Nitrogen Fertilization Modified the Responses of Schima superba Seedlings to Elevated CO2 in Subtropical China

Abstract
:1. Introduction
2. Results
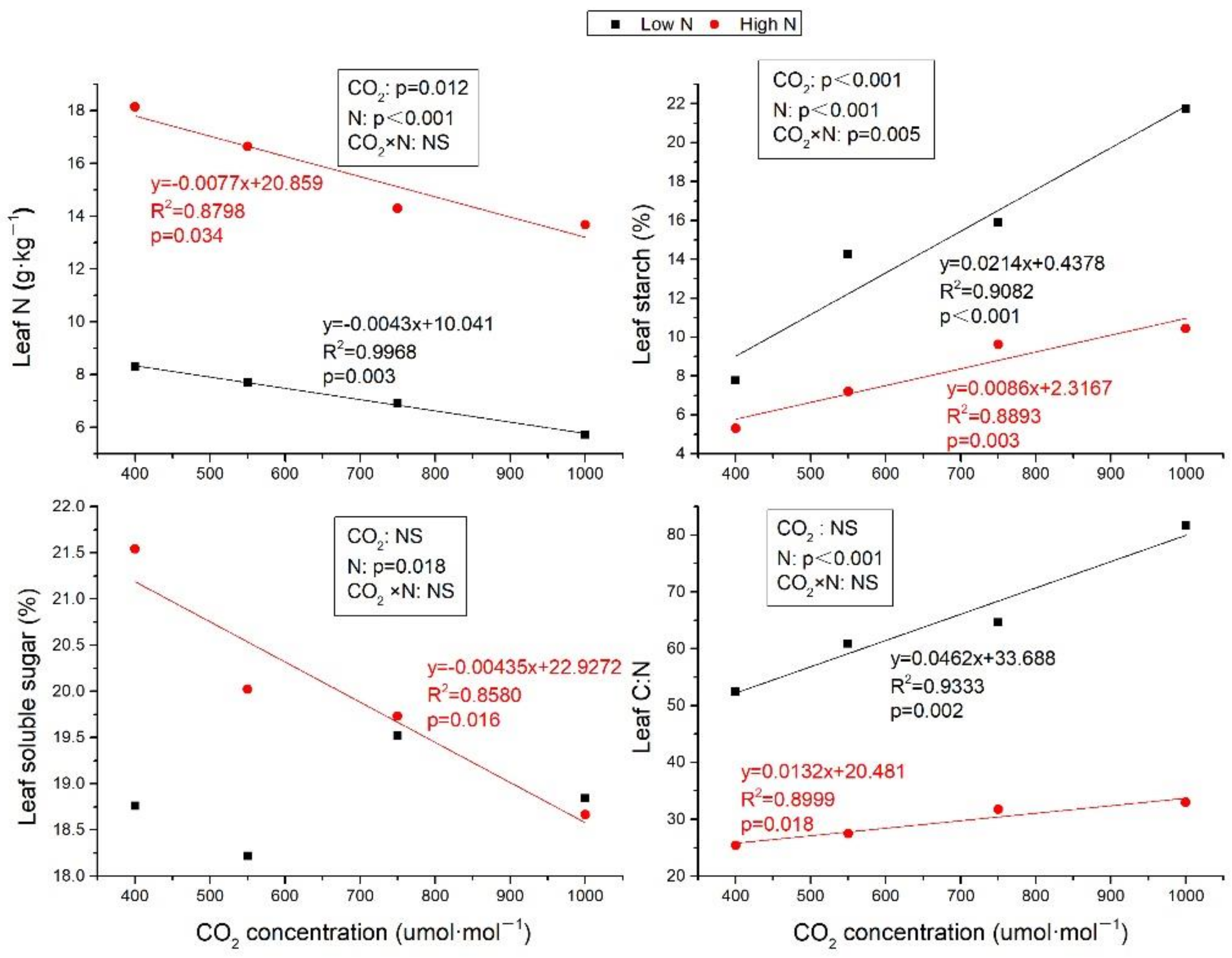
2.1. Leaf N, C:N Ratio, and Carbohydrates
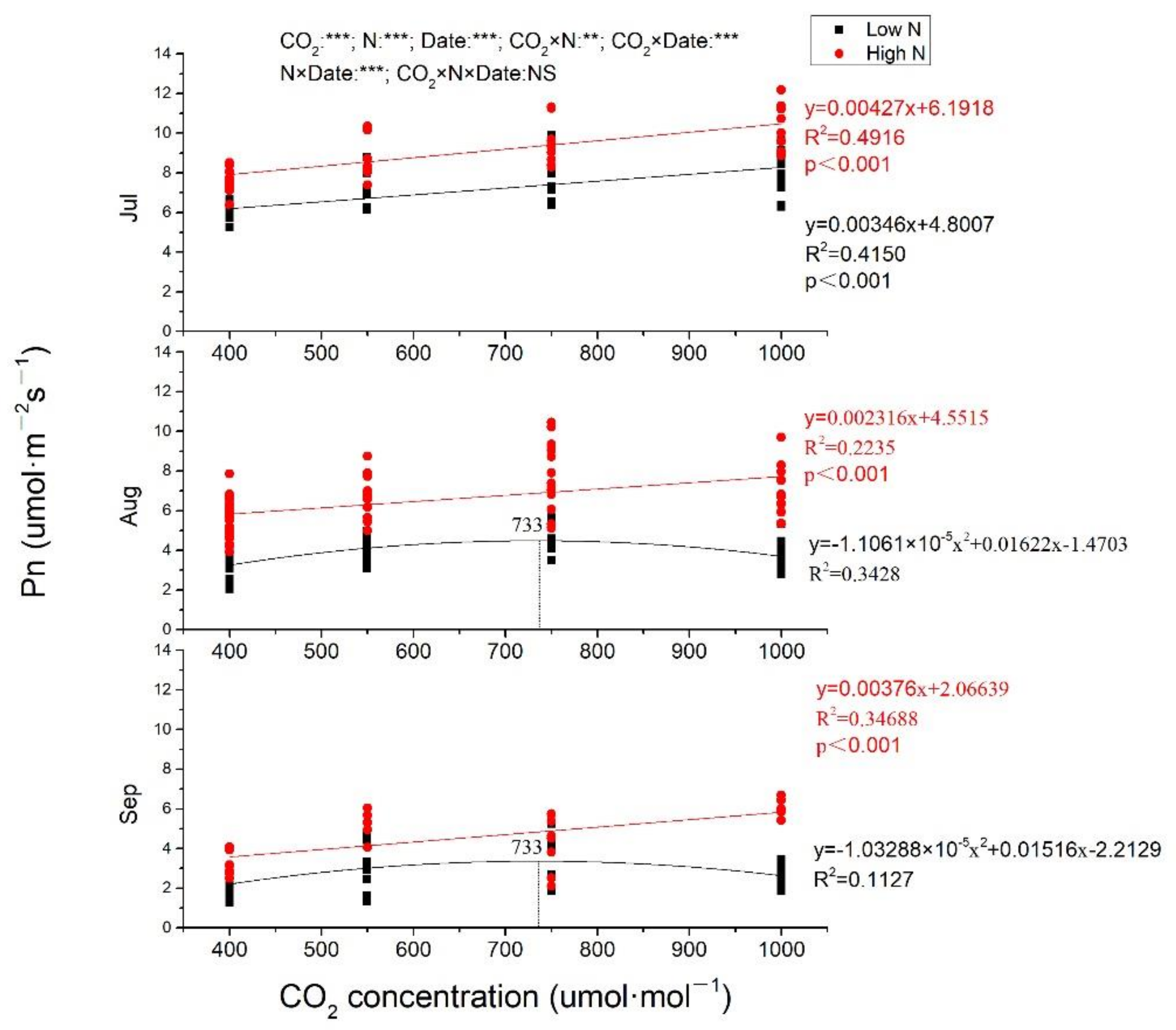
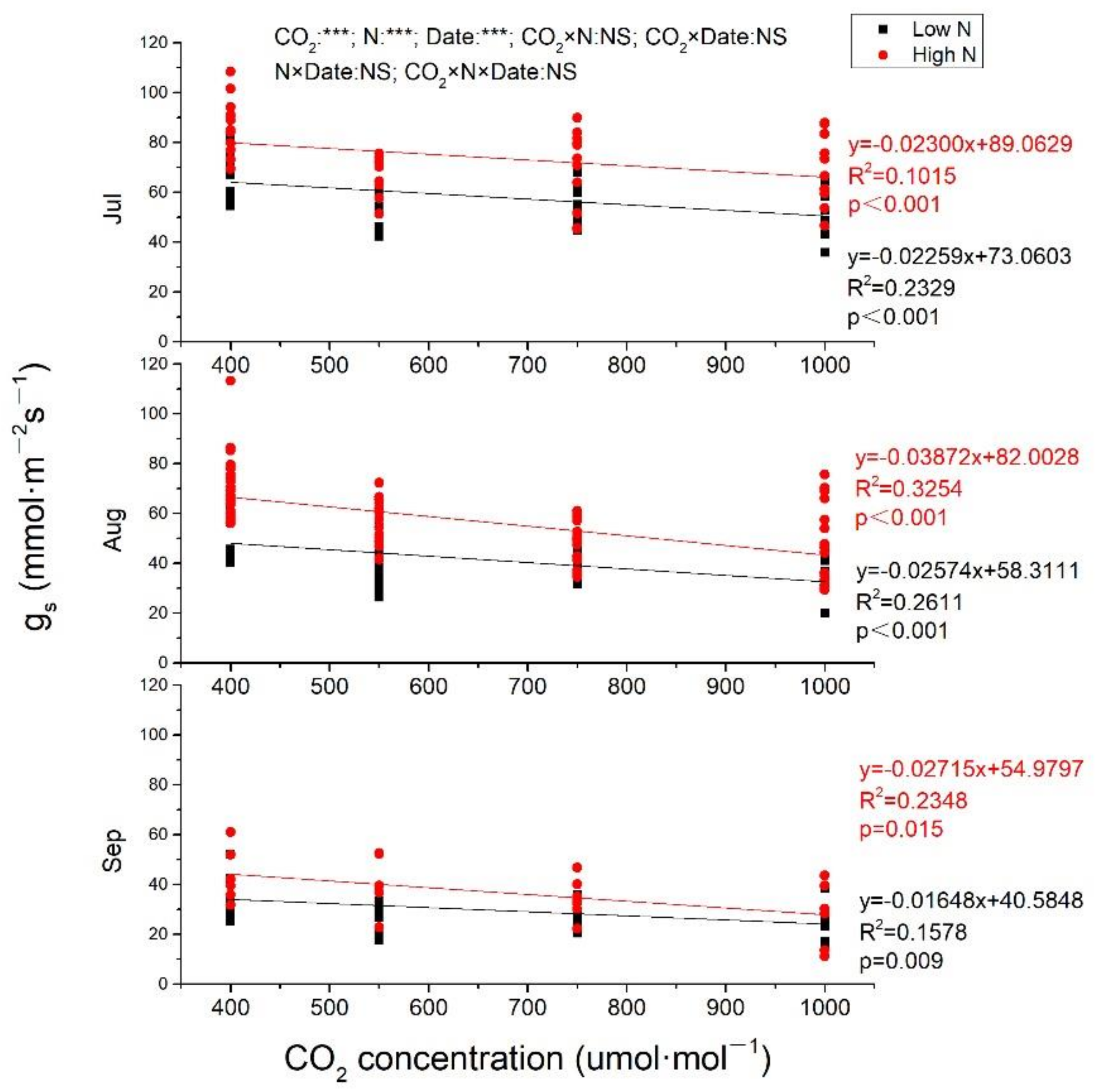
2.2. Gas Exchange
2.3. Plant Biomass
3. Discussion
4. Materials and Methods
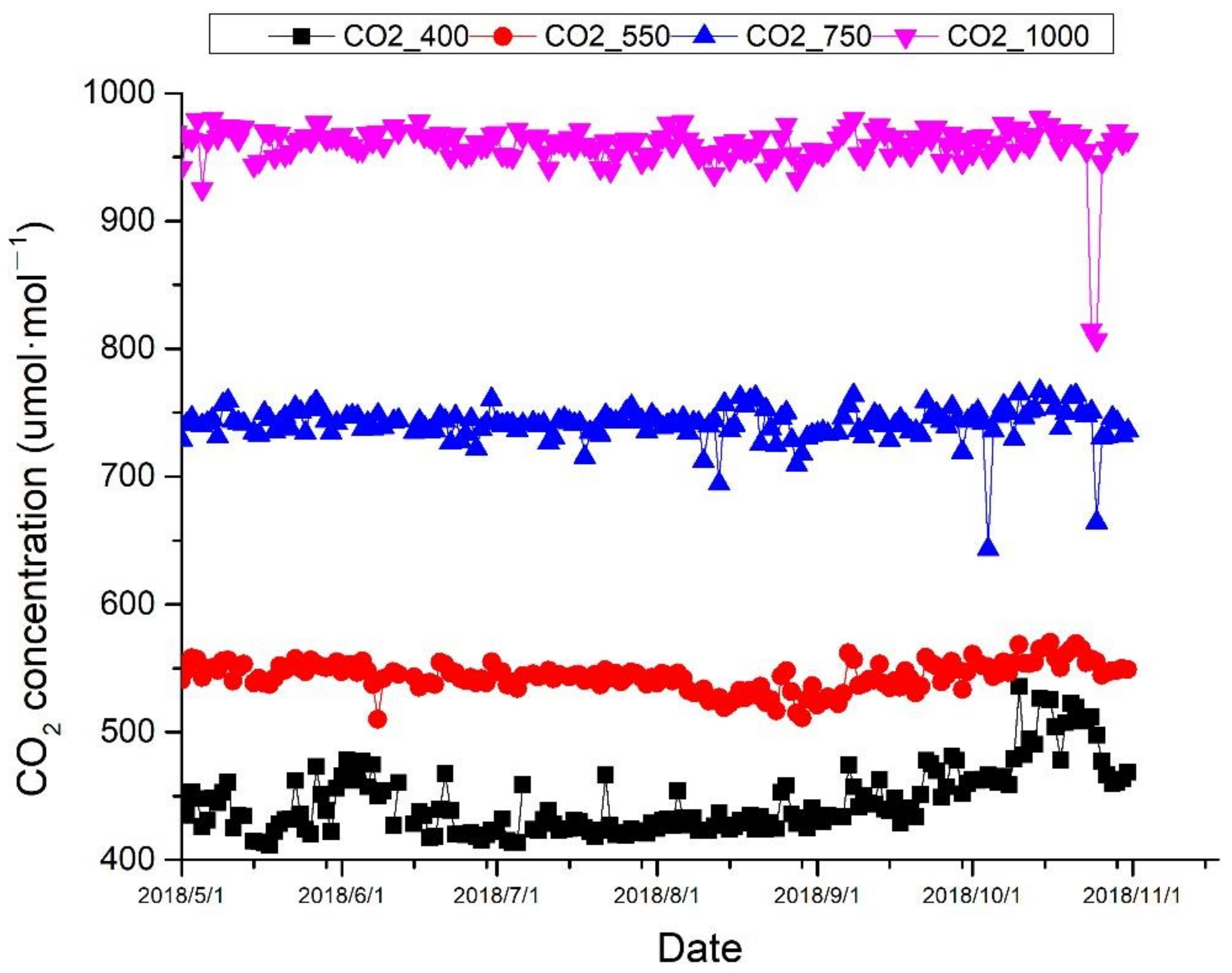
4.1. Experimental Site and Design
4.2. Plant Growth
4.3. Sampling, Physiological and Biochemical Measurements
4.3.1. Plant C, N, and Carbohydrates
4.3.2. Gas Exchange Measurement
4.4. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maroco, J.P.; Breia, E.; Faria, T.; Pereira, J.S.; Chaves, M.M. Effects of long-term exposure to elevated CO2 and n fertilization on the development of photosynthetic capacity and biomass accumulation in Quercus suber L. Plant Cell Environ. 2002, 25, 105–113. [Google Scholar] [CrossRef]
- De Kauwe, M.G.; Medlyn, B.E.; Zaehle, S.; Walker, A.P.; Dietze, M.C.; Hickler, T.; Jain, A.K.; Luo, Y.; Parton, W.J.; Prentice, I.C.; et al. Forest water use and water use efficiency at elevated CO2: A model-data intercomparison at two contrasting temperate forest FACE sites. Glob. Chang. Biol. 2013, 19, 1759–1779. [Google Scholar] [CrossRef] [PubMed]
- Newingham, B.A.; Vanier, C.H.; Charlet, T.N.; Ogle, K.; Smith, S.D.; Nowak, R.S. No cumulative effect of 10 years of elevated [CO2] on perennial plant biomass components in the Mojave Desert. Glob. Chang. Biol. 2013, 19, 2168–2181. [Google Scholar] [CrossRef]
- Taylor, B.N.; Strand, A.E.; Cooper, E.R.; Beidler, K.V.; Schönholz, M.; Pritchard, S.G. Root length, biomass, tissue chemistry and mycorrhizal colonization following 14 years of CO2 enrichment and 6 years of N fertilization in a warm temperate forest. Tree Physiol. 2014, 34, 955–965. [Google Scholar] [CrossRef] [Green Version]
- Terrer, C.; Vicca, S.; Stocker, B.D.; Hungate, B.A.; Phillips, R.P.; Reich, P.B.; Finzi, A.C.; Prentice, I.C. Ecosystem responses to elevated CO2 governed by plant–soil interactions and the cost of nitrogen acquisition. New Phytol. 2018, 217, 507–522. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Xie, H.; Wu, S.; Wang, Z.; He, K. Effects of elevated CO2 and increased N fertilization on plant secondary metabolites and chewing insect fitness. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; 151p. [Google Scholar]
- Wang, R.; Goll, D.; Balkanski, Y.; Hauglustaine, D.; Boucher, O.; Ciais, P.; Janssens, I.; Penuelas, J.; Guenet, B.; Sardans, J.; et al. Global forest carbon uptake due to nitrogen and phosphorus deposition from 1850 to 2100. Glob. Chang. Biol. 2017, 23, 4854–4872. [Google Scholar] [CrossRef]
- Kanakidou, M.; Myriokefalitakis, S.; Daskalakis, N.; Fanourgakis, G.; Nenes, A.; Baker, A.R.; Tsigaridis, K.; Mihalopoulos, N. Past, present, and future atmospheric nitrogen deposition. J. Atmos. Sci. 2016, 73, 2039–2047. [Google Scholar] [CrossRef] [Green Version]
- Decina, S.M.; Hutyra, L.R.; Templer, P.H. Hotspots of nitrogen deposition in the world’s urban areas: A global data synthesis. Front. Ecol. Environ. 2020, 18, 92–100. [Google Scholar] [CrossRef] [Green Version]
- Aranjuelo, I.; Cabrerizo, P.M.; Arrese-Igor, C.; Aparicio-Tejo, P.M. Pea plant responsiveness under elevated [CO2] is conditioned by the N source (N2 fixation versus NO3− fertilization). Environ. Exp. Bot. 2013, 95, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Lambreva, M.; Christov, K.; Tsonev, T. Short-term effect of elevated CO2 concentration and high irradiance on the antioxidant enzymes in bean plants. Biol. Plant. 2006, 50, 617–623. [Google Scholar] [CrossRef]
- Ryan, G.D.; Rasmussen, S.; Xue, H.; Parsons, A.J.; Newman, J.A. Metabolite analysis of the effects of elevated CO2 and nitrogen fertilization on the association between tall fescue (Schedonorus arundinaceus) and its fungal symbiont Neotyphodium coenophialum. Plant Cell Environ. 2014, 37, 204–212. [Google Scholar] [CrossRef]
- Yoder, C.K.; Vivin, P.; Defalco, L.A.; Seemann, J.R.; Nowak, R.S. Root growth and function of three Mojave Desert grasses in response to elevated atmospheric CO2 concentration. New Phytol. 2000, 145, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Polley, H.W.; Johnson, H.B.; Mayeux, H.S. Leaf physiology, production, water use, and nitrogen dynamics of the grassland invader Acacia smallii at elevated CO2 concentrations. Tree Physiol. 1997, 17, 89–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, E.P.; Erickson, J.E.; Kruger, E.L. Can decreased transpiration limit plant nitrogen acquisition in elevated CO2? Funct. Plant Biol. 2002, 29, 1115–1120. [Google Scholar] [CrossRef]
- Chang, Z.; Ma, Y.; Zhou, C. Effects of elevated CO2 concentration on water use efficiency of Tamarix ramosissima in an extremely arid region. Environ. Earth Sci. 2016, 75, 1409.1–1409.6. [Google Scholar] [CrossRef]
- Deng, Q.; Zhou, G.; Liu, J.; Liu, S.; Duan, H.; Zhang, D. Responses of soil respiration to elevated carbon dioxide and nitrogen addition in young subtropical forest ecosystems in China. Biogeosciences 2010, 7, 315–328. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Huang, W.; Zhou, G.; Zhang, D.; Liu, S.; Li, Y. Nitrogen to phosphorus ratios of tree species in response to elevated carbon dioxide and nitrogen addition in subtropical forests. Glob. Chang. Biol. 2013, 19, 208–216. [Google Scholar] [CrossRef]
- Sheu, B.H.; Lin, C.K. Photosynthetic response of seedlings of the sub-tropical tree Schima superba with exposure to elevated carbon dioxide and temperature. Environ. Exp. Bot. 1999, 41, 57–65. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, G.; Xu, Z.; Duan, H.; Li, Y.; Zhang, D. Photosynthesis acclimation, leaf nitrogen concentration, and growth of four tree species over 3 years in response to elevated carbon dioxide and nitrogen treatment in subtropical China. J. Soils Sediments 2011, 11, 1155–1164. [Google Scholar] [CrossRef]
- Kant, S.; Seneweera, S.; Rodin, J.; Materne, M.; Burch, D.; Rothstein, S.J.; Spangenberg, G. Improving yield potential in crops under elevated CO2: Integrating the photosynthetic and nitrogen utilization efficiencies. Front. Plant Sci. 2012, 3, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotfiomran, N.; Köhl, M.; Fromm, J. Interaction effect between elevated CO2 and fertilization on biomass, gas exchange and C/N ratio of European beech (Fagus sylvatica L.). Plants 2016, 5, 38. [Google Scholar] [CrossRef] [Green Version]
- Taub, D.R.; Wang, X. Why are nitrogen concentrations in plant tissues lower under elevated CO2? A critical examination of the hypotheses. J. Integr. Plant Biol. 2008, 50, 1365–1374. [Google Scholar] [CrossRef]
- Bloom, A.J.; Burger, M.; Kimball, B.A.; Pinter, J.P., Jr. Nitrate assimilation is inhibited by elevated CO2 in field-grown wheat. Nat. Clim. Chang. 2014, 4, 477–480. [Google Scholar] [CrossRef]
- Li, L.; Manning, W.; Wang, X. Elevated CO2 increases root mass and leaf nitrogen resorption in red maple (Acer rubrum L.). Forests 2019, 10, 420. [Google Scholar] [CrossRef] [Green Version]
- Calfapietra, C.; Tulva, I.; Eensalu, E.; Perez, M.; De Angelis, P.; Scarascia-Mugnozza, G.; Kull, O. Canopy profiles of photosynthetic parameters under elevated CO2 and N fertilization in a poplar plantation. Environ. Pollut. 2005, 137, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, D.; Zhou, G.; Duan, H. Changes in leaf nutrient traits and photosynthesis of four tree species: Effects of elevated [CO2], N fertilization and canopy positions. J. Plant Ecol. 2012, 5, 376–390. [Google Scholar] [CrossRef] [Green Version]
- Kou, L.; Guo, D.; Yang, H.; Gao, W.; Li, S. Growth, morphological traits and mycorrhizal colonization of fine roots respond differently to nitrogen addition in a slash pine plantation in subtropical China. Plant Soil 2015, 391, 207–218. [Google Scholar] [CrossRef]
- Wingler, A.; Purdy, S.; MacLean, J.A.; Pourtau, N. The role of sugars in integrating environmental signals during the regulation of leaf senescence. J. Exp. Bot. 2006, 57, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A.R.; Rasineni, G.K.; Raghavendra, A.S. The impact of global elevated CO2 concentration on photosynthesis and plant productivity. Curr. Sci. 2010, 99, 46–57. [Google Scholar]
- Kimball, B.A. Crop responses to elevated CO2 and interactions with H2O, N, and temperature. Curr. Opin. Plant Biol. 2016, 31, 36–43. [Google Scholar] [CrossRef]
- Vizoso, S.; Gerant, D.; Guehl, J.M.; Joffre, R.; Chalot, M.; Gross, P.; Maillard, P. Do elevation of CO2 concentration and nitrogen fertilization alter storage and remobilization of carbon and nitrogen in pedunculate oak saplings? Tree Physiol. 2008, 28, 1729–1739. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Davey, P.A.; Hymus, G.J.; Osborne, C.P.; Rogers, A.; Blum, H.; Nösberger, J.; Long, S.P. Is stimulation of leaf photosynthesis by elevated carbon dioxide concentration maintained in the long term? A test with Lolium perenne grown for 10 years at two nitrogen fertilization levels under Free Air CO2 Enrichment (FACE). Plant Cell Environ. 2003, 26, 705–714. [Google Scholar] [CrossRef] [Green Version]
- Duval, B.D.; Blankinship, J.C.; Dijkstra, P.; Hungate, B.A. CO2 effects on plant nutrient concentration depend on plant functional group and available nitrogen: A meta-analysis. Plant Ecol. 2012, 213, 505–521. [Google Scholar] [CrossRef]
- Duan, H.L.; Liu, J.X.; Deng, Q.; Chen, X.M.; Zhang, D.Q. Effects of elevated CO2 and N deposition on plant biomass accumulation and allocation in subtropical forest ecosystems: A mesocosm study. Chin. J. Plant Ecol. 2009, 33, 570–579. [Google Scholar]
- Chen, Z.; Maltz, M.R.; Cao, J.; Yu, H.; Shang, H.; Aronson, E. Elevated O3 alters soil bacterial and fungal communities and the dynamics of carbon and nitrogen. Sci. Total Environ. 2019, 677, 272–280. [Google Scholar] [CrossRef]
- Yu, H.; Cao, J.; Chen, Z.; Shang, H. Effects of elevated O3 on physiological and biochemical responses in three kinds of trees native to subtropical forest in China during non-growing period. Environ. Pollut. 2018, 234, 716–725. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z.L.; Wang, H.; Guo, C.; Bao, W. Rainfall pulse primarily drives litterfall respiration and its contribution to soil respiration in a young exotic pine plantation in subtropical China. Can. J. For. Res. 2012, 42, 657–666. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CO2 (μmol·mol−1) | Biomass (g dw) | ||||
|---|---|---|---|---|---|
| Root | Stem | Leaf | Total | ||
| ambient | Low N | 6.77 ± 0.89 | 5.37 ± 0.87 | 6.17 ± 1.08 | 18.30 ±2.73 |
| High N | 7.55 ± 0.66 (0.519) | 8.63 ± 1.52 (0.135) | 9.55 ± 1.52 (0.144) | 25.73 ±3.60 (0.176) | |
| 550 | Low N | 5.50 ± 1.05 | 5.18 ± 0.45 | 5.30 ± 1.15 | 15.98 ±2.60 |
| High N | 9.38 ± 1.75 (0.130) | 7.10 ± 0.40 (0.063) | 10.13 ± 0.52 (0.019) | 26.62 ± 2.83 (0.05) | |
| 750 | Low N | 5.42 ± 0.83 | 5.85 ± 0.81 | 5.53 ± 0.49 | 16.80 ± 1.35 |
| High N | 11.70 ± 1.70 (0.30) | 12.42 ± 2.65 (0.077) | 12.17 ± 2.44 (0.056) | 36.28 ± 6.51 (0.043) | |
| 1000 | Low N | 6.98 ± 1.02 | 5.95 ± 1.2 | 6.12 ± 0.91 | 19.05 ± 2.96 |
| High N | 13.60 ± 1.94 (0.039) | 12.45 ± 0.60 (0.008) | 13.27 ± 0.15 (0.014) | 39.32 ± 1.32 (0.003) | |
| CO2 | NS | NS | NS | NS | |
| N | <0.001 | <0.001 | <0.0001 | <0.0001 | |
| CO2 × N | NS | NS | NS | NS | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Ye, S.; Cao, J.; Shang, H. Nitrogen Fertilization Modified the Responses of Schima superba Seedlings to Elevated CO2 in Subtropical China. Plants 2021, 10, 383. https://doi.org/10.3390/plants10020383
Chen Z, Ye S, Cao J, Shang H. Nitrogen Fertilization Modified the Responses of Schima superba Seedlings to Elevated CO2 in Subtropical China. Plants. 2021; 10(2):383. https://doi.org/10.3390/plants10020383
Chicago/Turabian StyleChen, Zhan, Siyuan Ye, Jixin Cao, and He Shang. 2021. "Nitrogen Fertilization Modified the Responses of Schima superba Seedlings to Elevated CO2 in Subtropical China" Plants 10, no. 2: 383. https://doi.org/10.3390/plants10020383
APA StyleChen, Z., Ye, S., Cao, J., & Shang, H. (2021). Nitrogen Fertilization Modified the Responses of Schima superba Seedlings to Elevated CO2 in Subtropical China. Plants, 10(2), 383. https://doi.org/10.3390/plants10020383