
Solar Radiation Flux Provides a Method of Quantifying Weed-Crop Balance in Present and Future Climates



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sites, Crop Season and Weather Data
2.2. Methodology for Estimating Intercepted and Transmitted Solar Radiation from Ground Cover
2.3. Weed Dry Matter and Species
2.4. Analyses
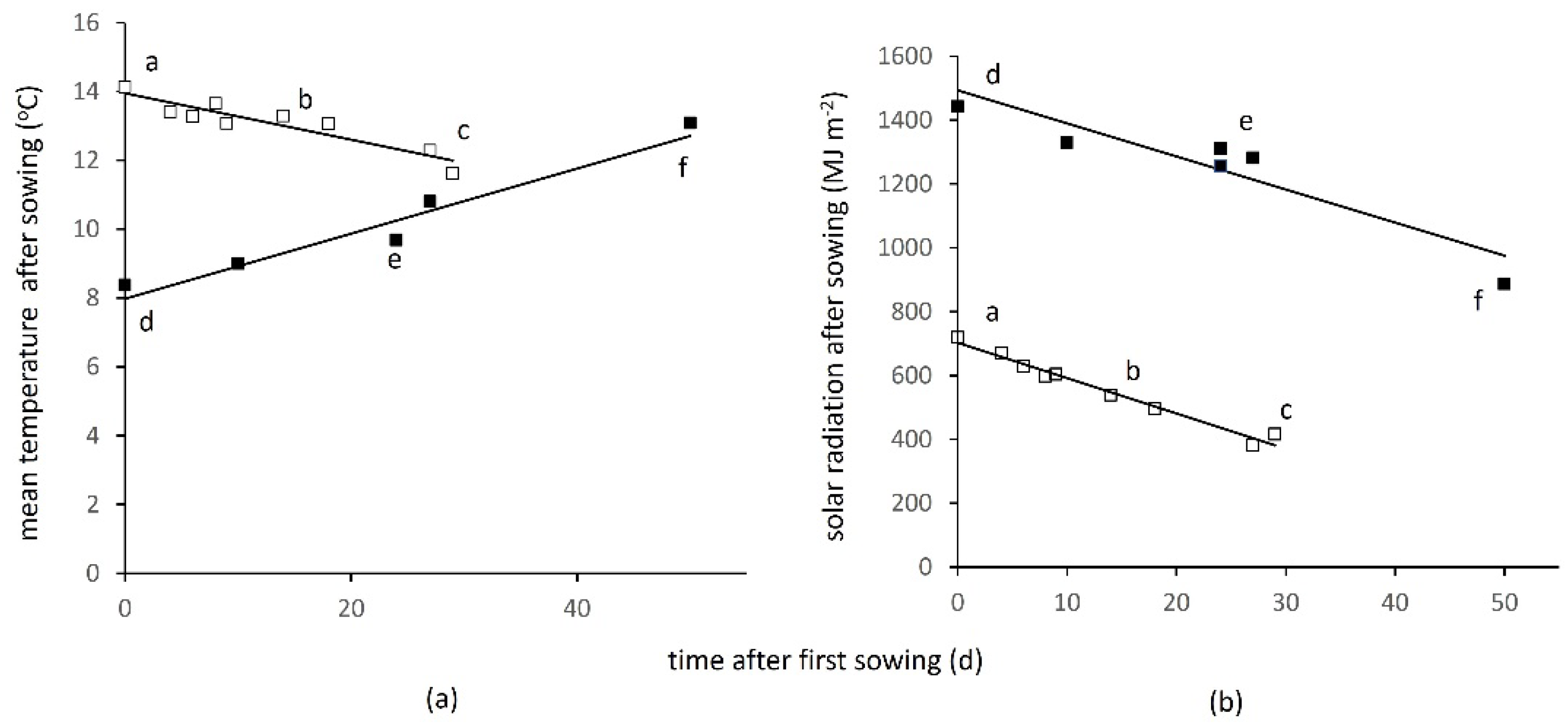
2.5. Interpreting Effects of Sowing Time through Temperature and Solar Radiation
3. Results
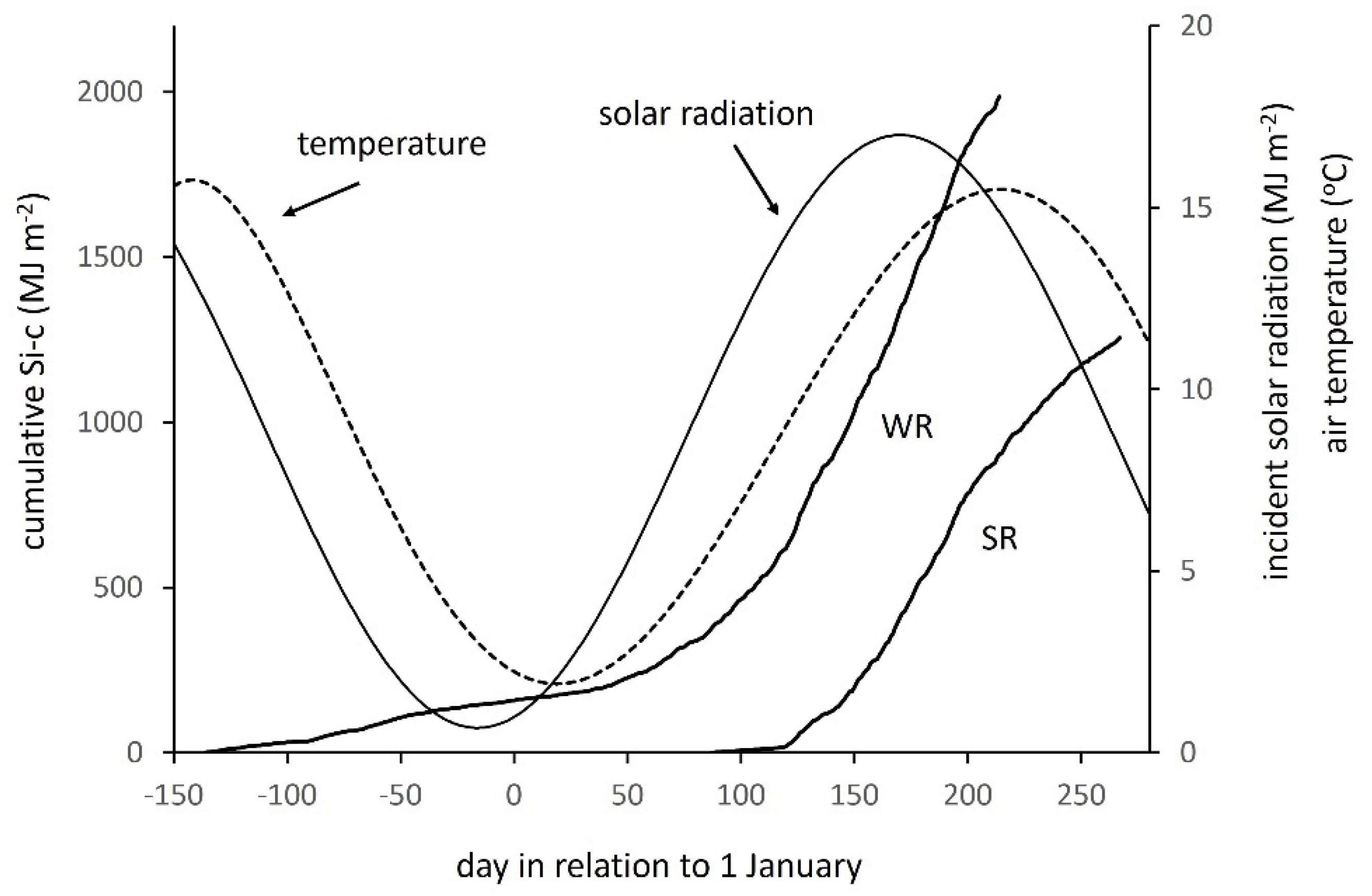
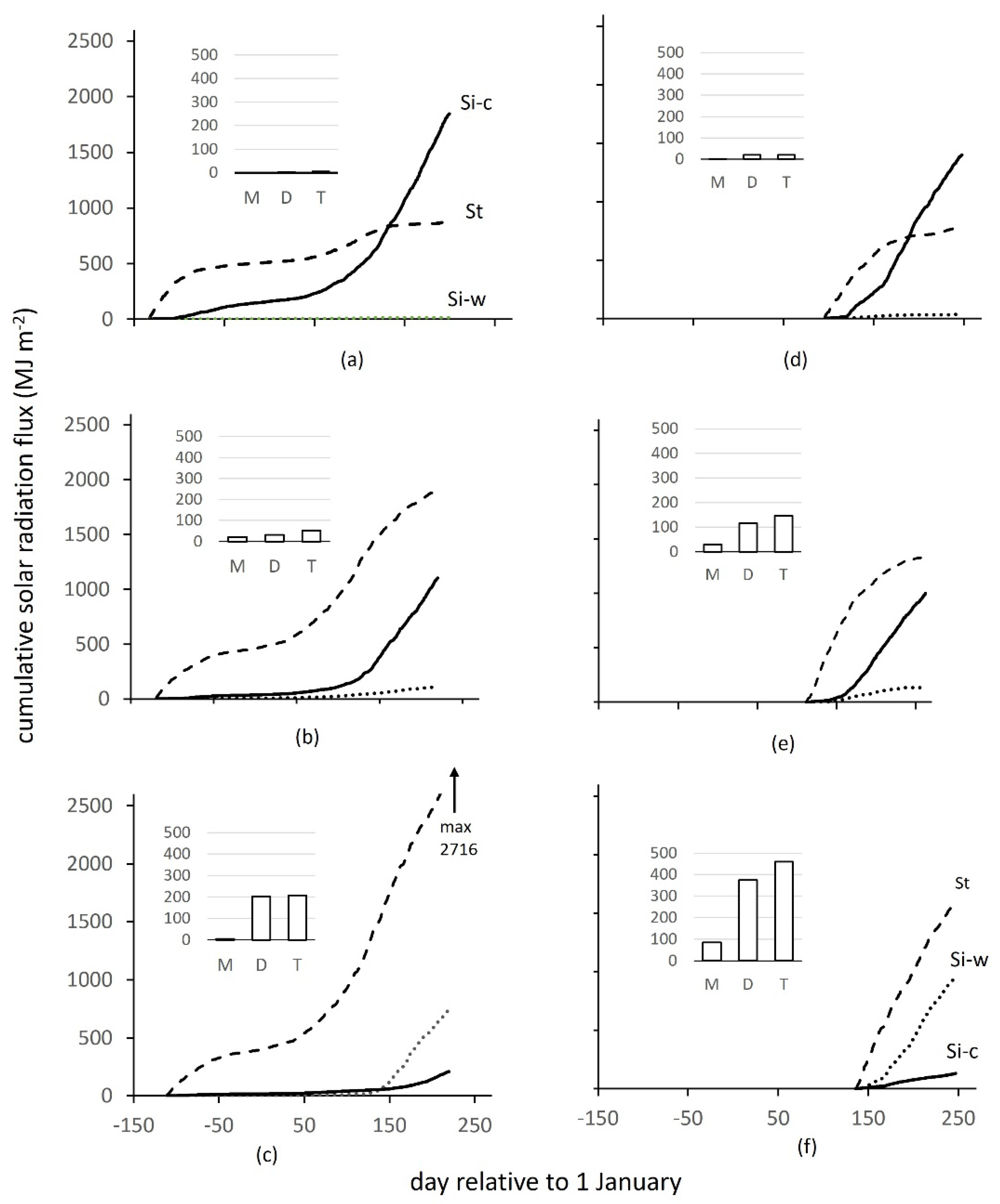
3.1. Time and Temperature Effects on Crop Solar Radiation Flux
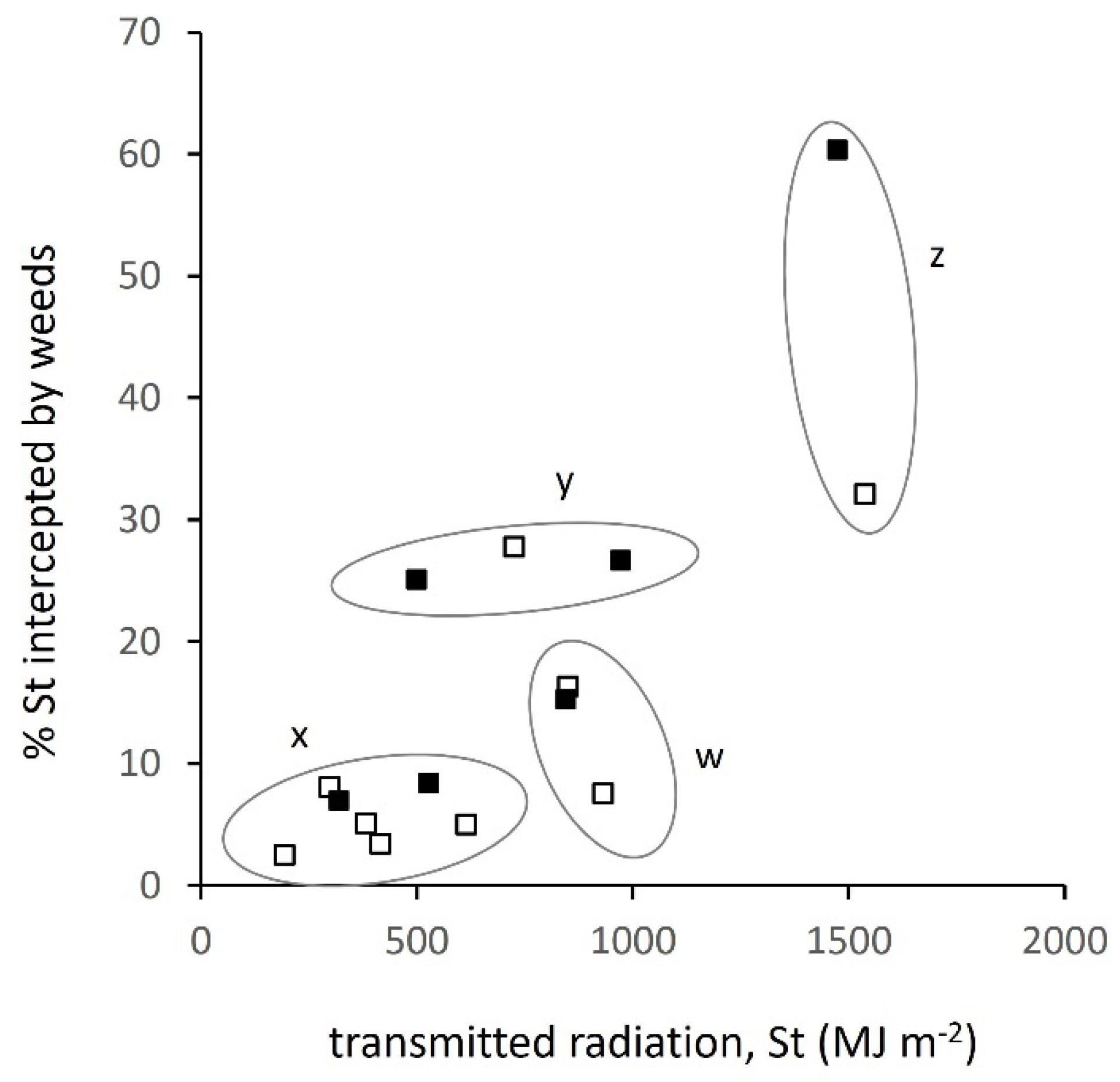
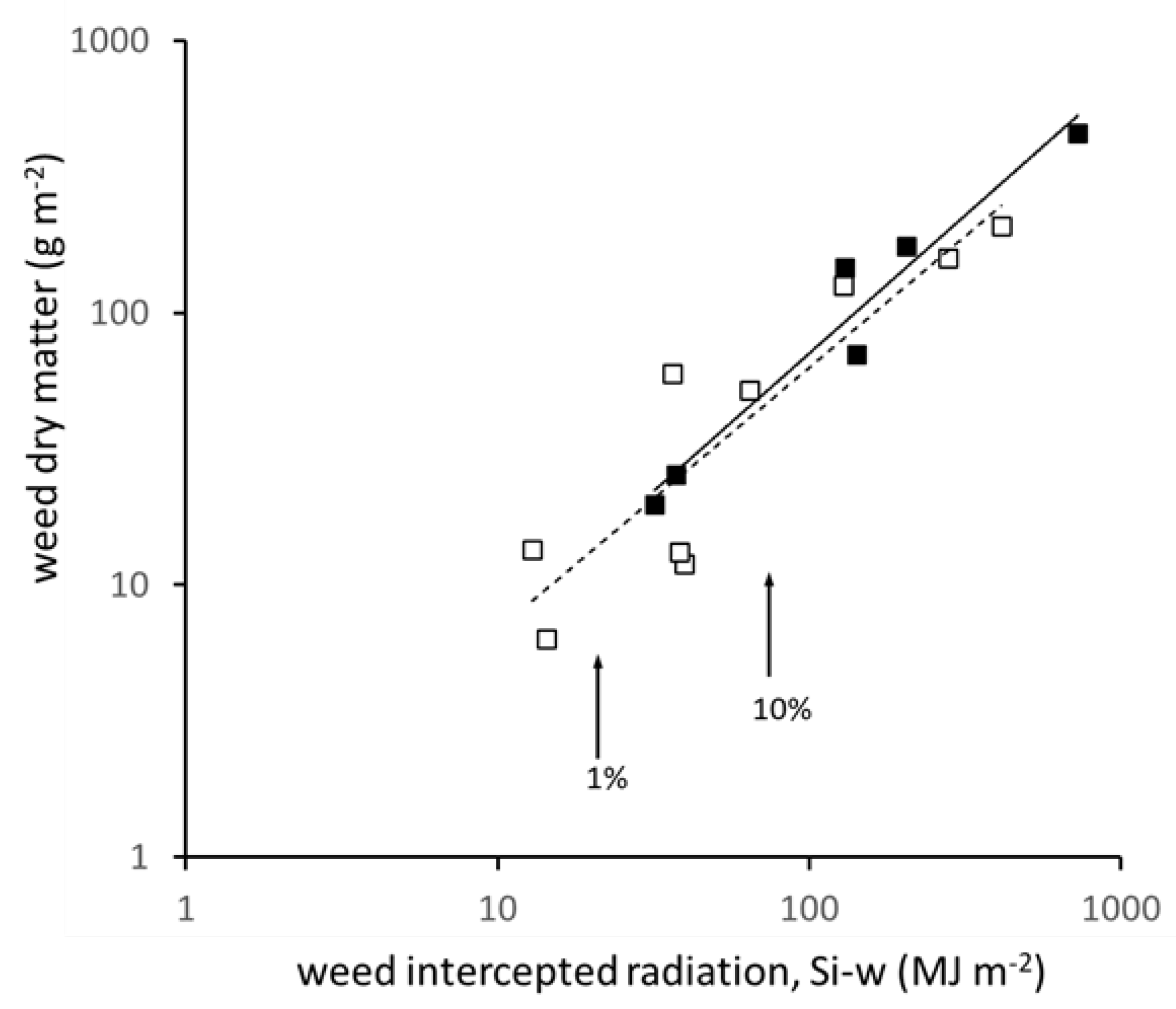
3.2. Relation between Transmitted Radiation and Attributes of the Weed Layer
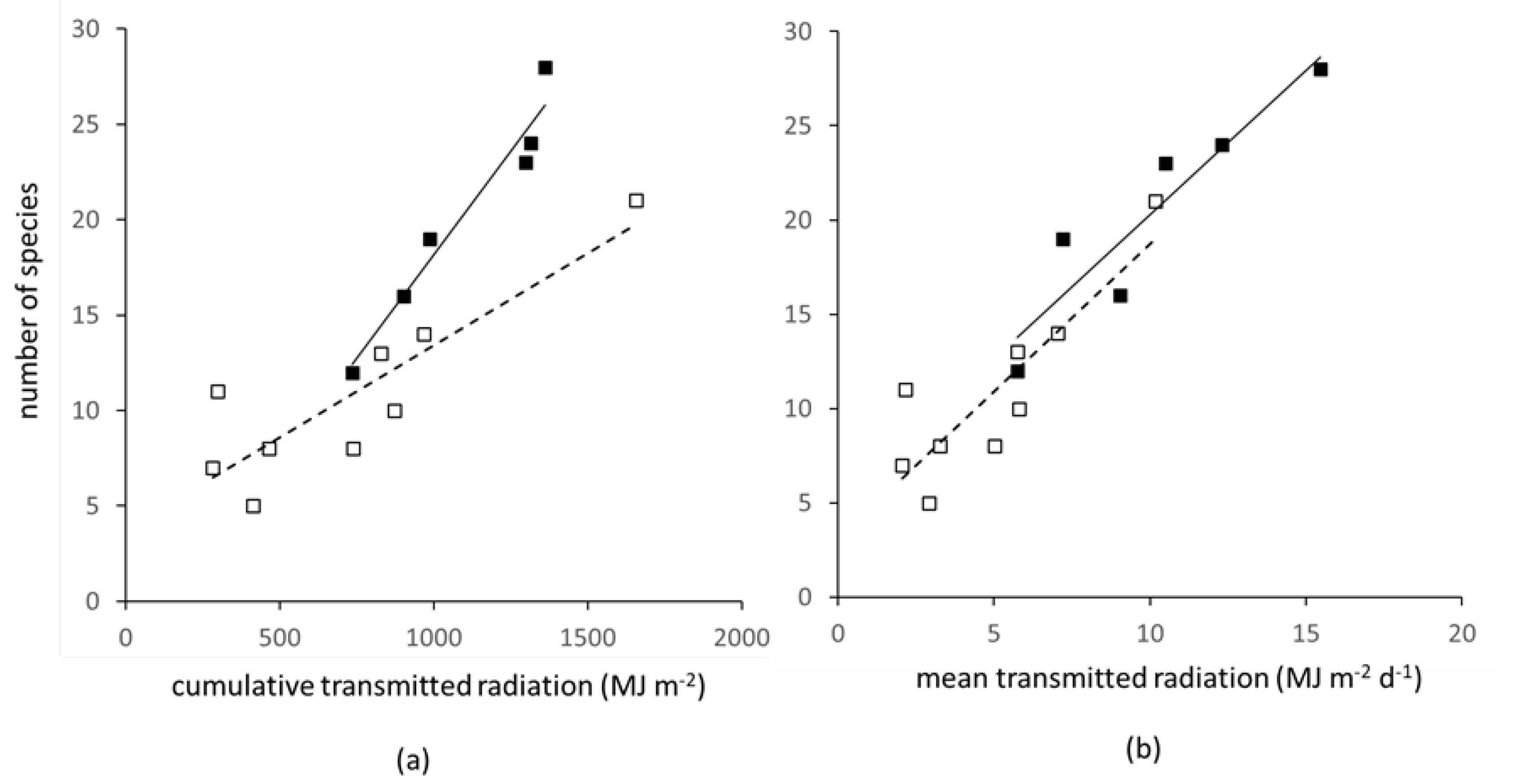
3.3. Determinants of Weed Species Number
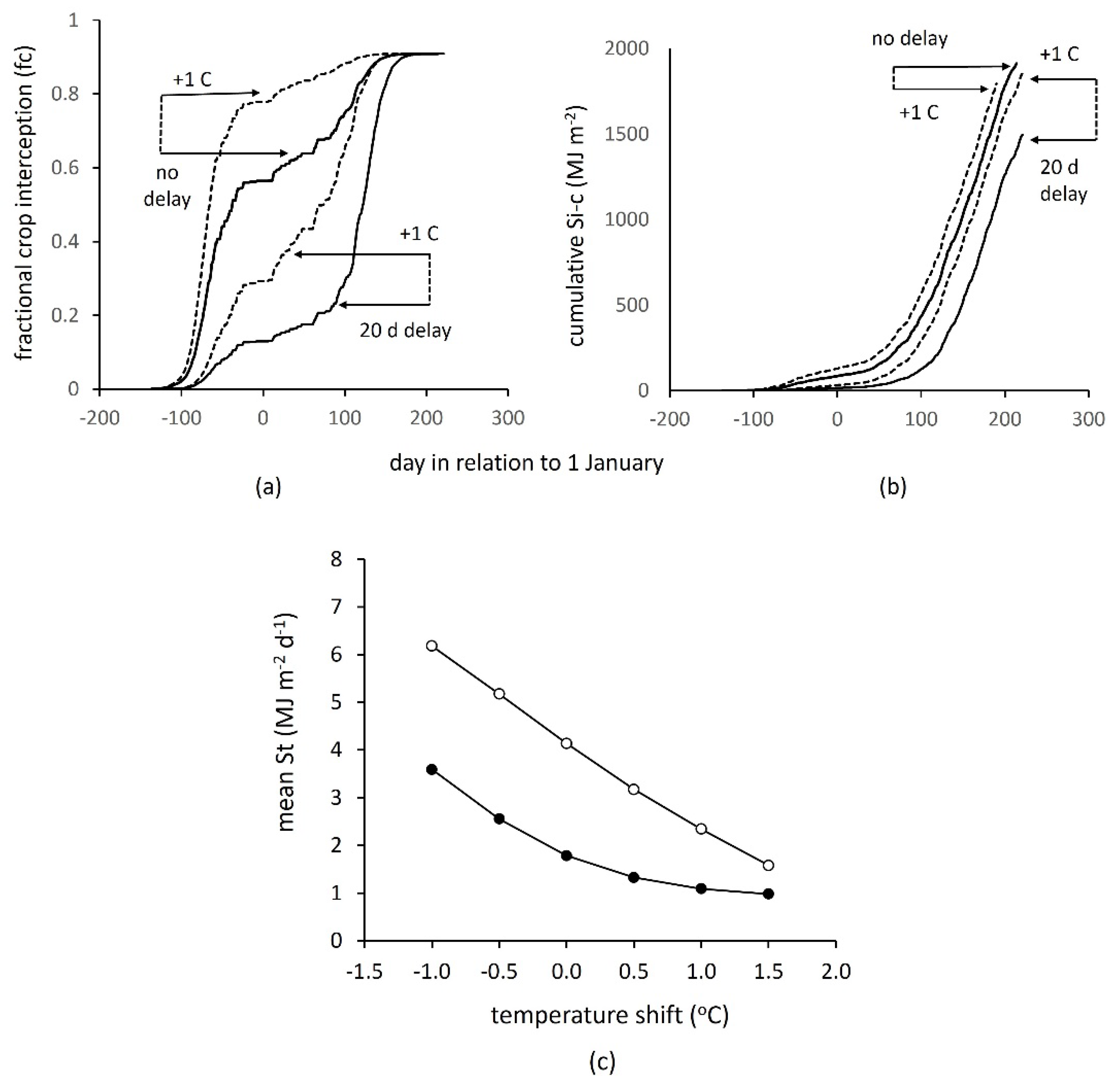
3.4. Interaction between Sowing Time, Temperature and Incident Solar Radiation
4. Discussion
Gauging the Sensitivity of the Crop-Weed Balance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brenchley, W.E. Weeds of Farm Land; Longmans, Green and Co.: London, UK, 1920. [Google Scholar]
- Froud-Williams, R.J.; Chancellor, R.J.; Drennan, D.S.H. Influence of cultivation regime upon buried weed seeds in arable cropping systems. J. Appl. Ecol. 1983, 20, 199–208. [Google Scholar] [CrossRef]
- Squire, G.R. Defining sustainable limits during and after intensification in a maritime agricultural ecosystem. Ecosyst. Health Sustain. 2017, 3, 1368873. [Google Scholar] [CrossRef] [Green Version]
- Norris, R.F.; Kogan, M. Interactions between weeds, arthropod pests and their natural enemies in managed ecosystems. Weed Sci. 2000, 48, 94–158. [Google Scholar] [CrossRef]
- Hawes, C.; Squire, G.R.; Hallett, P.D.; Watson, C.A.; Young, M. Arable plant communities as indicators of farming practice. Agric. Ecosyst. Environ. 2010, 138, 17–26. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Brown, V.K.; Boatman, N.D.; Lutman, P.J.W.; Squire, G.R.; Ward, L.K. The role of weeds in supporting biological diversity within cropped fields. Weed Res. 2003, 43, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Storkey, J.; Westbury, D.B. Managing arable weeds for biodiversity. Pest. Manag. Sci. 2007, 63, 517–523. [Google Scholar] [CrossRef]
- Bàrberi, P.; Burgio, G.; Dinelli, G.; Moonen, A.C.; Otto, S.; Vazzana, C.; Zanin, G. Functional biodiversity in the agricultural landscape: Relationships between weeds and arthropod fauna. Weed Res. 2010, 50, 388–401. [Google Scholar] [CrossRef]
- Smith, B.M.; Aebisher, N.J.; Ewald, J.; Moreby, S.; Potter, C.; Holland, J.M. The potential for arable weeds to reverse invertebrate declines and associated ecosystem services in cereal crops. Front. Sustain. Food Syst. 2020, 3, 118. [Google Scholar] [CrossRef]
- MacLaren, C.; Storkey, J.; Menegat, A.; Metcalfe, H.; Dehnen-Schmutz, K. A ecological future for weed science to sustain crop production and the environment. A review. Agron Sustain. Dev. 2020, 40, 24. [Google Scholar] [CrossRef]
- Ramesh, K.; Matloob, A.; Aslam, F.; Florentine, S.K.; Chauhan, B.S. Weeds in a changing climate: Vulnerabilities, consequences, and implications for future weed management. Front. Plant Sci. 2017, 8, 95. [Google Scholar] [CrossRef]
- Patterson, D.T. Weeds in a changing climate. Weed Sci. 1995, 43, 685–701. [Google Scholar] [CrossRef]
- Peters, K.; Breitsameter, L.; Gerowitt, B. Impact of climate change on weeds in agriculture: A review. Agron. Sustain. Dev. 2014, 34, 707–721. [Google Scholar] [CrossRef] [Green Version]
- Ziska, L.; McConnell, L.L. Climate change, carbon dioxide and pest biology: Monitor, mitigate, manage. J. Agric. Food Chem. 2016, 64, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Bastiaans, L.; Paolini, R.; Baumann, D.T. Focus on ecological weed management: What is hindering adoption? Weed Res. 2008, 48, 481–491. [Google Scholar] [CrossRef]
- Holst, N.; Rasmussen, I.A.; Bastiaans, L. Field weed population dynamics: A review of model approaches and applications. Weed Res. 2007, 47, 1–14. [Google Scholar] [CrossRef]
- Hawes, C.; Iannetta, P.P.M.; Squire, G.R. Agroecological practices for whole-system sustainability. CAB Rev. 2021, 16, 1–19. [Google Scholar] [CrossRef]
- Squire, G.R. Modelling biodiversity. In Reconciling Agricultural Production with Biodiversity Conservation, Chapter 2; Bárberi, P., Moonen, A.-C., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK, 2020. [Google Scholar] [CrossRef]
- Colbach, N.; Colas, F.; Cordeau, S.; Maillot, T.; Queyrel, W.; Villerd, J.; Moreau, D. The FLORSYS crop-weed canopy model, a tool to investigate and promote agroecologcal weed management. Field Crop. Res. 2021, 261, 108006. [Google Scholar] [CrossRef]
- Pascher, K.; Moser, D.; Dullinger, S.; Sachslehner, L.; Hottinger, L.; Traxler, A.; Sauberer, N.; Frank, T.; Grabherr, G. Monitoring design to evaluate biodiversity in Austrian agricultural regions. In Implications of GM-Crop Cultivation at Large Spatial Scales; Breckling, B., Reuter, H., Verhoeven, R., Eds.; Theorie in der Okologie. Peter Lang: Frankfurt, Germany, 2008; pp. 146–150. [Google Scholar]
- Squire, G.R.; Hawes, C.; Begg, G.S.; Young, M.W. Cumulative impact of GM herbicide-tolerant cropping on arable plants assessed through species-based and functional taxonomies. Environ. Sci. Pollut. R. 2009, 16, 85–94. [Google Scholar] [CrossRef]
- Monteith, J.L. Solar energy and productivity in tropical ecosystems. J. Appl. Ecol. 1972, 9, 747–766. [Google Scholar] [CrossRef] [Green Version]
- Monteith, J.L. Climate and the efficiency of crop production in Britain. Philos. Trans. R. Soc. Lond. B 1977, 281, 277–294. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Muchow, R.C. Radiation use efficiency. Adv. Agron. 1999, 65, 215–265. [Google Scholar] [CrossRef]
- Squire, G.R. Limits to crop production. In Food Production and Nature Conservation: Conflicts and Solutions; Gordon, I.J., Prins, H.T., Squire, G.R., Eds.; Routledge: Oxford, UK, 2017; pp. 41–67. [Google Scholar]
- Marshall, B.; Willey, R.W. Radiation interception and growth in an intercrop of pearl millet/groundnut. Field Crop. Res. 1983, 7, 141–160. [Google Scholar] [CrossRef]
- Boote, K.J.; Jones, J.W.; Hoogenboom, G.; White, J.W. The role of crop systems simulation in agriculture and environment. Int. J. Agric. Environ. Inf. Syst. 2010, 1, 41–54. [Google Scholar] [CrossRef]
- Steven, M.D.; Biscoe, P.V.; Jaggard, K.W.; Paruntu, J. Foliage cover and radiation interception. Field Crop. Res. 1986, 13, 75–87. [Google Scholar] [CrossRef]
- Franklin, B.T. A History of Scottish Farming; Thomas Nelson and Sons: London, UK, 1952. [Google Scholar]
- Long, H.C. Poisonous Plants on the Farm; Ministry of Agriculture and Fisheries, Miscellaneous Publications 57: London, UK, 1927. [Google Scholar]
- Weeds, A.C.T. Legislation.gov.uk. 1959. Available online: https://www.legislation.gov.uk/ukpga/Eliz2/7-8/54/introduction (accessed on 19 November 2021).
- Squire, G.R.; Rodger, S.; Wright, G.W. Community-scale seedbank response to less intense rotation and reduced herbicide input at three sites. Ann. Appl. Biol. 2000, 136, 47–57. [Google Scholar] [CrossRef]
- Climate Change Committee 2021 Independent Assessment of UK Climate Risk: Advice to Government for the UK’s Third Climate Change Risk Assessment. Available online: https://www.theccc.org.uk/publication/independent-assessment-of-uk-climate-risk (accessed on 19 November 2021).
- Hawes, C.; Haughton, A.J.; Osborne, J.L.; Roy, D.B.; Clark, S.J.; Perry, J.N.; Rothery, P.; Bohan, D.A.; Brooks, D.R.; Champion, G.T.; et al. Responses of plant and invertebrate trophic groups to contrasting herbicide regimes in the Farm Scale Evaluations of genetically-modified herbicide-tolerant crops. Philos. Trans. R. Soc. B 2003, 358, 1899–1913. [Google Scholar]
- Debeljak, M.; Squire, G.R.; Kocev, D.; Hawes, C.; Young, M.W.; Džeroski, S. Analysis of time series data on agroecosystem vegetation using predictive clustering trees. Ecol. Model 2011, 222, 2524–2529. [Google Scholar] [CrossRef]
- Mendham, N.J.; Shipway, P.A.; Scott, R.K. The effects of delayed sowing and weather on growth, development and yield of winter oilseed rape Brassica napus. J. Agr. Sci.—Camb. 1981, 96, 389–416. [Google Scholar] [CrossRef]
- Champion, G.T.; May, M.J.; Bennett, S.; Brooks, D.R.; Clark, S.J.; Daniels, R.E.; Firbank, L.G.; Haughton, A.J.; Hawes, C.; Heard, M.S.; et al. Crop management and agronomic context of the Farm Scale Evaluations of genetically modified herbicide-tolerant crops. Philos. Trans. R. Soc. B 2003, 358, 1801–1818. [Google Scholar] [CrossRef] [Green Version]
- Page, J.; Albuisson, M.; Wald, L. The European solar radiation atlas: A valuable digital tool. Sol. Energy 2001, 71, 81–83. [Google Scholar] [CrossRef] [Green Version]
- Met Office. UK and Regional Series Weather Observations. 2021. Available online: https://www.metoffice.gov.uk/research/climate/maps-and-data/uk-and-regional-series (accessed on 19 November 2021).
- Fertiliser Practice. 2020 British Survey of Fertiliser Practice and All Previous Yearbooks. Available online: https://www.gov.uk/government/statistics/british-survey-of-fertiliser-practice-2020 (accessed on 19 November 2021).
- PUS. Pesticide Use Survey Reports for 2020 and Previous Years; Science and Advice for Scottish Agriculture: Edinburgh, UK, 2020. Available online: https://www.sasa.gov.uk/pesticides/pesticide-usage/pesticide-usage-survey-reports (accessed on 19 November 2021).
- Kempton, R.A. The structure of species abundance and measurement of diversity. Biometrics 1979, 35, 307–321. [Google Scholar] [CrossRef]
- Fernandez-Pascual, E.; Mattana, E.; Pritchard, H.W. Seeds of future past: Climate change and the thermal memory of plant reproductive traits. Biol. Rev. 2019, 94, 439–456. [Google Scholar] [CrossRef] [PubMed]
- Marshall, B.; Squire, G.R.; Terry, A.C. Effect of temperature on interception and conversion of solar radiation by stands of groundnut. J. Exp. Bot. 1992, 43, 95–101. [Google Scholar] [CrossRef]
- Young, M.W.; Mullins, E.; Squire, G.R. Environmental risk assessment of blight-resistant potato: Use of a crop model to quantify nitrogen cycling at scales of the field and cropping system. Environ. Sci. Pollut. Res. 2017, 24, 21434–21444. [Google Scholar] [CrossRef] [PubMed]
- Monteith, J.L. Climatic variation and the growth of crops. Q. J. Roy. Met. Soc. 1981, 107, 749–774. [Google Scholar] [CrossRef]
- Habekotte, B. Options for increasing seed yield of winter oilseed rape Brassica napus L.: A simulation study. Field Crop. Res. 1997, 54, 109–126. [Google Scholar] [CrossRef]
- World Meteorological Organisation. 2021 State of Climate in Africa 2020. Available online: https://public.wmo.int/en/our-mandate/climate/wmo-statement-state-of-global-climate/Africa (accessed on 19 November 2021).
- Squire, G.R.; Cammarano, D.; Daniell, T.J.; Newton, A.C.; White, P.J. Agriculture and Forestry Climate Change Report Card Technical Paper. 3 Arable Crops. UK Natural Environment Research Council. 2016. Available online: https://nerc.ukri.org/research/partnerships/ride/lwec/report-cards/ (accessed on 19 November 2021).
- Cammarano, D.; Hawes, C.; Squire, G.; Holland, J.; Rivington, M.; Murgia, T.; Roggero, P.P.; Fontana, F.; Casa, R.; Ronga, D. Rainfall and temperature impacts on barle5y Hordeum vulgare L. yield and malting quality in Scotland. Field Crop. Res. 2019, 241, 107559. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Squire, G.R.; Young, M.W.; Hawes, C. Solar Radiation Flux Provides a Method of Quantifying Weed-Crop Balance in Present and Future Climates. Plants 2021, 10, 2657. https://doi.org/10.3390/plants10122657
Squire GR, Young MW, Hawes C. Solar Radiation Flux Provides a Method of Quantifying Weed-Crop Balance in Present and Future Climates. Plants. 2021; 10(12):2657. https://doi.org/10.3390/plants10122657
Chicago/Turabian StyleSquire, Geoffrey R., Mark W. Young, and Cathy Hawes. 2021. "Solar Radiation Flux Provides a Method of Quantifying Weed-Crop Balance in Present and Future Climates" Plants 10, no. 12: 2657. https://doi.org/10.3390/plants10122657
APA StyleSquire, G. R., Young, M. W., & Hawes, C. (2021). Solar Radiation Flux Provides a Method of Quantifying Weed-Crop Balance in Present and Future Climates. Plants, 10(12), 2657. https://doi.org/10.3390/plants10122657