
Transformation of Seed Non-Transmissible Hop Viroids in Nicotiana benthamiana Causes Distortions in Male Gametophyte Development

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Flower and Pollen Developmental Stages in Nicotiana benthamiana
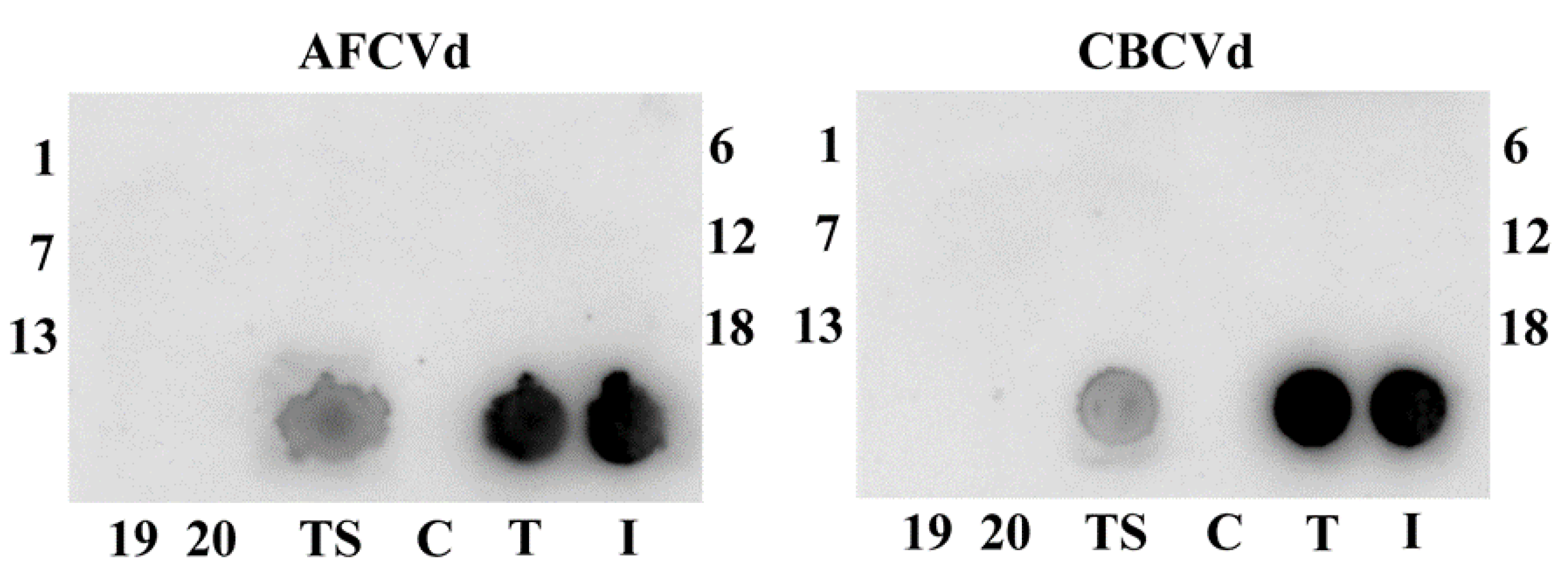
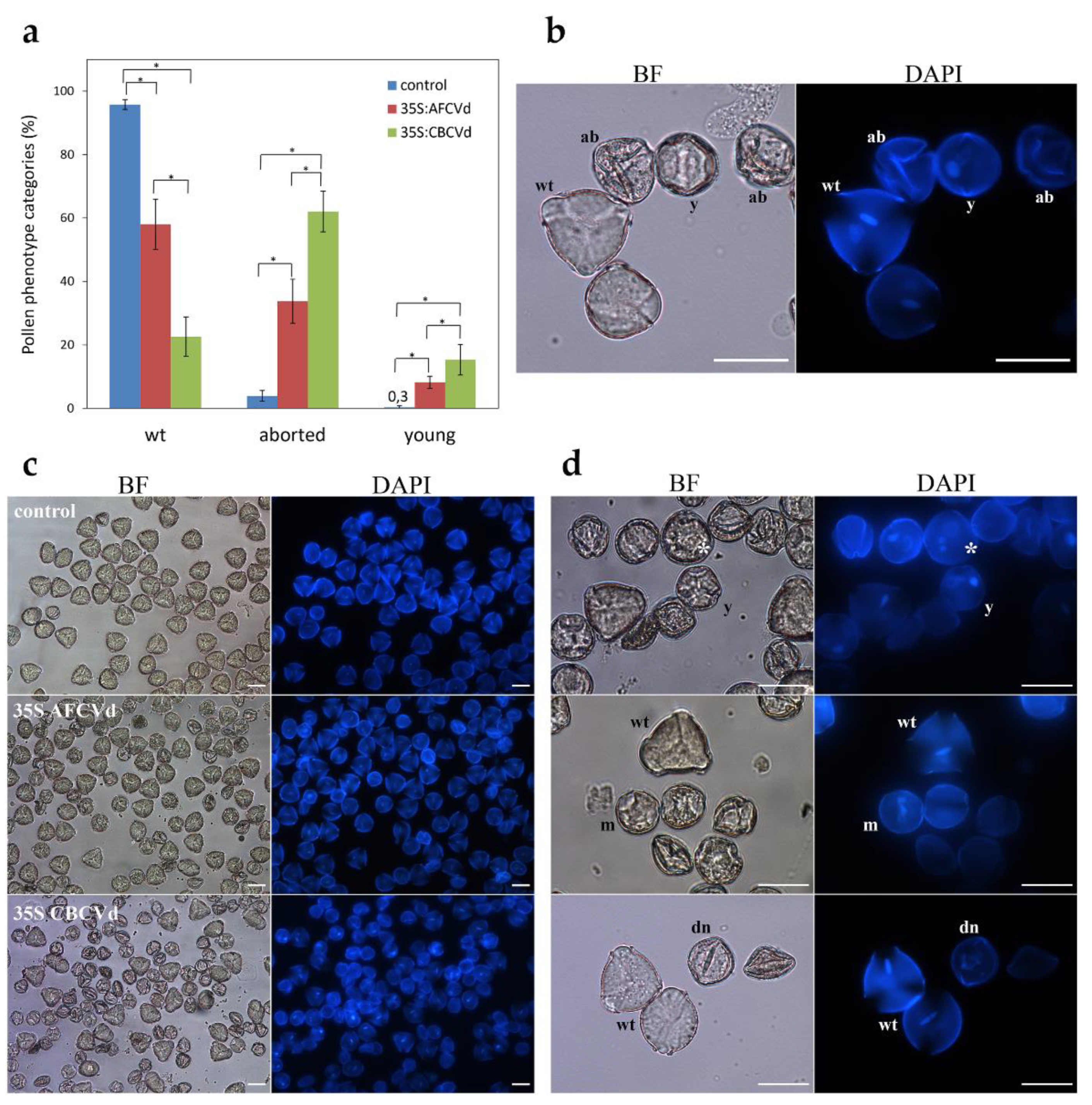
2.2. Viroid Elimination during Pollen Development
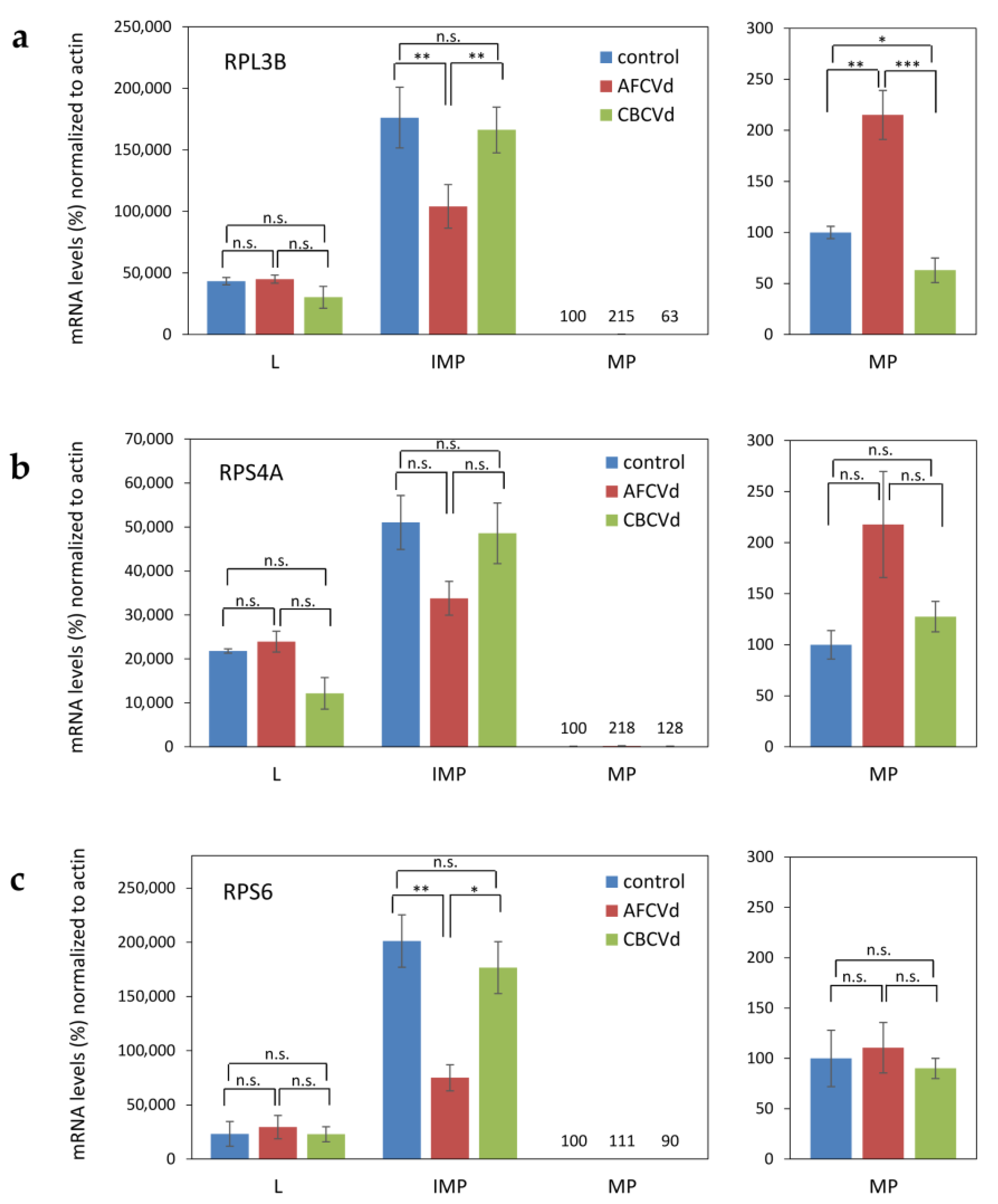
2.3. Viroid Pathogenesis Levels in Naturally Infected and Viroid cDNA Transformed Pollen
3. Materials and Methods
3.1. Plant Cultivation, Transformation and Viroid Inoculation
3.2. Viroid Detection and Quantification
3.3. mRNA Quantification of Ribosomal Protein mRNAs as Markers
3.4. Pollen Developmental Stages Characterization
3.5. Pollen Collection for Analysis
3.6. Mature Pollen Grain Phenotypes
3.7. Sequence Assembly and Structure Prediction of AFCVd, CBCVd and PSTVd
3.8. Statistical Evaluation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hafidh, S.; Honys, D. Reproduction Multitasking: The Male Gametophyte. Annu. Rev. Plant Biol. 2021, 72, 581–614. [Google Scholar] [CrossRef] [PubMed]
- Brewbaker, J.L. The distribution and phylogenetic significance of binucleate and trinucleate pollen grains in the Angiosperms. Amer. J. Bot. 1967, 54, 1069–1083. [Google Scholar] [CrossRef]
- McCormick, S. Male gametophyte development. Plant Cell 1993, 5, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Hafidh, S.; Potešil, D.; Fíla, J.; Čapková, V.; Zdráhal, Z.; Honys, D. Quantitative proteomics of the tobacco pollen tube secretome identifies novel pollen tube guidance proteins important for fertilization. Genome Biol. 2016, 17, 81. [Google Scholar] [CrossRef] [Green Version]
- Slotkin, R.; Vaughn, M.; Borges, F.; Tanurdzi, M.; Becker, J.; Feijó, J.; Martienssen, R. Epigenetic reprogramming and small RNA silencing of transposable elements in pollen. Cell 2009, 136, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Matoušek, J.; Orctová, L.; Škopek, J.; Pešina, K.; Steger, G. Elimination of hop latent viroid upon developmental activation of pollen nucleases. Biol. Chem. 2008, 389, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Matoušek, J.; Steinbachová, L.; Záveská Drábková, L.; Kocábek, T.; Potěšil, D.; Mishra, A.; Honys, D.; Steger, G. Elimination of viroids from tobacco pollen involves a decrease in propagation rate and an increase of the degradation processes. Int. J. Mol. Sci. 2020, 21, 3029. [Google Scholar] [CrossRef] [PubMed]
- Diener, T. Origin and evolution of viroids and viroid-like satellite RNAs. Virus Genes 1995, 11, 119–131. [Google Scholar] [CrossRef]
- Flores, R.; Hernández, C.; Martínez de Alba, A.E.; Daròs, J.A.; Di Serio, F. Viroids and viroid-host interactions. Annu. Rev. Phytopathol. 2005, 43, 117–139. [Google Scholar] [CrossRef]
- Venkataraman, S.; Badar, U.; Shoeb, E.; Hashim, G.; Abouhaidar, M.; Hefferon, K. An inside look into biological miniatures: Molecular mechanisms of viroids. Int. J. Mol. Sci. 2021, 22, 2795. [Google Scholar] [CrossRef] [PubMed]
- Sano, T. Compendium of Hop Disease and Pests; Mahaffee, W., Pethybridge, S., Gent, D.H., Eds.; American Phytopathological Society: St. Paul, MI, USA, 2009; pp. 39–132. [Google Scholar]
- Jakše, J.; Radišek, S.; Pokorn, T.; Matoušek, J.; Javornik, B. Deep-sequencing revealed Citrus bark cracking viroid (CBCVd) as a highly aggressive pathogen on hop. Plant Pathol. 2015, 64, 831–842. [Google Scholar] [CrossRef]
- Schindler, I.M.; Mühlbach, H.P. Involvement of nuclear DNA-dependent RNA polymerases in potato spindle tuber viroid replication: A reevaluation. Plant Sci. 1992, 84, 221–229. [Google Scholar] [CrossRef]
- Zhong, X.; Archual, A.J.; Amin, A.A.; Ding, B. A genomic map of viroid RNA motifs critical for replication and systemic trafficking. Plant Cell 2008, 20, 35–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, D.; Wang, M.; Li, S. Functional analysis of a viroid RNA motif mediating cell-to-cell movement in Nicotiana benthamiana. J. Gen. Virol. 2017, 98, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Wassenegger, M.; Spieker, R.L.; Thalmeir, S.; Riedel, L.; Sänger, H.L. A single nucleotide substitution converts Potato spindle tuber viroid (PSTVd) from a noninfectious to an infectious RNA for Nicotiana tabacum. Virology 1996, 226, 191–197. [Google Scholar] [CrossRef]
- Matsushita, Y.; Yanagisawa, H.; Sano, T. Vertical and horizontal transmission of Pospiviroids. Viruses 2018, 10, 706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desjardins, P.; Drake, R.; Atkins, E.; Bergh, B. Pollen transmission of avocado sunblotch virus experimentally demonstrated. Calif. Agric. 1979, 33, 14–15. [Google Scholar]
- Kryczynski, S.; Paduch-Cichal, E.; Skrzeczkowski, L. Transmission of three viroids through seed and pollen of tomato plants. J. Phytopathol. 1988, 121, 51–57. [Google Scholar] [CrossRef]
- Singh, R.; Boucher, A.; Somerville, T. Detection of potato spindle tuber viroid in the pollen and various parts of potato plant pollinated with viroid-infected pollen. Plant Dis. 1992, 76, 951–953. [Google Scholar] [CrossRef]
- Mink, G. Pollen and seed-transmitted viruses and viroids. Annu. Rev. Phytopathol. 1993, 31, 375–402. [Google Scholar] [CrossRef]
- Pacumbaba, E.; Zelazny, B.; Orense, J.; Rillo, E. Evidence for pollen and seed transmission of the coconut cadang-cadang viroid in Cocos nucifera. J. Phytopathol. 1994, 142, 37–42. [Google Scholar] [CrossRef]
- Yanagisawa, H.; Sano, T.; Hase, S.; Matsushita, Y. Influence of the terminal left domain on horizontal and vertical transmissions of tomato planta macho viroid and potato spindle tuber viroid through pollen. Virology 2018, 526, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Castellano, M.; Martinez, G.; Marques, M.C.; Moreno-Romero, J.; Köhler, C.; Pallas, V.; Gomez, G. Changes in the DNA methylation pattern of the host male gametophyte of viroid-infected cucumber plants. J. Exp. Bot. 2016, 67, 5857–5868. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.; Mishra, A.K.; Matoušek, J.; Steinbachová, L.; Potměšil, D.; Nath, V.S.; Awasthi, P.; Kocábek, T.; Jakše, J.; Záveská Drábková, L.; et al. Integrated Proteo-Transcriptomic Analyses Reveal Insights into Regulation of Pollen Development Stages and Dynamics of Cellular Response to Apple Fruit Crinkle Viroid (AFCVd)-Infection in Nicotiana tabacum. Int. J. Mol. Sci. 2020, 21, 8700. [Google Scholar] [CrossRef]
- Matoušek, J.; Siglová, K.; Jakše, J.; Radišek, S.; Tsushima, T.; Brass, J.R.; Guček, T.; Duraisamy, G.; Sano, T.; Steger, G. Propagation and some physiological effects of Citrus bark cracking viroid and Apple fruit crinkle viroid in multiple infected hop (Humulus lupulus L.). J. Plant Physiol. 2017, 213, 166–177. [Google Scholar] [CrossRef]
- Tupý, J.; Süss, J.; Hrabětová, E.; Říhová, L. Developmental changes in gene expression during pollen differentiation and maturation in Nicotiana tabacum L. Biol. Plant. 1983, 25, 231. [Google Scholar] [CrossRef]
- Honys, D.; Reňák, D.; Twell, D. Male gametophyte development and function. In Floriculture, Ornamental and Plant Biotechnology-Advances and Topical Issues; da Silva, J.A.T., Ed.; Global Science Books Ltd.: Ikenobe, Japan, 2006; Volume I, pp. 76–87. [Google Scholar]
- Matoušek, J.; Trněná, L.; Svoboda, P.; Oriniaková, P.; Lichtenstein, C.P. The gradual reduction of viroid levels in hop mericlones following heat therapy: A possible role for a nuclease degrading dsRNA. Biol. Chem. Hoppe-Seyler 1995, 376, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Matoušek, J.; Patzak, J. A low transmissibility of hop latent viroid through a generative phase of Humulus lupulus L. Biol. Plant. 2000, 43, 145–148. [Google Scholar] [CrossRef]
- Fernow, K.H.; Peterson, L.C.; Plaisted, R.L. Spindle tuber virus in seeds and pollen of infected potato plants. Am. Potato J. 1970, 47, 75–80. [Google Scholar] [CrossRef]
- Matsushita, Y.; Tsuda, S. Seed transmission of potato spindle tuber viroid, tomato chlorotic dwarf viroid, tomato apical stunt viroid, and Columnea latent viroid in horticultural plants. Eur. J. Plant Pathol. 2016, 145, 1007–1011. [Google Scholar] [CrossRef]
- Verhoeven, J.T.J.; Botermans, M.; Roenhorst, J.W.; Westerhof, J.; Meekes, E.T.M. First Report of Potato spindle tuber viroid in Cape Gooseberry (Physalis peruviana) from Turkey and Germany. Plant Dis. 2009, 93, 316. [Google Scholar] [CrossRef] [PubMed]
- Matoušek, J.; Kozlová, P.; Orctová, L.; Schmitz, A.; Pešina, K.; Bannach, O.; Diermann, D.; Steger, G.; Riesner, D. Accumulation of viroid-specific small RNAs and increase of nucleolytic activities linked to viroid-caused pathogenesis. Biol. Chem. 2007, 388, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Keese, P.; Symons, R.H. Domains in viroids: Evidence of intermolecular RNA rearrangement and their contribution to viroid evolution. Proc. Natl. Acad. Sci. USA 1985, 82, 4582–4586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, X.; Tao, X.; Stombaugh, J.; Leontis, N.; Ding, B. Tertiary structure and function of an RNA motif required for plant vascular entry to initiate systemic trafficking. EMBO J. 2007, 26, 3836–3846. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhou, C.; Li, J.; Li, C.; Tao, X.; Leontis, N.B.; Zirbel, C.; Bisaro, D.M.; Ding, B. Functional analysis reveals G/U pairs critical for replication and trafficking of an infectious non-coding viroid RNA. Nucl. Acids Res. 2020, 48, 3134–3155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafidh, S.; Potěšil, D.; Müller, K.; Fíla, J.; Michailidis, C.; Herrmannová, A.; Feciková, J.; Ischebeck, T.; Valášek, L.S.; Zdráhal, Z.; et al. Dynamics of the Pollen Sequestrome Defined by Subcellular Coupled Omics. Plant Physiol. 2018, 178, 258–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matoušek, J.; Piernikarczyk, R.; Týcová, A.; Duraisamy, G.; Kocábek, T.; Steger, G. Expression of SANT/HTH Myb mRNA, a plant morphogenesis-regulating transcription factor, changes due to viroid infection. J. Plant Physiol. 2015, 183, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Horsch, R.; Fry, J.; Hoffman, N.; Eichholtz, D.; Rogers, S.; Fraley, R. A simple and general method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar] [CrossRef] [PubMed]
- Matoušek, J.; Schröder, A.R.W.; Trněná, L.; Reimers, M.; Baumstark, T.; Dědič, P.; Vlasák, J.; Becker, I.; Kreuzaler, F.; Fladung, M.; et al. Inhibition of viroid infection by antisense RNA expression in transgenic plants. Biol. Chem. Hoppe-Seyler 1994, 375, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Palukaitis, P.; Cotts, S.; Zaitlin, M. Detection and identification of viroids and viral nucleic acids by “dot-blot” hybridization. Acta Hortic. 1985, 164, 109–118. [Google Scholar] [CrossRef]
- Pfaffl, M. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Matoušek, J.; Junker, V.; Vrba, L.; Schubert, J.; Patzak, J.; Steger, G. Molecular characterization and genome organization of 7 SL RNA genes from hop (Humulus lupulus L.). Gene 1999, 239, 173–183. [Google Scholar] [CrossRef]
- Viral Genome Browser (Pospiviroidae). Available online: https://www.ncbi.nlm.nih.gov/genomes/GenomesGroup.cgi?taxid=185751 (accessed on 26 May 2021).
- Brister, J.R.; Ako-Adjei, D.; Bao, Y.; Blinkova, O. NCBI viral genomes resource. Nucleic Acids Res. 2015, 43, D571–D577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgulis, A.; Coulouris, G.; Raytselis, Y.; Madden, T.; Agarwala, R.; Schäffer, A. Database indexing for production MegaBLAST searches. Bioinformatics 2008, 24, 1757–1764. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Toh, H. Improved accuracy of multiple ncRNA alignment by incorporating structural information into a MAFFT-based framework. BMC Bioinform. 2008, 9, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilm, A.; Linnenbrink, K.; Steger, G. ConStruct: Improved construction of RNA consensus structures. BMC Bioinform. 2008, 9, 219. [Google Scholar] [CrossRef] [Green Version]
- Gruber, A.; Lorenz, R.; Bernhart, S.; Neuböck, R.; Hofacker, I. The Vienna RNA websuite. Nucleic Acids Res. 2008, 36, W70–W74. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, Z.; Breaker, R. R2R–software to speed the depiction of aesthetic consensus RNA secondary structures. BMC Bioinform. 2011, 12, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matoušek, J.; Kocábek, T.; Patzak, J.; Bříza, J.; Siglová, K.; Mishra, A.; Duraisamy, G.; Týcová, A.; Ono, E.; Krofta, K. The “putative” role of transcription factors from HlWRKY family in the regulation of the final steps of prenylflavonid and bitter acids biosynthesis in hop (Humulus lupulus L.). Plant Mol. Biol. 2016, 92, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Matoušek, J.; Stehlík, J.; Procházková, J.; Orctová, L.; Wullenweber, J.; Füssy, Z.; Kováčik, J.; Duraisamy, G.S.; Ziegler, A.; Schubert, J.; et al. Biological and molecular analysis of the pathogenic variant C3 of potato spindle tuber viroid (PSTVd) evolved during adaptation to chamomile (Matricaria chamomilla). Biol. Chem. 2012, 393, 605–615. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steinbachová, L.; Matoušek, J.; Steger, G.; Matoušková, H.; Radišek, S.; Honys, D. Transformation of Seed Non-Transmissible Hop Viroids in Nicotiana benthamiana Causes Distortions in Male Gametophyte Development. Plants 2021, 10, 2398. https://doi.org/10.3390/plants10112398
Steinbachová L, Matoušek J, Steger G, Matoušková H, Radišek S, Honys D. Transformation of Seed Non-Transmissible Hop Viroids in Nicotiana benthamiana Causes Distortions in Male Gametophyte Development. Plants. 2021; 10(11):2398. https://doi.org/10.3390/plants10112398
Chicago/Turabian StyleSteinbachová, Lenka, Jaroslav Matoušek, Gerhard Steger, Helena Matoušková, Sebastjan Radišek, and David Honys. 2021. "Transformation of Seed Non-Transmissible Hop Viroids in Nicotiana benthamiana Causes Distortions in Male Gametophyte Development" Plants 10, no. 11: 2398. https://doi.org/10.3390/plants10112398
APA StyleSteinbachová, L., Matoušek, J., Steger, G., Matoušková, H., Radišek, S., & Honys, D. (2021). Transformation of Seed Non-Transmissible Hop Viroids in Nicotiana benthamiana Causes Distortions in Male Gametophyte Development. Plants, 10(11), 2398. https://doi.org/10.3390/plants10112398