
The Potential of Rhizoctonia-Like Fungi for the Biological Protection of Cereals against Fungal Pathogens


Abstract
:1. Introduction
2. Results
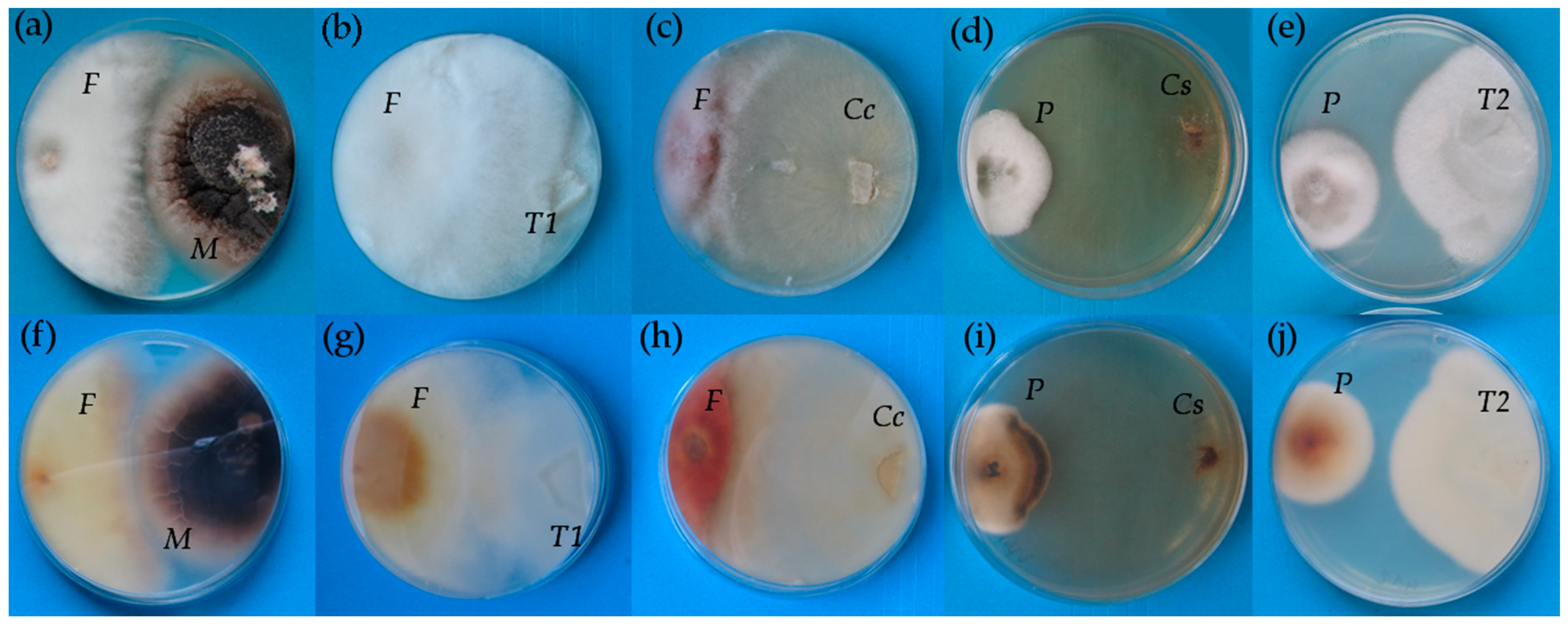
2.1. Competitive In Vitro Test
2.2. Pot Experiments
2.2.1. Experiment 1
2.2.2. Experiment 2
2.2.3. Experiment 3
2.3. Field Experiment
3. Discussion
4. Materials and Methods
4.1. Identification of Isolates and Inoculum Preparation
4.2. Competitive In Vitro Test
4.3. Pot Experiments in a Growth Chamber
4.3.1. Experiment 1
4.3.2. Experiment 2
4.3.3. Experiment 3
4.3.4. Evaluation
4.4. Field Experiment
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Williams, P.G. Orchidaceous Rhizoctonias in Pot Cultures of Vesicular–Arbuscular Mycorrhizal Fungi. Can. J. Bot. 1985, 63, 1329–1333. [Google Scholar] [CrossRef]
- Bokati, D. The Cryptic Sebacinales: An Obscure but Ubiquitous Group of Root Symbionts Comes to Light. Fungal Ecol. 2016, 2, 115–119. [Google Scholar] [CrossRef]
- Weiß, M.; Waller, F.; Zuccaro, A.; Selosse, M. Sebacinales—One Thousand and One Interactions with Land Plants. New Phytol. 2016, 211, 20–40. [Google Scholar] [CrossRef] [PubMed]
- Dearnaley, J.D.W.; Martos, F.; Selosse, M.-A. 12 Orchid Mycorrhizas: Molecular Ecology, Physiology, Evolution and Conservation Aspects. In Fungal Associations; Hock, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 207–230. ISBN 978-3-642-30825-3. [Google Scholar]
- Moore, R. The Genera of Rhizoctonia-like Fungi: Ascorhizoctonia, Ceratorhiza Gen. Nov., Epulorhiza Gen. Nov., Moniliopsis, and Rhizoctonia. Mycotaxon 1987, 29, 91–99. [Google Scholar]
- Oberwinkler, F.; Riess, K.; Bauer, R.; Selosse, M.-A.; Weiß, M.; Garnica, S.; Zuccaro, A. Enigmatic Sebacinales. Mycol. Prog. 2013, 12, 1–27. [Google Scholar] [CrossRef]
- Weiß, M.; Sýkorová, Z.; Garnica, S.; Riess, K.; Martos, F.; Krause, C.; Oberwinkler, F.; Bauer, R.; Redecker, D. Sebacinales Everywhere: Previously Overlooked Ubiquitous Fungal Endophytes. PLoS ONE 2011, 6, e16793. [Google Scholar] [CrossRef] [PubMed]
- Gónzalez, D.; Rodriguez-Carres, M.; Boekhout, T.; Stalpers, J.; Kuramae, E.E.; Nakatani, A.K.; Vilgalys, R.; Cubeta, M.A. Phylogenetic Relationships of Rhizoctonia Fungi within the Cantharellales. Fungal Biol. 2016, 120, 603–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warcup, J.H. Mycorrhizal Associations of Isolates of Sebacina Vermifera. New Phytol. 1988, 110, 227–231. [Google Scholar] [CrossRef]
- Oberwinkler, F.; Riess, K.; Bauer, R.; Garnica, S. Morphology and Molecules: The Sebacinales, a Case Study. Mycol. Prog. 2014, 13, 445–470. [Google Scholar] [CrossRef]
- Verma, S.; Varma, A.; Rexer, K.-H.; Hassel, A.; Kost, G.; Sarbhoy, A.; Bisen, P.; Bütehorn, B.; Franken, P. Piriformospora Indica, Gen. et Sp. Nov., a New Root-Colonizing Fungus. Mycologia 1998, 90, 896–903. [Google Scholar] [CrossRef]
- Shoresh, M.; Harman, G.E.; Mastouri, F. Induced Systemic Resistance and Plant Responses to Fungal Biocontrol Agents. Annu. Rev. Phytopathol. 2010, 48, 21–43. [Google Scholar] [CrossRef] [Green Version]
- Waller, F.; Achatz, B.; Baltruschat, H.; Fodor, J.; Becker, K.; Fischer, M.; Heier, T.; Huckelhoven, R.; Neumann, C.; von Wettstein, D.; et al. The Endophytic Fungus Piriformospora Indica Reprograms Barley to Salt-Stress Tolerance, Disease Resistance, and Higher Yield. Proc. Natl. Acad. Sci. USA 2005, 102, 13386–13391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Card, S.; Johnson, L.; Teasdale, S.; Caradus, J. Deciphering Endophyte Behaviour: The Link between Endophyte Biology and Efficacious Biological Control Agents. FEMS Microbiol. Ecol. 2016, 92, fiw114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Látalová, K.; Baláž, M. Carbon Nutrition of Mature Green Orchid Serapias Strictiflora and Its Mycorrhizal Fungus Epulorhiza sp. Biol. Plant. 2010, 54, 97–104. [Google Scholar] [CrossRef]
- Rodriguez, R.J., Jr.; Arnold, A.E.; Redman, R.S. Fungal Endophytes: Diversity and Functional Roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Brader, G.; Compant, S.; Vescio, K.; Mitter, B.; Trognitz, F.; Ma, L.-J.; Sessitsch, A. Ecology and Genomic Insights into Plant-Pathogenic and Plant-Nonpathogenic Endophytes. Annu. Rev. Phytopathol. 2017, 55, 61–83. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.-H.; Tam, S.-L.; Toda, T.; Chen, L.-C. Controlling Rhizoctonia Damping-off of Chinese Mustard by Using Endomycorrhizal Rhizoctonia Spp. Isolated from Orchid Mycorrhizae. Plant Dis. 2016, 100, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Mosquera-Espinosa, A.T.; Bayman, P.; Prado, G.A.; Gómez-Carabalí, A.; Otero, J.T. The Double Life of Ceratobasidium: Orchid Mycorrhizal Fungi and Their Potential for Biocontrol of Rhizoctonia Solani Sheath Blight of Rice. Mycologia 2013, 105, 141–150. [Google Scholar] [CrossRef]
- Bungtongdee, N.; Sopalun, K.; Laosripaiboon, W.; Iamtham, S. The Chemical Composition, Antifungal, Antioxidant and Antimutagenicity Properties of Bioactive Compounds from Fungal Endophytes Associated with Thai Orchids. J. Phytopathol. 2019, 167, 56–64. [Google Scholar] [CrossRef]
- O’Brien, P.A. Biological Control of Plant Diseases. Australas. Plant Pathol. 2017, 46, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.; Haber, S. Overview of Some Recent Research Developments in Fusarium Head Blight of Wheat. Can. J. Plant Pathol. 2013, 35, 149–174. [Google Scholar] [CrossRef]
- Sanjay, R.; Ponmurugan, P.; Baby, U.I. Evaluation of Fungicides and Biocontrol Agents against Grey Blight Disease of Tea in the Field. Crop Prot. 2008, 27, 689–694. [Google Scholar] [CrossRef]
- Silva, V.; Mol, H.G.J.; Zomer, P.; Tienstra, M.; Ritsema, C.J.; Geissen, V. Pesticide Residues in European Agricultural Soils—A Hidden Reality Unfolded. Sci. Total Environ. 2019, 653, 1532–1545. [Google Scholar] [CrossRef]
- Harman, G.E. Multifunctional Fungal Plant Symbionts: New Tools to Enhance Plant Growth and Productivity: Commentary. New Phytol. 2011, 189, 647–649. [Google Scholar] [CrossRef]
- Jaber, L.R. Seed Inoculation with Endophytic Fungal Entomopathogens Promotes Plant Growth and Reduces Crown and Root Rot (CRR) Caused by Fusarium Culmorum in Wheat. Planta 2018, 248, 1525–1535. [Google Scholar] [CrossRef]
- Kuga, Y.; Sakamoto, N.; Yurimoto, H. Stable Isotope Cellular Imaging Reveals That Both Live and Degenerating Fungal Pelotons Transfer Carbon and Nitrogen to Orchid Protocorms. New Phytol. 2014, 202, 594–605. [Google Scholar] [CrossRef] [PubMed]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. Fusarium Culmorum: Causal Agent of Foot and Root Rot and Head Blight on Wheat: The Wheat Pathogen. Fusarium Culmorum. Mol. Plant Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef]
- Tóth, B.; Kászonyi, G.; Bartók, T.; Varga, J.; Mesterházy, Á. Common Resistance of Wheat to Members of the Fusarium Graminearum Species Complex and F. Culmorum: Common Wheat Resistance to Fusarium Graminearum Species Complex. Plant Breed. 2008, 127, 1–8. [Google Scholar] [CrossRef]
- Hogg, A.C.; Johnston, R.H.; Johnston, J.A.; Klouser, L.; Kephart, K.D.; Dyer, A.T. Monitoring Fusarium Crown Rot Populations in Spring Wheat Residues Using Quantitative Real-Time Polymerase Chain Reaction. Phytopathology 2010, 100, 49–57. [Google Scholar] [CrossRef] [Green Version]
- McMullen, M.; Bergstrom, G.; De Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A Unified Effort to Fight an Enemy of Wheat and Barley: Fusarium Head Blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef] [Green Version]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From Simple to Complex Mycotoxins. Toxins 2011, 3, 802. [Google Scholar] [CrossRef]
- Walter, S.; Nicholson, P.; Doohan, F.M. Action and Reaction of Host and Pathogen during Fusarium Head Blight Disease. New Phytol. 2010, 185, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Mechanisms of Action, Human Exposure, and Toxicological Relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Ryu, D. Worldwide Occurrence of Mycotoxins in Cereals and Cereal-Derived Food Products: Public Health Perspectives of Their Co-Occurrence. J. Agric. Food Chem. 2017, 18, 7034–7051. [Google Scholar] [CrossRef]
- Al-Hatmi, A.M.S. Current Antifungal Treatment of Fusariosis. Int. J. Antimicrob. Agents 2018, 7, 326–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czembor, E.; Stępień, Ł.; Waśkiewicz, A. Effect of Environmental Factors on Fusarium Species and Associated Mycotoxins in Maize Grain Grown in Poland. PLoS ONE 2015, 10, e0133644. [Google Scholar] [CrossRef]
- Dweba, C.C. Fusarium Head Blight of Wheat: Pathogenesis and Control Strategies. Crop Prot. 2017, 9, 114–122. [Google Scholar] [CrossRef]
- Antalová, Z.; Bleša, D.; Martinek, P.; Matušinsky, P. Transcriptional Analysis of Wheat Seedlings Inoculated with Fusarium Culmorum under Continual Exposure to Disease Defence Inductors. PLoS ONE 2020, 15, e0224413. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, Z.; Ling, N.; Yuan, Y.; Zheng, X.; Shen, B.; Shen, Q. Bacillus Subtilis SQR 9 Can Control Fusarium Wilt in Cucumber by Colonizing Plant Roots. Biol. Fertil. Soils 2011, 12, 495–506. [Google Scholar] [CrossRef]
- Lemańczyk, G.; Kwaśna, H. Effects of Sharp Eyespot (Rhizoctonia cerealis) on Yield and Grain Quality of Winter Wheat. Eur. J. Plant Pathol. 2013, 135, 187–200. [Google Scholar] [CrossRef] [Green Version]
- Rothen, C.; Miranda, V.; Fracchia, S.; Godeas, A.; Rodríguez, A. Microdochium bolleyi (Ascomycota: Xylariales): Physiological Characterization and Structural Features of Its Association with Wheat. Bol. Soc. Argent. Bot. 2018, 53, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Perotto, S.; Rodda, M.; Benetti, A.; Sillo, F.; Ercole, E.; Rodda, M.; Girlanda, M.; Murat, C.; Balestrini, R. Gene Expression in Mycorrhizal Orchid Protocorms Suggests a Friendly Plant–Fungus Relationship. Planta 2014, 239, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Adholeya, A.; Tiwari, P.; Singh, R. Large-Scale Inoculum Production of Arbuscular Mycorrhizal Fungi on Root Organs and Inoculation Strategies. In In Vitro Culture of Mycorrhizas; Declerck, S., Fortin, J.A., Strullu, D.-G., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2005; Volume 4, pp. 315–338. ISBN 978-3-540-24027-3. [Google Scholar]
- Dehne, H.-W.; Backhaus, G.F. The Use of Vesicular-Arbuscular Mycorrhizal Fungi in Plant Production. I. Inoculum Production/Zur Nutzung Vesikulär-Arbuskulärer Mykorrhizapilze in Der Pflanzenproduktion I. Inokulumgewinnung. Z. Pflanz. Pflanzenschutz J. Plant Dis. Prot. 1986, 93, 415–424. [Google Scholar]
- Dsouza, J. Techniques for the mass production of Arbuscular Mycorrhizal fungal species. In Advances in Biological Science Research; Elsevier: Amsterdam, The Netherlands, 2019; pp. 445–451. ISBN 978-0-12-817497-5. [Google Scholar]
- IJdo, M.; Cranenbrouck, S.; Declerck, S. Methods for Large-Scale Production of AM Fungi: Past, Present, and Future. Mycorrhiza 2011, 21, 1–16. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of Plant Growth-Promoting Bacteria for Biocontrol of Plant Diseases: Principles, Mechanisms of Action, and Future Prospects. AEM 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [Green Version]
- Ownley, B.; Weller, D.; Thomashow, L. Influence of in Situ and in Vitro PH on Suppression of Gaeumannomyces graminis Var. Tritici by Pseudomonas fluorescens 2-79. Phytopathology 1992, 82, 178–184. [Google Scholar] [CrossRef]
- Nicholson, P.; Parry, D.W. Development and Use of a PCR Assay to Detect Rhizoctonia Cerealis, the Cause of Sharp Eyespot in Wheat. Plant Pathol. 1996, 45, 872–883. [Google Scholar] [CrossRef]
- Zhu, X.; Qi, L.; Liu, X.; Cai, S.; Xu, H.; Huang, R.; Li, J.; Wei, X.; Zhang, Z. The Wheat Ethylene Response Factor Transcription Factor PATHOGEN-INDUCED ERF1 Mediates Host Responses to Both the Necrotrophic Pathogen Rhizoctonia cerealis and Freezing Stresses. Plant Physiol. 2014, 164, 1499–1514. [Google Scholar] [CrossRef] [Green Version]
- Neate, S.M.; Cruickshank, R.H. Pectic Enzyme Patterns of Ceratobasidium and Rhizoctonia Spp. Associated with Sharp Eyespot-like Lesions on Cereals in South Australia. Trans. Br. Mycol. Soc. 1988, 91, 267–272. [Google Scholar] [CrossRef]
- Sticher, L.; Mauch-Mani, B. Systemic acquired resistance. Annu. Rev. Phytopathol. 1997, 35, 235–270. [Google Scholar] [CrossRef] [PubMed]
- Adamo, M.; Chialva, M.; Calevo, J.; De Rose, S.; Girlanda, M.; Perotto, S.; Balestrini, R. The Dark Side of Orchid Symbiosis: Can Tulasnella Calospora Decompose Host Tissues? IJMS 2020, 21, 3139. [Google Scholar] [CrossRef] [PubMed]
- Blanco, R.; Aveling, T.A.S. Seed-Borne Fusarium Pathogens in Agricultural Crops. In Acta Horticulturae; International Society for Horticultural Science (ISHS): Leuven, Belgium, 2018; pp. 161–170. [Google Scholar]
- Leišová, L.; Kučera, L.; Chrpová, J.; Sýkorová, S.; Šíp, V.; Ovesná, J. Quantification of Fusarium Culmorum in Wheat and Barley Tissues Using Real-Time PCR in Comparison with DON Content. J. Phytopathol. 2006, 154, 603–611. [Google Scholar] [CrossRef]
- Bai, G.; Shaner, G. Management and resistance in wheat and barley to fusarium head blight. Annu. Rev. Phytopathol. 2004, 42, 135–161. [Google Scholar] [CrossRef] [PubMed]
- Clement, J.A.; Parry, D.W. Stem-Base Disease and Fungal Colonisation of Winter Wheat Grown in Compost Inoculated with Fusarium Culmorum, F. Graminearum and Microdochium Nivale. Eur. J. Plant Pathol. 1998, 104, 323–330. [Google Scholar] [CrossRef]
- Beccari, G.; Covarelli, L.; Nicholson, P. Infection Processes and Soft Wheat Response to Root Rot and Crown Rot Caused by Fusarium Culmorum: Root Rot and Crown Rot Caused by F. culmorum on Soft Wheat. Plant Pathol. 2011, 60, 671–684. [Google Scholar] [CrossRef]
- Schroeder, K.L.; Paulitz, T.C. Root Diseases of Wheat and Barley during the Transition from Conventional Tillage to Direct Seeding. Plant Dis. 2006, 90, 1247–1253. [Google Scholar] [CrossRef] [Green Version]
- Smiley, R.W.; Gourlie, J.A.; Easley, S.A.; Patterson, L.-M. Pathogenicity of Fungi Associated with the Wheat Crown Rot Complex in Oregon and Washington. Plant Dis. 2005, 89, 949–957. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Ellwood, S.R.; Oliver, R.P.; Friesen, T.L. Pyrenophora Teres: Profile of an Increasingly Damaging Barley Pathogen: Pathogen Profile of Pyrenophora Teres. Mol. Plant Pathol. 2011, 12, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Murray, G.M.; Brennan, J.P. Estimating Disease Losses to the Australian Wheat Industry. Austral. Plant Pathol. 2009, 38, 558. [Google Scholar] [CrossRef]
- Leišová-Svobodová, L.; Minaříková, V.; Matušinsky, P.; Hudcovicová, M.; Ondreičková, K.; Gubiš, J. Genetic Structure of Pyrenophora Teres Net and Spot Populations as Revealed by Microsatellite Analysis. Fungal Biol. 2014, 118, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Matusinsky, P.; Frei, P.; Mikolasova, R.; Svacinova, I.; Tvaruzek, L.; Spitzer, T. Species-Specific Detection of Bipolaris Sorokiniana from Wheat and Barley Tissues. Crop Prot. 2010, 29, 1325–1330. [Google Scholar] [CrossRef]
- Anderson, R.C.; Liberta, A.E.; Dickman, L.A. Interaction of Vascular Plants and Vesicular-Arbuscular Mycorrhizal Fungi across a Soil Moisture-Nutrient Gradient. Oecologia 1984, 64, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular Mycorrhizal Fungi in Alleviation of Salt Stress: A Review. Ann. Bot. 2009, 104, 1263–1280. [Google Scholar] [CrossRef] [Green Version]
- Malinowski, D.P.; Belesky, D.P. Adaptations of Endophyte-Infected Cool-Season Grasses to Environmental Stresses: Mechanisms of Drought and Mineral Stress Tolerance. Crop Sci. 2000, 40, 923–940. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. 38—Amplification and direct sequencing of fungal ribosomal rna genes for phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Matusinsky, P.; Zouhar, M.; Pavela, R.; Novy, P. Antifungal Effect of Five Essential Oils against Important Pathogenic Fungi of Cereals. Ind. Crops Prod. 2015, 67, 208–215. [Google Scholar] [CrossRef]
- Koske, R.E.; Gemma, J.N. A Modified Procedure for Staining Roots to Detect VA Mycorrhizas. Mycol. Res. 1989, 92, 486–488. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Inhibition Zone with Fusarium culmorum | Reduction of Fusarium culmorum Growth | Causing Degradation of Pyrenophora teres |
|---|---|---|---|
| Ceratobasidium 2015/1 | - | - | + |
| Tulasnella 2015/2 | - | - | - |
| Tulasnella 2016/2 | - | - | - |
| Tulasnella 2016/7 | - | - | - |
| Tulasnella 2016/11 | - | - | + |
| Microdochium bolleyi | + | - | - |
| Ceratobasidium cereale | - | + | + |
| Treatment | Number of Plants | Number of Vital Plants |
|---|---|---|
| Ceratobasidium 2015/1 | 45.7 ± 2.3 b | 24.0 ± 7.0 bc |
| Tulasnella 2015/2 | 43.7 ± 4.9 b | 27.7 ± 7.2 b |
| Tulasnella 2016/2 | 40.3 ± 5.5 bc | 26.0 ± 2.6 bc |
| Tulasnella 2016/7 | 41.7 ± 2.1 b | 32.3 ± 5.1 b |
| Tulasnella 2016/11 | 32.0 ± 6.1 c | 17.0 ± 4.4 c |
| Kinto Duo | 89.3 ± 2.1 a | 89.0 ± 2.0 a |
| Control (with Fusarium) | 47.7 ± 10.1 b | 29.0 ± 7.9 b |
| Treatment | Experiment 1 | Experiment 2 | Experiment 3 | Field |
|---|---|---|---|---|
| Ceratobasidium 2015/1 | 3.3 ± 2.9 b | 13.3 ± 5.5 de | 25.7 ± 10.3 c | 19.3 ± 6.7 cd |
| Tulasnella 2015/2 | 3.3 ± 2.9 b | 30.0 ± 11.5 bc | 79.7 ± 12.9 a | 5.7 ± 1.5 ab |
| Tulasnella 2016/2 | 12.3 ± 4.9 a | 41.0 ± 7.0 b | 74.0 ± 8.0 a | 16.7 ± 10.1 cd |
| Tulasnella 2016/7 | 8.3 ± 3.5 ab | 81.3 ± 6.7 a | 87.0 ± 3.6 a | 2.7 ± 1.2 a |
| Tulasnella 2016/11 | 12.0 ± 4.6 a | 83.3 ± 4.2 a | 87.7 ± 3.8 a | 29.3 ± 16.4 d |
| Ceratobasidium cereale | 5.3 ± 2.5 e | 55.7 ± 7.1 b | ||
| Microdochium bolleyi | 27.7 ± 12.9 c | 75.0 ± 9.8 a | ||
| Serendipita indica | 55.0 ± 6.6 b | 11.0 ± 2.6 bc | ||
| Kinto Duo | ND | ND | ND | ND |
| Control (with Fusarium) | 4.3 ± 4.0 b | 18.7 ± 3.5 cd | 51.0 ± 14.0 b | |
| Control (without Fusarium) | ND | ND |
| Treatment | Number of Symptomless Plants | Number of Vital Plants | Dry Shoot Mass Per Plant (mg) |
|---|---|---|---|
| Ceratobasidium 2015/1 | 16.0 ± 5.3 ab | 64.3 ± 12.9 b | 16.4 ± 0.5 ab |
| Tulasnella 2015/2 | 5.7 ± 6.0 c | 54.0 ± 10.0 b | 16.3 ± 0.7 ab |
| Tulasnella 2016/2 | 12.3 ± 11.0 bc | 60.7 ± 15.5 b | 17.4 ± 1.1 a |
| Tulasnella 2016/7 | 7.3 ± 6.1 bc | 54.0 ± 10.5 b | 15.8 ± 0.6 bc |
| Tulasnella 2016/11 | 8.7 ± 5.5 bc | 60.7 ± 14.0 b | 15.9 ± 1.6 abc |
| Ceratobasidium cereale | 0.0 ± 0.0 d | 68.0 ± 1.0 b | 14.6 ± 0.9 cd |
| Microdochium bolleyi | 20.0 ± 5.3 ab | 70.0 ± 13.0 b | 16.1 ± 0.1 abc |
| Kinto Duo | 47.3 ± 22.7 a | 88.7 ± 3.1 a | 14.1 ± 0.7 d |
| Control (with Fusarium) | 17.0 ± 5.6 ab | 58.3 ± 5.1 b | 17.0 ± 1.2 ab |
| Treatment | Number of Plants | Number of Vital Plants | Dry Shoot Mass (g) | Dry Mass Per Plant (mg) |
|---|---|---|---|---|
| Ceratobasidium 2015/1 | 73.3 ± 16.1 e | 51.0 ± 12.3 e | 0.922 ± 0.22 cd | 13.3 ± 5.7 abc |
| Tulasnella 2015/2 | 89.0 ± 7.0 abcd | 56.0 ± 9.8 e | 1.188 ± 0.113 b | 13.4 ± 1.0 abc |
| Tulasnella 2016/2 | 92.0 ± 4.6 abc | 81.0 ± 8.5 bc | 1.198 ± 0.104 b | 12.7 ± 1.4 bc |
| Tulasnella 2016/7 | 75.7 ± 8.4 de | 60.3 ± 8.5 de | 0.955 ± 0.109 cd | 13.0 ± 0.8 bc |
| Tulasnella 2016/11 | 93.3 ± 1.5 abc | 75.3 ± 9.0 cd | 1.181 ± 0.121 b | 12.7 ± 1.5 bc |
| Ceratobasidium cereale | 56.7 ± 3.1 f | 5.0 ± 3.6 g | 0.428 ± 0.141 e | 7.5 ± 1.5 d |
| Microdochium bolleyi | 85.3 ± 9.3 bcde | 60.7 ± 8.1 de | 0.998 ± 0.121 bcd | 11.6 ± 0.8 bc |
| Serendipita indica | 99.3 ± 0.6 a | 97.3 ± 2.3 a | 1.512 ± 0.067 a | 15.2 ± 0.6 ab |
| Kinto Duo | 90.7 ± 8.7 abc | 94.7 ± 4.0 ab | 1.108 ± 0.032 bc | 11.7 ± 1.4 bc |
| Control (with Fusarium) | 81.3 ± 10.0 cde | 30.3 ± 16.8 f | 0.831 ± 0.177 d | 10.2 ± 2.1 cd |
| Control (without Fusarium) | 95.3 ± 3.8 ab | 89.3 ± 8.1 abc | 1.504 ± 0.089 a | 16.6 ± 1.1 a |
| Isolate Origin | Internal Code | Species | Accession Number GenBank |
|---|---|---|---|
| Ophrys bombyliflora | 2015/1 | Ceratobasidium sp. | MW488152 |
| Orchis italica | 2015/2 | Tulasnella sp. | MW485781 |
| ×Serapicamptis capitata | 2016/2 | Tulasnella sp. | MW485782 |
| Serapias lingua | 2016/7 | Tulasnella sp. | MW485784 |
| Serapias lingua | 2016/11 | Tulasnella sp. | MW485827 |
| Triticum aestivum | 18–301 | Microdochium bolleyi | MW485763 |
| Triticum aestivum | 18–300 | Ceretobasidium cereale | MW485776 |
| Prosopis juliflora rhizosphere [11] | CBS 125645 | Serendipita indica | MH863568 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bleša, D.; Matušinský, P.; Sedmíková, R.; Baláž, M. The Potential of Rhizoctonia-Like Fungi for the Biological Protection of Cereals against Fungal Pathogens. Plants 2021, 10, 349. https://doi.org/10.3390/plants10020349
Bleša D, Matušinský P, Sedmíková R, Baláž M. The Potential of Rhizoctonia-Like Fungi for the Biological Protection of Cereals against Fungal Pathogens. Plants. 2021; 10(2):349. https://doi.org/10.3390/plants10020349
Chicago/Turabian StyleBleša, Dominik, Pavel Matušinský, Romana Sedmíková, and Milan Baláž. 2021. "The Potential of Rhizoctonia-Like Fungi for the Biological Protection of Cereals against Fungal Pathogens" Plants 10, no. 2: 349. https://doi.org/10.3390/plants10020349
APA StyleBleša, D., Matušinský, P., Sedmíková, R., & Baláž, M. (2021). The Potential of Rhizoctonia-Like Fungi for the Biological Protection of Cereals against Fungal Pathogens. Plants, 10(2), 349. https://doi.org/10.3390/plants10020349