684661 | POD1 | cationic peroxidase 1 | ROS scavenging | late response gene |
| LOC110704239 | POD12 | peroxidase 12 | ROS scavenging | post-transcriptional modification |
| LOC110710365 | HKT5 | potassium transporter 5 | osmotic adjustment | late response gene |
| LOC110689438 | PT1-3 | phosphate transporter 1-3 | osmotic adjustment | late response gene |
| LOC110720352 | PHO1 | phosphate transporter PHO1 | osmotic adjustment | late response gene |
| LOC110689401 | PHO1 | phosphate transporter PHO1 | osmotic adjustment | late response gene |
| LOC110717783 | PT1-3 | phosphate transporter 1-3 | osmotic adjustment | late response gene |
| LOC110727554 | SMT | sodium/metabolite cotransporter | osmotic adjustment | late response gene |
| LOC110688100 | NRT2.1 | high-affinity nitrate transporter 2.1 | osmotic adjustment | late response gene |
| LOC110684366 | NRT2.1 | high-affinity nitrate transporter 2.1 | osmotic adjustment | late response gene |
| LOC110715529 | NRT3.2 | high-affinity nitrate transporter 3.2 | osmotic adjustment | late response gene |
| LOC110684367 | NRT2.4 | high-affinity nitrate transporter 2.4 | osmotic adjustment | late response gene |
| LOC110699138 | CAH20 | cation/H+ antiporter 20 | osmotic adjustment | late response gene |
| LOC110709231 | NCL | sodium/calcium exchanger NCL | osmotic adjustment | late response gene |
| LOC110697673 | PIP2-5 | aquaporin PIP2-5 | osmotic adjustment | post-transcriptional modification |
| LOC110737811 | OCTN7 | organic cation/carnitine transporter 7 | osmotic adjustment | late response gene |
| LOC110688161 | ALMT2 | aluminum-activated malate transporter 2 | osmotic adjustment | late response gene |
| LOC110725786 | SWEET1 | bidirectional sugar transporter SWEET1 | osmotic adjustment | late response gene |
| LOC110735791 | SWEET1 | bidirectional sugar transporter SWEET1 | osmotic adjustment | late response gene |
| LOC110732264 | SWEET7 | bidirectional sugar transporter SWEET7 | osmotic adjustment | late response gene |
| LOC110722677 | POT | polyol transporter | osmotic adjustment | late response gene |
| LOC110733528 | AAT | amino acid transporter | osmotic adjustment | late response gene |
| LOC110708068 | NAT7 | nucleobase-ascorbate transporter 7 | osmotic adjustment | late response gene |
| LOC110712440 | ABCC15 | ABC transporter C family member 15 | osmotic adjustment | late response gene |
| LOC110707705 | ABCB8 | ABC transporter B family member 8 | osmotic adjustment | late response gene |
| LOC110729523 | ABCC10 | ABC transporter C family member 10 | osmotic adjustment | late response gene |
| LOC110695413 | ABCA2 | ABC transporter A family member 2 | osmotic adjustment | late response gene |
| LOC110721597 | ABCC15 | ABC transporter C family member 15 | osmotic adjustment | late response gene |
| LOC110722212 | ABCB25 | ABC transporter B family member 25 | osmotic adjustment | late response gene |
| LOC110717180 | GLU1 | glucan endonucleases-1 | osmotic adjustment | post-transcriptional modification |
| LOC110717177 | GLU1 | lucan endonucleases-1 | osmotic adjustment | early response gene |
| LOC110699037 | GLU1 | glucan endonucleases-1 | osmotic adjustment | late response gene |
| LOC110717159 | GLU1 | glucan endonucleases-1 | osmotic adjustment | late response gene |
| LOC110699174 | GLU1 | glucan endonucleases-1 | osmotic adjustment | late response gene |
| LOC110736258 | GLU1 | glucan endonucleases-1 | osmotic adjustment | late response gene |
| LOC110727927 | SS | sucrose synthase | osmotic adjustment | post-transcriptional modification |
| LOC110689796 | SS | sucrose synthase | osmotic adjustment | post-transcriptional modification |
| LOC110739769 | BGLU13 | beta-glucosidase 13 | cell wall construction | late response gene |
| LOC110724275 | BGLU12 | beta-glucosidase 12 | cell wall construction | late response gene |
| LOC110682558 | BGAL3 | beta-galactosidase 3 | cell wall construction | late response gene |
| LOC110685863 | BGAL3 | beta-galactosidase 3 | cell wall construction | late response gene |
| LOC110715976 | CESA | cellulose synthase A | cell wall construction | late response gene |
| LOC110717430 | CESG2 | cellulose synthase G2 | cell wall construction | late response gene |
| LOC110689768 | CESD5 | cellulose synthase D5 | cell wall construction | late response gene |
| LOC110689717 | CESA | cellulose synthase A | cell wall construction | late response gene |
| LOC110721870 | CESA | cellulose synthase A | cell wall construction | late response gene |
| LOC110715157 | TBL38 | trichome birefringence-like protein 38 | cell wall construction | late response gene |
| LOC110685228 | TBL39 | trichome birefringence-like protein 39 | cell wall construction | late response gene |
| LOC110732550 | GRP1.8 | glycine-rich cell wall structural protein 1.8 | cell wall construction | late response gene |
| LOC110730178 | GRP1.8 | glycine-rich cell wall structural protein 1.8 | cell wall construction | late response gene |
| LOC110730179 | GRP1.8 | glycine-rich cell wall structural protein 1.8 | cell wall construction | late response gene |
| LOC110732549 | GRP1.8 | glycine-rich cell wall structural protein 1.8 | cell wall construction | late response gene |
| LOC110714725 | CGT | crocetin glucosyltransferase | secondary metabolism | late response gene |
| LOC110729660 | OGT | O-glucosyltransferase | secondary metabolism | late response gene |
| LOC110706607 | 7DGT | 7-deoxyloganetin glucosyltransferase | secondary metabolism | late response gene |
| LOC110739778 | 7DGT | 7-deoxyloganetic acid glucosyltransferase | secondary metabolism | late response gene |
| LOC110683464 | OGT | O-glucosyltransferase | secondary metabolism | late response gene |
| LOC110722666 | GT | hydroquinone glucosyltransferase | secondary metabolism | late response gene |
| LOC110711362 | ABGT | anthocyanin 3′-O-beta-glucosyltransferase | secondary metabolism | late response gene |
| LOC110738265 | ABGT | anthocyanin 3′-O-beta-glucosyltransferase | secondary metabolism | late response gene |
| LOC110735480 | UDPGT | UDP-glycosyltransferase 90A1 | secondary metabolism | late response gene |
| LOC110718641 | 7DGT | 7-deoxyloganetin glucosyltransferase | secondary metabolism | post-transcriptional modification |
| LOC110691783 | GPAT | glycerol-3-phosphate acyltransferase | secondary metabolism | late response gene |
| LOC110722317 | GPAT7 | glycerol-3-phosphate acyltransferase 7 | secondary metabolism | late response gene |
| LOC110733316 | GPAT5 | glycerol-3-phosphate acyltransferase 5 | secondary metabolism | late response gene |
| LOC110714505 | WSD1 | O-acyltransferase WSD1 | secondary metabolism | late response gene |
| LOC110691992 | CHS3 | chalcone synthase 3 | secondary metabolism | late response gene |
| LOC110691988 | CHS3 | chalcone synthase 3 | secondary metabolism | late response gene |
| LOC110702060 | CHS3 | chalcone synthase 3 | secondary metabolism | late response gene |
| LOC110735138 | GELP | GDSL esterase/lipase | secondary metabolism | late response gene |
| LOC110712448 | GELP | GDSL esterase/lipase | secondary metabolism | late response gene |
| LOC110709557 | GELP | GDSL esterase/lipase | secondary metabolism | late response gene |
| LOC110717860 | GELP | GDSL esterase/lipase | secondary metabolism | late response gene |
| LOC110703315 | GELP | GDSL esterase/lipase | secondary metabolism | post-transcriptional modification |
| LOC110731693 | CYP76AD1 | cytochrome P450 76AD1 | secondary metabolism | late response gene |
| LOC110739776 | CYP72A219 | cytochrome P450 72A219 | secondary metabolism | late response gene |
| LOC110718248 | CYP71A6 | cytochrome P450 71A6 | secondary metabolism | late response gene |
| LOC110727125 | CYP71A6 | cytochrome P450 71A6 | secondary metabolism | late response gene |
| LOC110681912 | CYP89A2 | cytochrome P450 89A2 | secondary metabolism | late response gene |
| LOC110724693 | CYP83B1 | cytochrome P450 83B1 | secondary metabolism | late response gene |
| LOC110703261 | OMT | O-methyltransferase | secondary metabolism | post-transcriptional modification |
| LOC110728006 | NMT | N-methyltransferase | secondary metabolism | post-transcriptional modification |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Q.; Su, C.; Dong, C.-H. Genome-Wide Transcriptomic and Proteomic Exploration of Molecular Regulations in Quinoa Responses to Ethylene and Salt Stress. Plants 2021, 10, 2281. https://doi.org/10.3390/plants10112281
Ma Q, Su C, Dong C-H. Genome-Wide Transcriptomic and Proteomic Exploration of Molecular Regulations in Quinoa Responses to Ethylene and Salt Stress. Plants. 2021; 10(11):2281. https://doi.org/10.3390/plants10112281
Chicago/Turabian StyleMa, Qian, Chunxue Su, and Chun-Hai Dong. 2021. "Genome-Wide Transcriptomic and Proteomic Exploration of Molecular Regulations in Quinoa Responses to Ethylene and Salt Stress" Plants 10, no. 11: 2281. https://doi.org/10.3390/plants10112281
APA StyleMa, Q., Su, C., & Dong, C. -H. (2021). Genome-Wide Transcriptomic and Proteomic Exploration of Molecular Regulations in Quinoa Responses to Ethylene and Salt Stress. Plants, 10(11), 2281. https://doi.org/10.3390/plants10112281