
Machine Learning-Mediated Development and Optimization of Disinfection Protocol and Scarification Method for Improved In Vitro Germination of Cannabis Seeds



Abstract
:1. Introduction
2. Results
2.1. Effect of Different Disinfectants at Various Immersion Times on Contamination
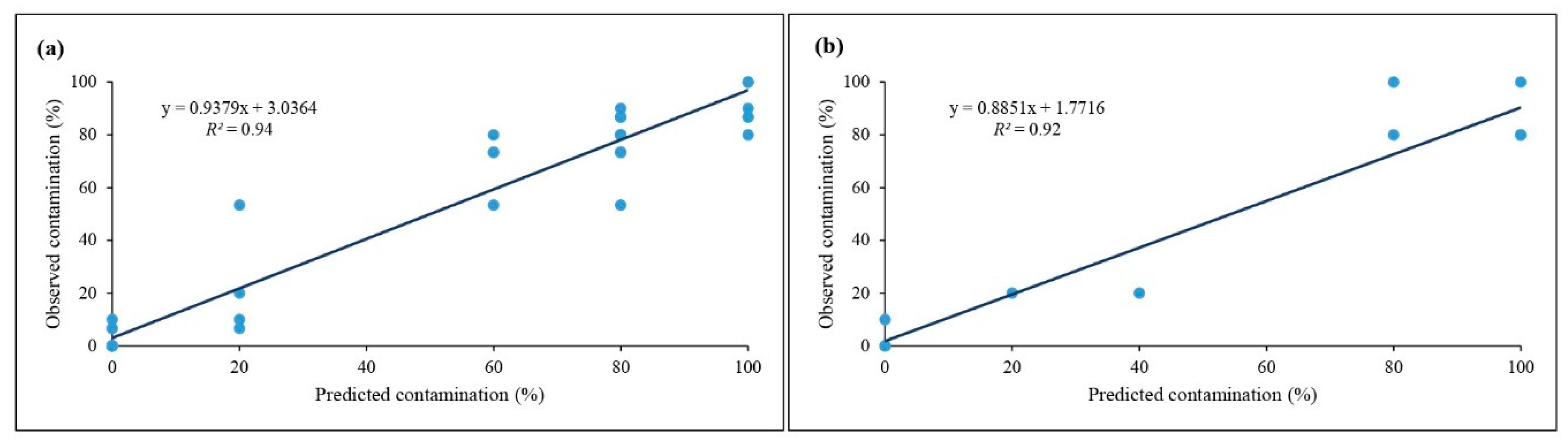
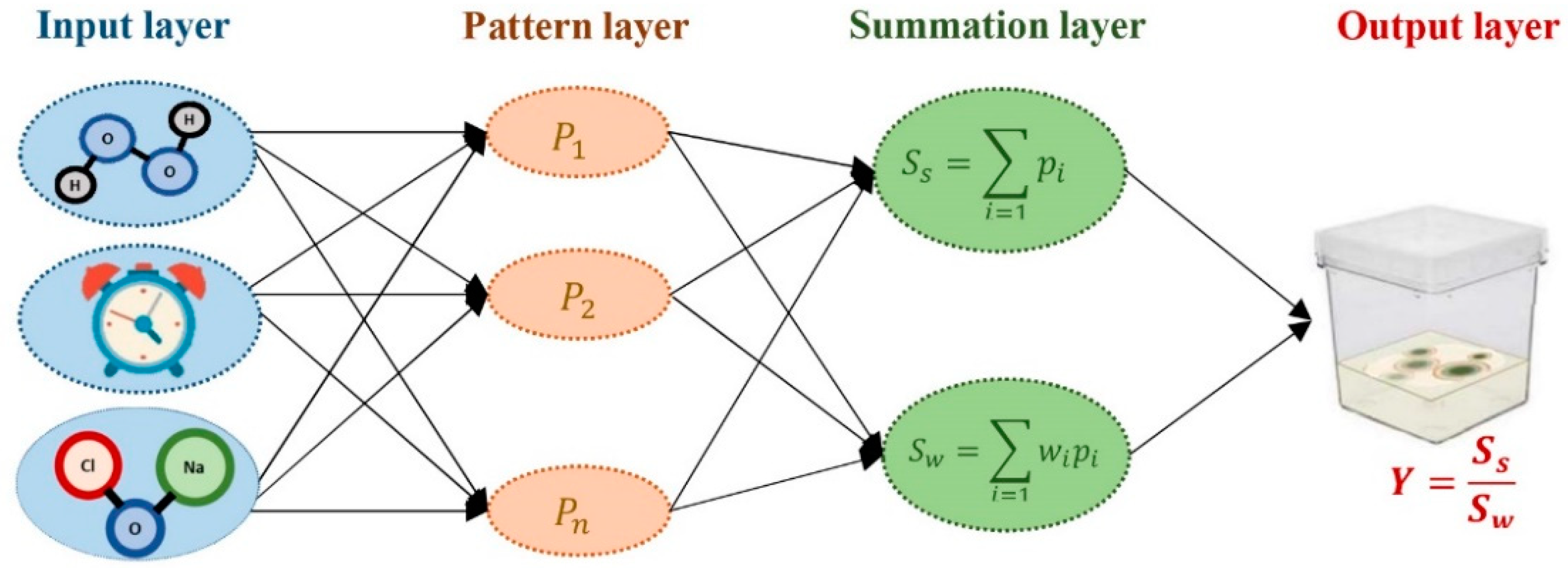
2.2. Data Modeling by Using GRNN
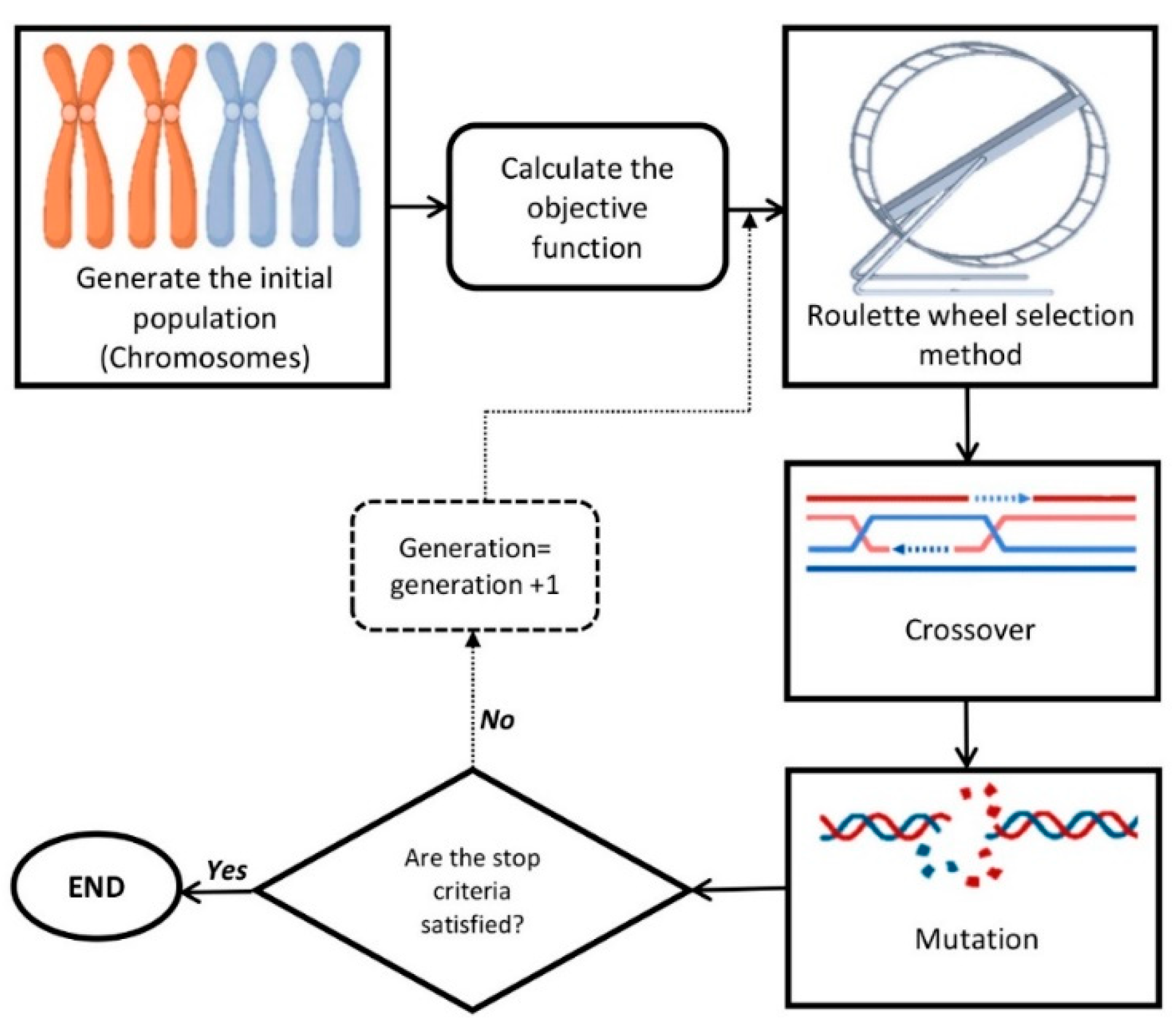
2.3. Optimization via GA and Validation Experiment
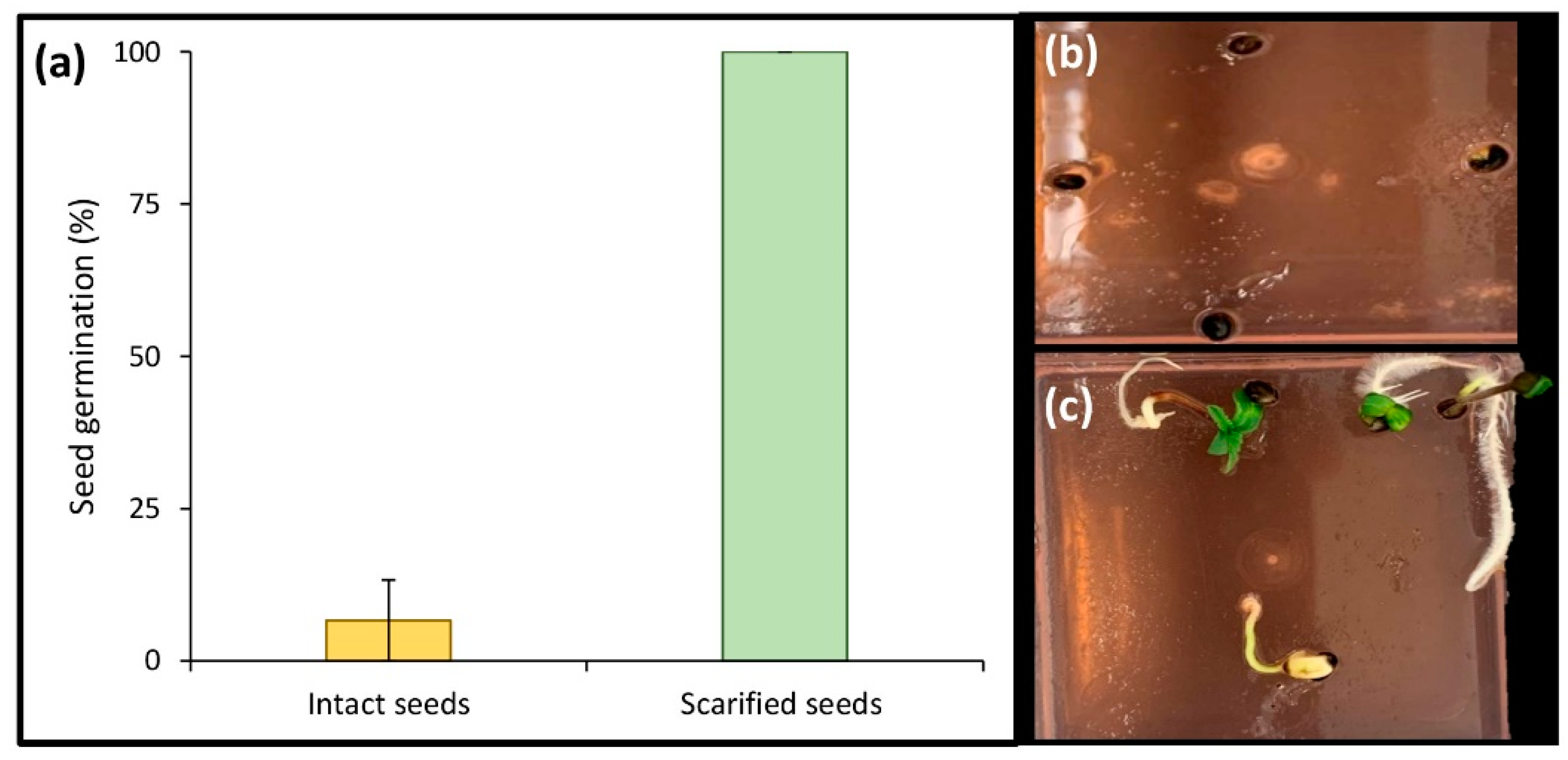
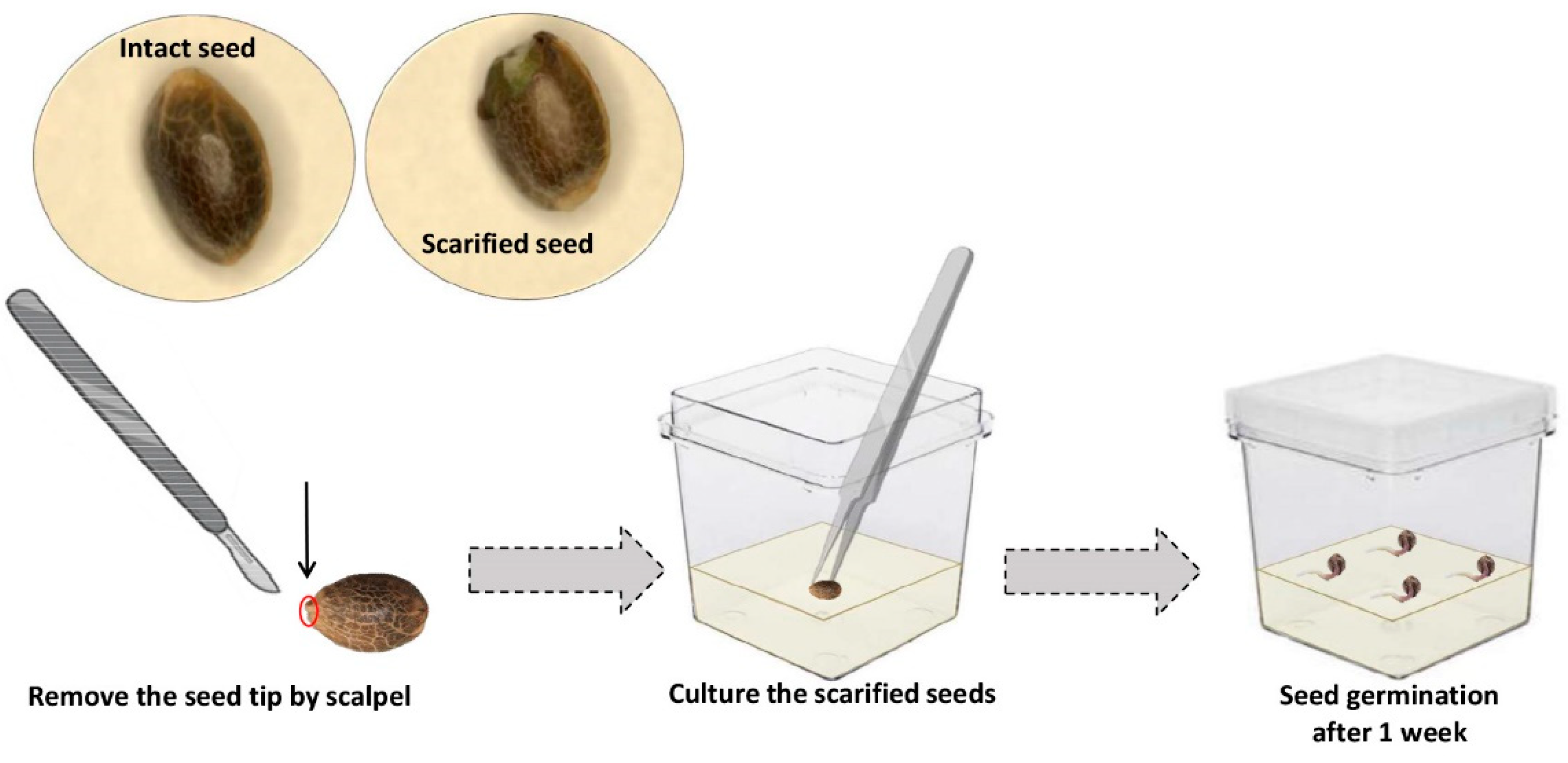
2.4. Effect of Scarification on In Vitro Seed Germination
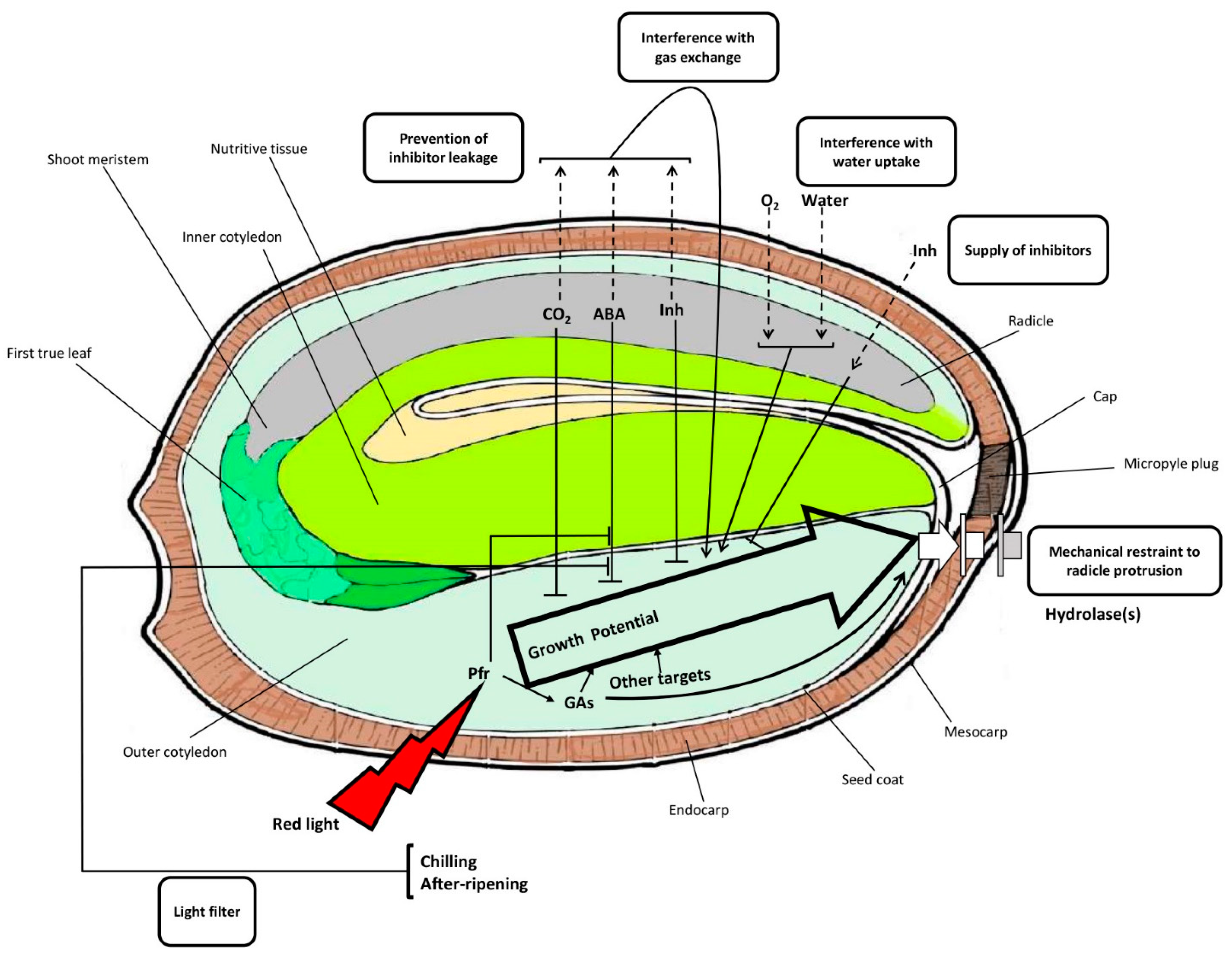
3. Discussion
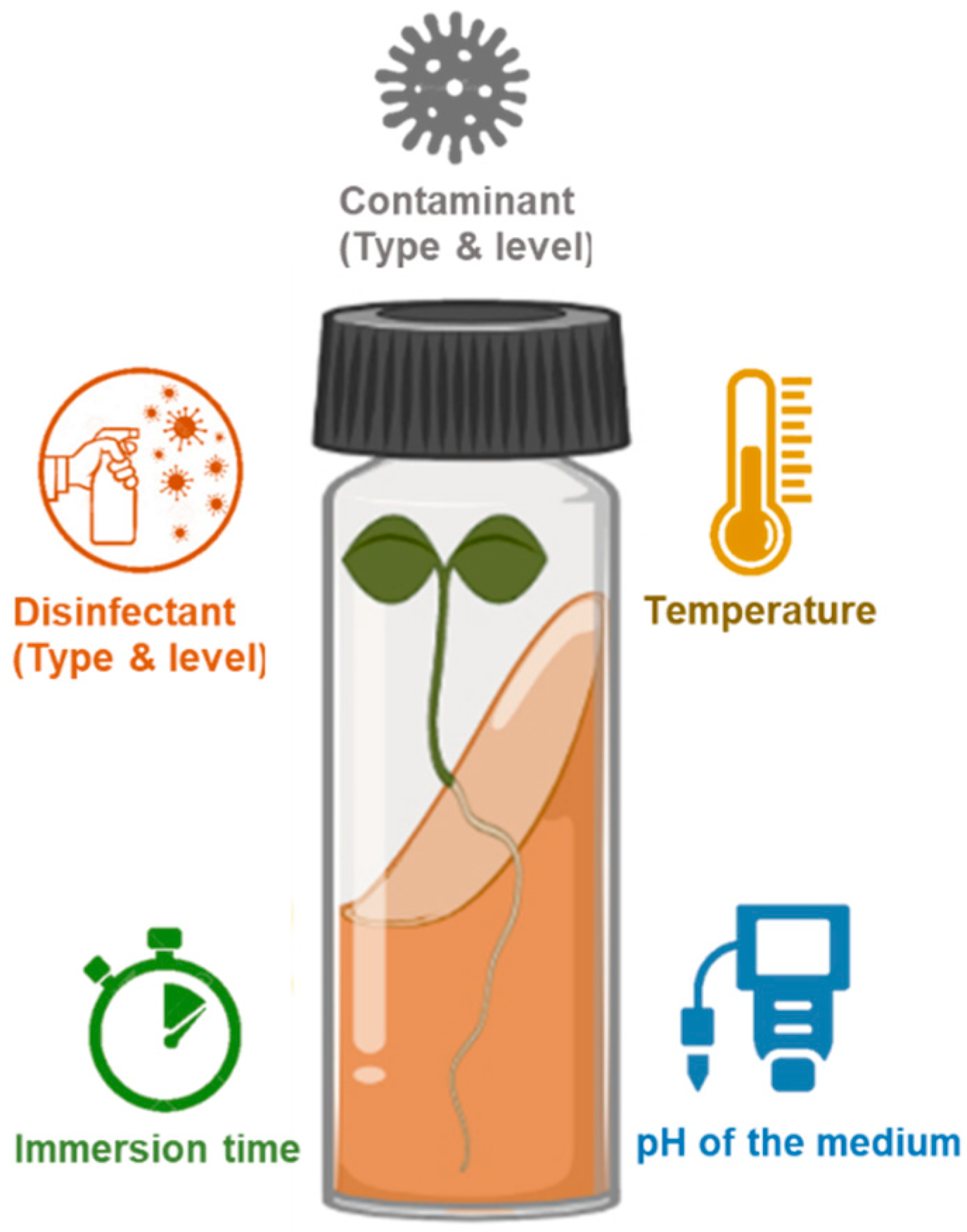
4. Materials and Methods
4.1. Sterilization Procedure
4.2. Modeling Procedure
4.3. Optimization Procedure and Validation Experiment
4.4. Scarification Procedure
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hesami, M.; Pepe, M.; Alizadeh, M.; Rakei, A.; Baiton, A.; Phineas Jones, A.M. Recent advances in cannabis biotechnology. Ind. Crop. Prod. 2020, 158, 113026. [Google Scholar] [CrossRef]
- Torkamaneh, D.; Jones, A.M.P. Cannabis, the multibillion dollar plant that no genebank wanted. Genome 2021, 64, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Small, E. Evolution and Classification of Cannabis sativa (Marijuana, Hemp) in Relation to Human Utilization. Bot. Rev. 2015, 81, 189–294. [Google Scholar] [CrossRef]
- Adhikary, D.; Kulkarni, M.; El-Mezawy, A.; Mobini, S.; Elhiti, M.; Gjuric, R.; Ray, A.; Polowick, P.; Slaski, J.J.; Jones, M.P.; et al. Medical cannabis and industrial hemp tissue culture: Present status and future potential. Front. Plant Sci. 2021, 12, 627240. [Google Scholar] [CrossRef] [PubMed]
- Hesami, M.; Pepe, M.; Monthony, A.S.; Baiton, A.; Phineas Jones, A.M. Modeling and optimizing in vitro seed germination of industrial hemp (Cannabis sativa L.). Ind. Crop. Prod. 2021, 170, 113753. [Google Scholar] [CrossRef]
- Page, S.R.G.; Monthony, A.S.; Jones, A.M.P. DKW basal salts improve micropropagation and callogenesis compared with MS basal salts in multiple commercial cultivars of Cannabis sativa. Botany 2021, 99, 269–279. [Google Scholar] [CrossRef]
- Hesami, M.; Jones, A.M.P. Application of artificial intelligence models and optimization algorithms in plant cell and tissue culture. Appl. Microbiol. Biotechnol. 2020, 104, 9449–9485. [Google Scholar] [CrossRef] [PubMed]
- Cuba-Díaz, M.; Rivera-Mora, C.; Navarrete, E.; Klagges, M. Advances of native and non-native Antarctic species to in vitro conservation: Improvement of disinfection protocols. Sci. Rep. 2020, 10, 3845. [Google Scholar] [CrossRef]
- Barampuram, S.; Allen, G.; Krasnyanski, S. Effect of various sterilization procedures on the in vitro germination of cotton seeds. Plant Cell Tissue Organ Cult. 2014, 118, 179–185. [Google Scholar] [CrossRef]
- Hesami, M.; Naderi, R.; Tohidfar, M. Modeling and Optimizing in vitro Sterilization of Chrysanthemum via Multilayer Perceptron-Non-dominated Sorting Genetic Algorithm-II (MLP-NSGAII). Front. Plant Sci. 2019, 10, 282. [Google Scholar] [CrossRef] [Green Version]
- Yoosefzadeh-Najafabadi, M.; Tulpan, D.; Eskandari, M. Application of machine learning and genetic optimization algorithms for modeling and optimizing soybean yield using its component traits. PLoS ONE 2021, 16, e0250665. [Google Scholar] [CrossRef] [PubMed]
- Hameg, R.; Arteta, T.A.; Landin, M.; Gallego, P.P.; Barreal, M.E. Modeling and optimizing culture medium mineral composition for in vitro propagation of Actinidia arguta. Front. Plant Sci. 2020, 11, 554905. [Google Scholar] [CrossRef]
- Niazian, M.; Niedbała, G. Machine learning for plant breeding and biotechnology. Agriculture 2020, 10, 436. [Google Scholar] [CrossRef]
- Yoosefzadeh-Najafabadi, M.; Tulpan, D.; Eskandari, M. Using hybrid artificial intelligence and evolutionary optimization algorithms for estimating soybean yield and fresh biomass using hyperspectral vegetation indices. Remote Sens. 2021, 13, 2555. [Google Scholar] [CrossRef]
- Zielińska, S.; Kępczyńska, E. Neural modeling of plant tissue cultures: A review. BioTechnologia 2013, 94, 253–268. [Google Scholar] [CrossRef]
- Hesami, M.; Jones, A.M.P. Modeling and optimizing callus growth and development in Cannabis sativa using random forest and support vector machine in combination with a genetic algorithm. Appl. Microbiol. Biotechnol. 2021, 105, 5201–5212. [Google Scholar] [CrossRef]
- Ivashchuk, O.A.; Fedorova, V.I.; Shcherbinina, N.V.; Maslova, E.V.; Shamraeva, E. Microclonal propagation of plant process modeling and optimization of its parameters based on neural network. Drug Invent. Today 2018, 10, 3170–3175. [Google Scholar]
- Pepe, M.; Hesami, M.; Small, F.; Jones, A.M.P. Comparative analysis of machine learning and evolutionary optimization algorithms for precision micropropagation of Cannabis sativa: Prediction and validation of in vitro shoot growth and development based on the optimization of light and carbohydrate sources. Front. Plant Sci. 2021, 12, 757869. [Google Scholar] [CrossRef]
- Salehi, M.; Farhadi, S.; Moieni, A.; Safaie, N.; Hesami, M. A hybrid model based on general regression neural network and fruit fly optimization algorithm for forecasting and optimizing paclitaxel biosynthesis in Corylus avellana cell culture. Plant Methods 2021, 17, 13. [Google Scholar] [CrossRef] [PubMed]
- Hesami, M.; Condori-Apfata, J.A.; Valderrama Valencia, M.; Mohammadi, M. Application of artificial neural network for modeling and studying in vitro genotype-independent shoot regeneration in wheat. Appl. Sci. 2020, 10, 5370. [Google Scholar] [CrossRef]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular aspects of seed dormancy. Annu. Rev. Plant Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef] [Green Version]
- Smýkal, P.; Vernoud, V.; Blair, M.W.; Soukup, A.; Thompson, R.D. The role of the testa during development and in establishment of dormancy of the legume seed. Front. Plant Sci. 2014, 5, 351. [Google Scholar] [CrossRef] [Green Version]
- Tognacca, R.S.; Botto, J.F. Post-transcriptional regulation of seed dormancy and germination: Current understanding and future directions. Plant Commun. 2021, 2, 100169. [Google Scholar] [CrossRef] [PubMed]
- Magrini, S.; De Vitis, M. In vitro reproduction of three Limodorum species (Orchidaceae): Impacts of scarification methods and nitrogen sources on mature seed germination and seedling development. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2017, 151, 419–428. [Google Scholar] [CrossRef]
- Musarurwa, H.T.; van Staden, J.; Makunga, N.P. In vitro seed germination and cultivation of the aromatic medicinal Salvia stenophylla (Burch. ex Benth.) provides an alternative source of α-bisabolol. Plant Growth Regul. 2010, 61, 287–295. [Google Scholar] [CrossRef]
- Carrera-Castaño, G.; Calleja-Cabrera, J.; Pernas, M.; Gómez, L.; Oñate-Sánchez, L. An updated overview on the regulation of seed germination. Plants 2020, 9, 703. [Google Scholar] [CrossRef]
- Han, C.; Yang, P. Studies on the molecular mechanisms of seed germination. Proteomics 2015, 15, 1671–1679. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, M.; Hoy, R.; Lu, B.; Song, L. Team effort: Combinatorial control of seed maturation by transcription factors. Curr. Opin. Plant Biol. 2021, 63, 102091. [Google Scholar] [CrossRef]
- Wu, Y.; Shen, Y.B. Seed coat structural and permeability properties of Tilia miqueliana seeds. J. Plant Growth Regul. 2021, 40, 1198–1209. [Google Scholar] [CrossRef]
- Saffari, P.; Majd, A.; Jonoubi, P.; Najafi, F. Effect of treatments on seed dormancy breaking, seedling growth, and seedling antioxidant potential of Agrimonia eupatoria L. J. Appl. Res. Med. Aromat. Plants 2021, 20, 100282. [Google Scholar] [CrossRef]
- Pérez-Jiménez, M.; Guevara-Gázquez, A.; Carrillo-Navarro, A.; Cos-Terrer, J. How carbon source and seedcoat influence the in vitro culture of peach (Prunus persica l. Batsch) immature seeds. HortScience 2021, 56, 136–137. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sodium Hypochlorite (%) | H2O2 (%) | Time (min) | Contamination (%) ± SE |
|---|---|---|---|
| 0 | 0 | 0 | 100.0 ± 0.00 |
| 0 | 10 | 5 | 86.7 ± 6.67 |
| 0 | 10 | 10 | 86.7 ± 6.67 |
| 0 | 10 | 20 | 86.7 ± 6.67 |
| 0 | 20 | 5 | 86.7 ± 6.67 |
| 0 | 20 | 10 | 86.7 ± 6.67 |
| 0 | 20 | 20 | 73.3 ± 6.67 |
| 0 | 30 | 5 | 86.7 ± 6.67 |
| 0 | 30 | 10 | 80.0 ± 11.55 |
| 0 | 30 | 20 | 73.3 ± 6.67 |
| 5 | 0 | 5 | 53.3 ± 17.64 |
| 5 | 0 | 10 | 26.7 ± 6.67 |
| 5 | 0 | 15 | 0.0 ± 0.00 |
| 10 | 0 | 5 | 6.7 ± 6.67 |
| 10 | 0 | 10 | 0.0 ± 0.00 |
| 10 | 0 | 15 | 0.0 ± 0.00 |
| 15 | 0 | 5 | 6.7 ± 6.67 |
| 15 | 0 | 10 | 0.0 ± 0.00 |
| 15 | 0 | 15 | 0.0 ± 0.00 |
| Criteria | Training Set | Testing Set |
|---|---|---|
| Histogram |  |  |
| R2 | 0.938 | 0.918 |
| RMSE | 9.888 | 12.247 |
| MBE | −2.250 | −5.000 |
| Optimal Level of Input Variables | Predicted Contamination (%) | Contamination (%) in Validation Experiment |
|---|---|---|
| 4.6% sodium hypochlorite + 0.008% hydrogen peroxide for 16.81 min | 0 | 0 ± 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pepe, M.; Hesami, M.; Jones, A.M.P. Machine Learning-Mediated Development and Optimization of Disinfection Protocol and Scarification Method for Improved In Vitro Germination of Cannabis Seeds. Plants 2021, 10, 2397. https://doi.org/10.3390/plants10112397
Pepe M, Hesami M, Jones AMP. Machine Learning-Mediated Development and Optimization of Disinfection Protocol and Scarification Method for Improved In Vitro Germination of Cannabis Seeds. Plants. 2021; 10(11):2397. https://doi.org/10.3390/plants10112397
Chicago/Turabian StylePepe, Marco, Mohsen Hesami, and Andrew Maxwell Phineas Jones. 2021. "Machine Learning-Mediated Development and Optimization of Disinfection Protocol and Scarification Method for Improved In Vitro Germination of Cannabis Seeds" Plants 10, no. 11: 2397. https://doi.org/10.3390/plants10112397
APA StylePepe, M., Hesami, M., & Jones, A. M. P. (2021). Machine Learning-Mediated Development and Optimization of Disinfection Protocol and Scarification Method for Improved In Vitro Germination of Cannabis Seeds. Plants, 10(11), 2397. https://doi.org/10.3390/plants10112397