
Examining the Role of Buzzing Time and Acoustics on Pollen Extraction of Solanum elaeagnifolium



Abstract
:1. Introduction
2. Results
2.1. Major Pollinators of S. elaeagnifolium in Its Native Range in LRGV, Texas
2.2. Bee Visit Time
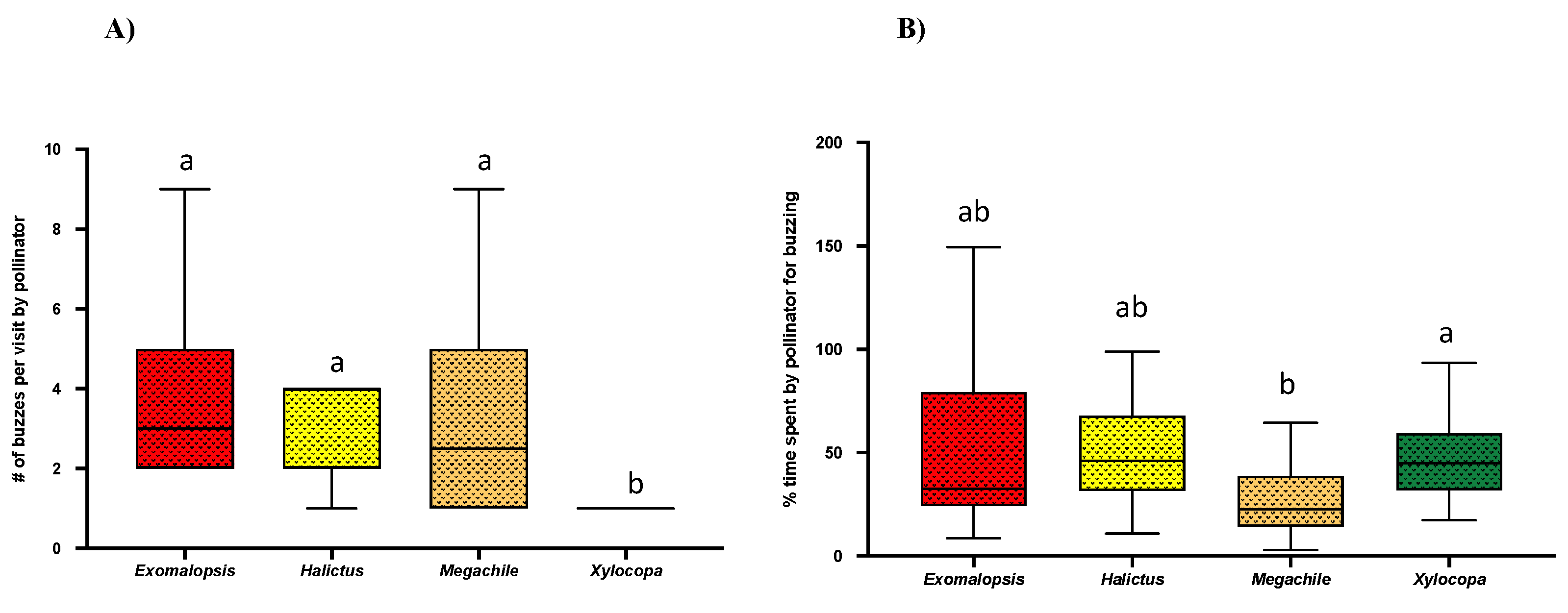
2.3. No. of Pulses Per Bee Visit and Buzz % over Visit Time
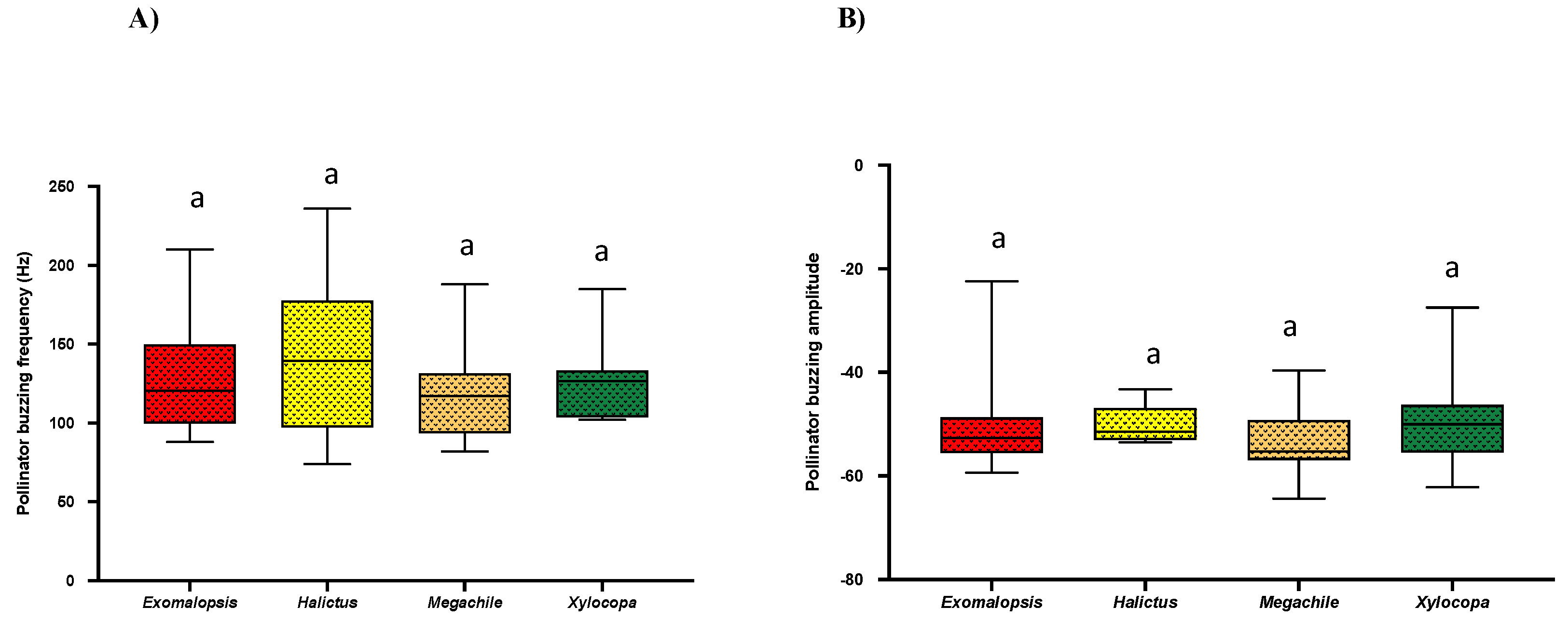
2.4. Bee Buzzing Frequency and Buzzing Amplitude
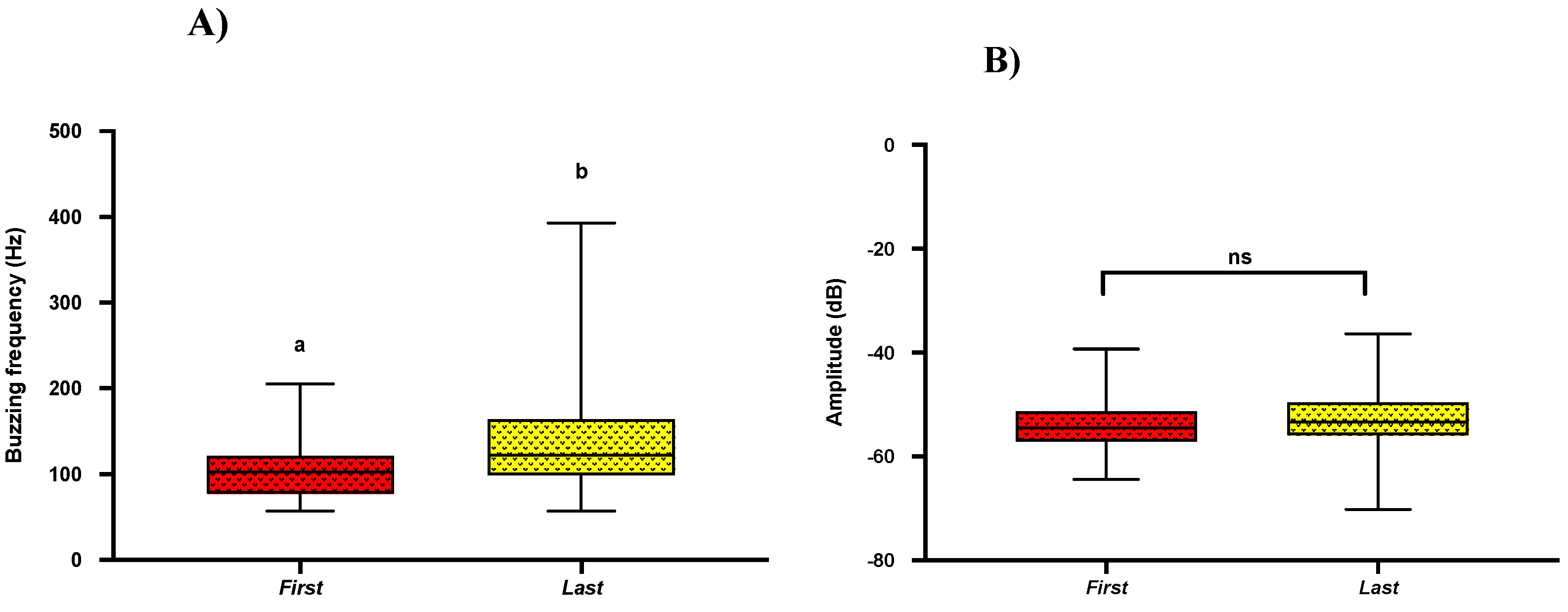
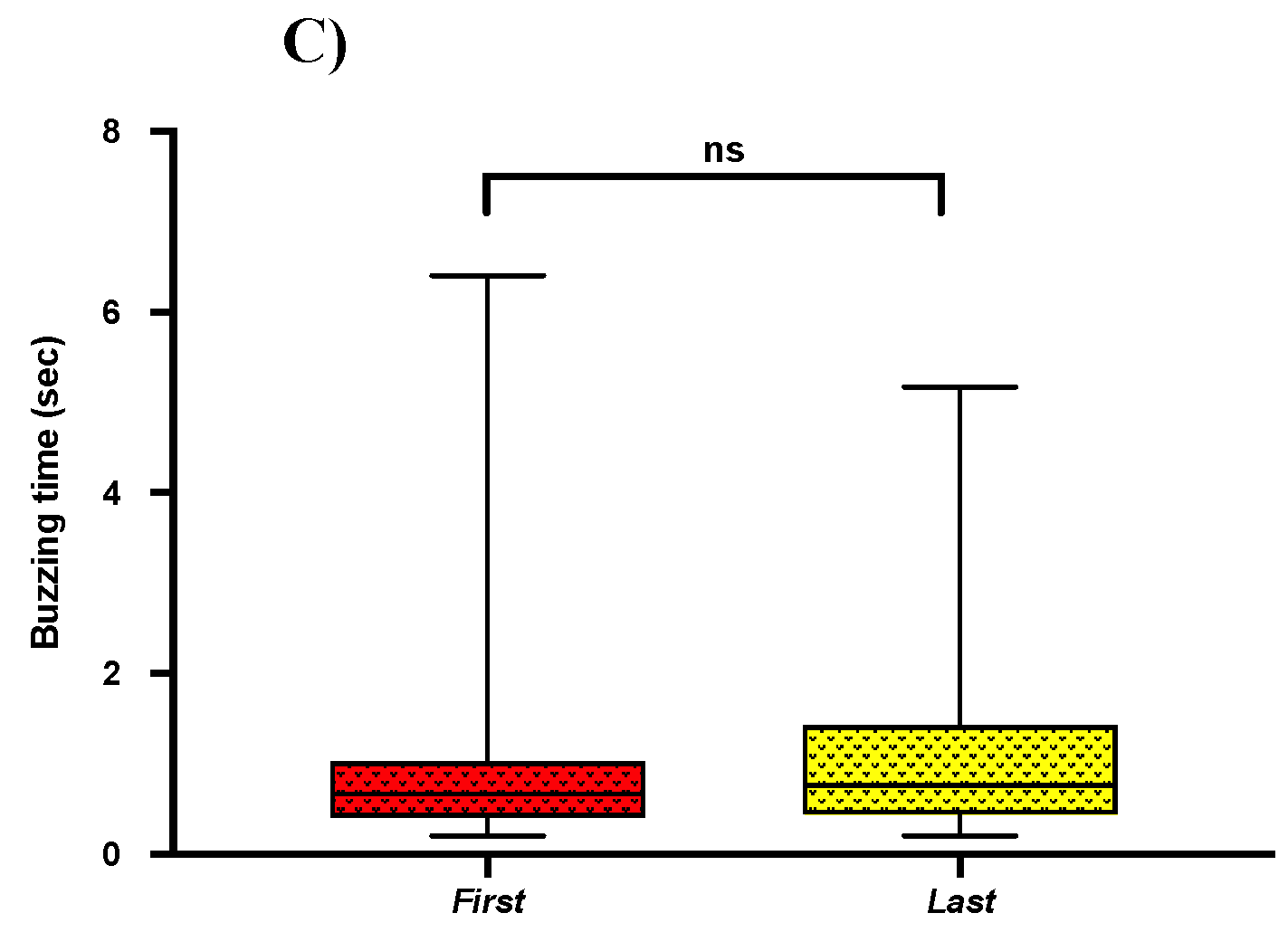
2.5. Acoustics of First and Last Buzz
2.6. Comparison of Bee Size among Different Genera
2.7. Artificial Pollen Extraction and Buzzing Time Intervals
3. Discussion
4. Materials and Methods
4.1. Study System
4.2. Field Survey and Bee Incidence
4.3. Bee Vibrations Recording and Bee Capturing
4.4. Estimating Vibration Frequency and Amplitude
4.5. Bee Mass and Intertegular Distance (ITD)
4.6. Artificial Pollen Extraction
Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buchmann, S.L. Buzz pollination in angiosperms. In Handbook of Experimental Pollination Biology; Scientific and Academic Editions; Jones, C.E., Little, J.R., Eds.; Van Nostrand Reinhold: New York, NY, USA, 1983; pp. 73–113. [Google Scholar]
- King, M.J.; Buchmann, S.L. Sonication Dispensing of Pollen from Solanum laciniatum Flowers. Funct. Ecol. 1996, 10, 449–456. [Google Scholar] [CrossRef]
- Arroyo-Correa, B.; Beattie, C.; Vallejo-Marín, M. Bee and floral traits affect the characteristics of the vibrations experienced by flowers during buzz-pollination. J. Exp. Biol. 2019, 222, jeb198176. [Google Scholar] [CrossRef] [Green Version]
- Cardinal, S.; Buchmann, S.L.; Russell, A.L. The evolution of floral sonication, a pollen foraging behavior used by bees (Anthophila). Evolution 2018, 72, 590–600. [Google Scholar] [CrossRef] [Green Version]
- Harder, L.D.; Barclay, R.M.R. The Functional Significance of Poricidal Anthers and Buzz Pollination: Controlled Pollen Removal from Dodecatheon. Funct. Ecol. 1994, 8, 509–517. [Google Scholar] [CrossRef]
- De Luca, P.A.; Vallejo-Marín, M. What’s the “buzz” about? The ecology and evolutionary significance of buzz-pollination. Curr. Opin. Plant Biol. 2013, 16, 429–435. [Google Scholar] [CrossRef]
- Kariyat, R.R.; Bentley, T.G.; Nihranz, C.T.; Stephenson, A.G.; De Moraes, C.M.; Mescher, M.C. Inbreeding in Solanum carolinense alters floral attractants and rewards and adversely affects pollinator visitation. Am. J. Bot. 2021, 108, 74–82. [Google Scholar] [CrossRef]
- Vogel, S. Evolutionary shifts from reward to deception in pollen flowers. In The Pollination of Flowers by Insects; Richards, A.J., Ed.; Academic Press: London, UK, 1978; pp. 89–96. [Google Scholar]
- Mast, A.R.; Feller, D.M.S.; Kelso, S.; Conti, E. Buzz-pollinated Dodecatheon originated from within the heterostylous Primula subgenus Auriculastrum (Primulaceae): A seven-region cpDNA phylogeny and its implications for floral evolution. Am. J. Bot. 2004, 91, 926–942. [Google Scholar] [CrossRef] [PubMed]
- Barrett, S.C.H. Darwin’s legacy: The forms, function and sexual diversity of flowers. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 351–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solís-Montero, L.; Vallejo-Marín, M. Does the morphological fit between flowers and pollinators affect pollen deposition? An experimental test in a buzz-pollinated species with anther dimorphism. Ecol. Evol. 2017, 7, 2706–2715. [Google Scholar] [CrossRef] [PubMed]
- Vallejo-Marín, M. Buzz pollination: Studying bee vibrations on flowers. New Phytol. 2019, 224, 1068–1074. [Google Scholar] [CrossRef] [Green Version]
- De Luca, P.A.; Buchmann, S.; Galen, C.; Mason, A.C.; Vallejo-Marín, M. Does body size predict the buzz-pollination frequencies used by bees? Ecol. Evol. 2019, 9, 4875–4887. [Google Scholar] [CrossRef]
- Tayal, M.; Chavana, J.; Kariyat, R.R. Efficiency of using electric toothbrush as an alternative to a tuning fork for artificial buzz pollination is independent of instrument buzzing frequency. BMC Ecol. 2020, 20, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, J.A. The dehiscence of anthers by apical pores. In Missouri Botanical Garden Sixteenth Annual Report; Missouri Botanical Garden Press: St. Louis, MO, USA, 1905; pp. 167–257. [Google Scholar]
- Teppner, H. The first records of vibratory pollen-collection by bees. Phyton Horn 2018, 57, 135–141. [Google Scholar]
- Vallejo-Marín, M.; Vallejo, G.C. Comparison of defence buzzes in hoverflies and buzz-pollinating bees. J. Zool. 2021, 313, 237–249. [Google Scholar] [CrossRef]
- Pritchard, D.J.; Vallejo-Marín, M. Floral vibrations by buzz-pollinating bees achieve higher frequency, velocity and acceleration than flight and defence vibrations. J. Exp. Biol. 2020, 223, jeb220541. [Google Scholar] [CrossRef] [PubMed]
- Switzer, C.M.; Combes, S.A. Bumblebee sonication behavior changes with plant species and environmental conditions. Apidologie 2016, 48, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Morgan, T.; Whitehorn, P.; Lye, G.C.; Vallejo-Marín, M. Floral Sonication is an Innate Behaviour in Bumblebees that can be Fine-Tuned with Experience in Manipulating Flowers. J. Insect Behav. 2016, 29, 233–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellinger, A.S.; Chartier, M.; Fernández-Fernández, D.; Penneys, D.S.; Alvear, M.; Almeda, F.; Michelangeli, F.A.; Staedler, Y.; Armbruster, W.S.; Schönenberger, J. Beyond buzz-pollination—Departures from an adaptive plateau lead to new pollination syndromes. New Phytol. 2019, 221, 1136–1149. [Google Scholar] [CrossRef] [Green Version]
- Dellinger, A.S.; Artuso, S.; Pamperl, S.; Michelangeli, F.A.; Penneys, D.S.; Fernández-Fernández, D.M.; Alvear, M.; Almeda, F.; Armbruster, W.S.; Staedler, Y.; et al. Author Correction: Modularity increases rate of floral evolution and adaptive success for functionally specialized pollination systems. Commun. Biol. 2019, 2. [Google Scholar] [CrossRef]
- Bochorny, T.; Bacci, L.F.; Dellinger, A.S.; Michelangeli, F.A.; Goldenberg, R.; Brito, V.L.G. Connective appendages in Huberia bradeana (Melastomataceae) affect pollen release during buzz pollination. Plant Biol. 2021, 23, 556–563. [Google Scholar] [CrossRef]
- Nevard, L.; Russell, A.L.; Foord, K.; Vallejo-Marín, M. Transmission of bee-like vibrations in buzz-pollinated plants with different stamen architectures. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- De Luca, P.A.; Bussière, L.F.; Souto-Vilaros, D.; Goulson, D.; Mason, A.C.; Vallejo-Marín, M. Variability in bumblebee pollination buzzes affects the quantity of pollen released from flowers. Oecologia 2013, 172, 805–816. [Google Scholar] [CrossRef]
- Boyd, J.W.; Murray, D.S.; Tyrl, R.J. Silverleaf nightshade, Solarium elaeagnifolium, origin, distribution, and relation to man. Econ. Bot. 1984, 38, 210–217. [Google Scholar] [CrossRef]
- Travlos, I.S. Responses of invasive silverleaf nightshade (Solanum elaeagnifolium) populations to varying soil water availability. Phytoparasitica 2013, 41, 41–48. [Google Scholar] [CrossRef]
- Petanidou, T.; Price, M.V.; Bronstein, J.L.; Kantsa, A.; Tscheulin, T.; Kariyat, R.; Krigas, N.; Mescher, M.C.; De Moraes, C.M.; Waser, N.M. Pollination and reproduction of an invasive plant inside and outside its ancestral range. Acta Oecologica 2018, 89, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Qasem, J.R.; Al Abdallat, A.M.; Hasan, S.M. Genetic diversity of Solanum elaeagnifolium, an invasive problematic weed in Jordan. Weed Res. 2019, 59, 222–234. [Google Scholar] [CrossRef]
- Chavana, J.; Singh, S.; Vazquez, A.; Christoffersen, B.; Racelis, A.; Kariyat, R.R. Local adaptation to continuous mowing makes the noxious weed Solanum elaeagnifolium a superweed candidate by improving fitness and defense traits. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Thinakaran, J.; Pierson, E.; Kunta, M.; Munyaneza, J.E.; Rush, C.M.; Henne, D.C. Silverleaf Nightshade (Solanum elaeagnifolium), a Reservoir Host for ‘Candidatus Liberibacter solanacearum’, the Putative Causal Agent of Zebra Chip Disease of Potato. Plant Dis. 2015, 99, 910–915. [Google Scholar] [CrossRef] [Green Version]
- Cane, J.H. Estimation of bee size using intertegular span (Apoidea). J. Kans. Entomol. Soc. 1987, 60, 145–147. [Google Scholar]
- King, M.J. Buzz foraging mechanism of bumble bees. J. Apic. Res. 1993, 32, 41–49. [Google Scholar] [CrossRef]
- King, M.J.; Buchmann, S.L. Floral Sonication by Bees: Mesosomal Vibration by Bombus and Xylocopa, but Not Apis (Hymenoptera: Apidae), Ejects Pollen from Poricidal Anthers. J. Kans. Entomol. Soc. 2003, 76, 295–305. [Google Scholar]
- Vallejo-Marin, M.; Manson, J.S.; Thomson, J.D.; Barrett, S.C. Division of labour within flowers: Heteranthery, a floral strategy to reconcile contrasting pollen fates. J. Evol. Biol. 2009, 22, 828–839. [Google Scholar] [CrossRef]
- Corbet, S.A.; Huang, S.-Q. Buzz pollination in eight bumblebee-pollinated Pedicularis species: Does it involve vibration-induced triboelectric charging of pollen grains? Ann. Bot. 2014, 114, 1665–1674. [Google Scholar] [CrossRef] [Green Version]
- Buehmann, S.L.; Cane, J.H. Bees assess pollen returns while sonicating Solanum flowers. Oecologia 1989, 81, 289–294. [Google Scholar] [CrossRef]
- Russell, A.L.; Buchmann, S.L.; Papaj, D.R. How a generalist bee achieves high efficiency of pollen collection on diverse floral resources. Behav. Ecol. 2017, 28, 991–1003. [Google Scholar] [CrossRef] [Green Version]
- Rosi-Denadai, C.A.; Araújo, P.C.S.; de Oliveira Campos, L.A.; Cosme, L., Jr.; Guedes, R.N.C. Buzz-pollination in Neotropical bees: Genus-dependent frequencies and lack of optimal frequency for pollen release. Insect Sci. 2020, 27, 133–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mekki, M. Biology, distribution and impacts of silverleaf nightshade (Solanum elaeagnifolium Cav.). EPPO Bull. 2007, 37, 114–118. [Google Scholar] [CrossRef]
- Bellue, M.K. Weed Seed Handbook; Bulletin of the California State Department of Agriculture Series VI; 1946; Volume 35, pp. 15–16. [Google Scholar]
- Cuthbertson, E.G.; Leys, A.R.; McMaster, G. Silverleaf nightshade—A potential threat to agriculture. Agric. Gaz. N. S. W. 1976, 87, 11–13. [Google Scholar]
- Molnar, V.M.; McKenzie, D.N. Progress report on silverleaf nightshade research. Pamphlet No. 61. In Vermin and Noxious Weeds Destruction Report; Board of Crown Lands and Survey: Victoria, Australia, 1976. [Google Scholar]
- Macior, L.W. Pollination adaptation in Pedicularis groenlandica. Am. J. Bot. 1968, 55, 927–932. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bee Genera/Parameters | Exomalopsis | Halictus | Megachile | Xylocopa |
|---|---|---|---|---|
| Bee visitation time | 40 | 40 | 40 | 40 |
| Number of pulses/visits | 18 | 8 | 16 | 22 |
| Buzz % over time | 18 | 8 | 16 | 22 |
| Bee Buzzing frequency and amplitude | 18 | 8 | 16 | 22 |
| Bee size (Bee mass and ITD) | 18 | 8 | 16 | 22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tayal, M.; Kariyat, R. Examining the Role of Buzzing Time and Acoustics on Pollen Extraction of Solanum elaeagnifolium. Plants 2021, 10, 2592. https://doi.org/10.3390/plants10122592
Tayal M, Kariyat R. Examining the Role of Buzzing Time and Acoustics on Pollen Extraction of Solanum elaeagnifolium. Plants. 2021; 10(12):2592. https://doi.org/10.3390/plants10122592
Chicago/Turabian StyleTayal, Mandeep, and Rupesh Kariyat. 2021. "Examining the Role of Buzzing Time and Acoustics on Pollen Extraction of Solanum elaeagnifolium" Plants 10, no. 12: 2592. https://doi.org/10.3390/plants10122592
APA StyleTayal, M., & Kariyat, R. (2021). Examining the Role of Buzzing Time and Acoustics on Pollen Extraction of Solanum elaeagnifolium. Plants, 10(12), 2592. https://doi.org/10.3390/plants10122592