
Identification and Detection of Botryosphaeria dothidea from Kiwifruit (Actinidia chinensis) in China


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
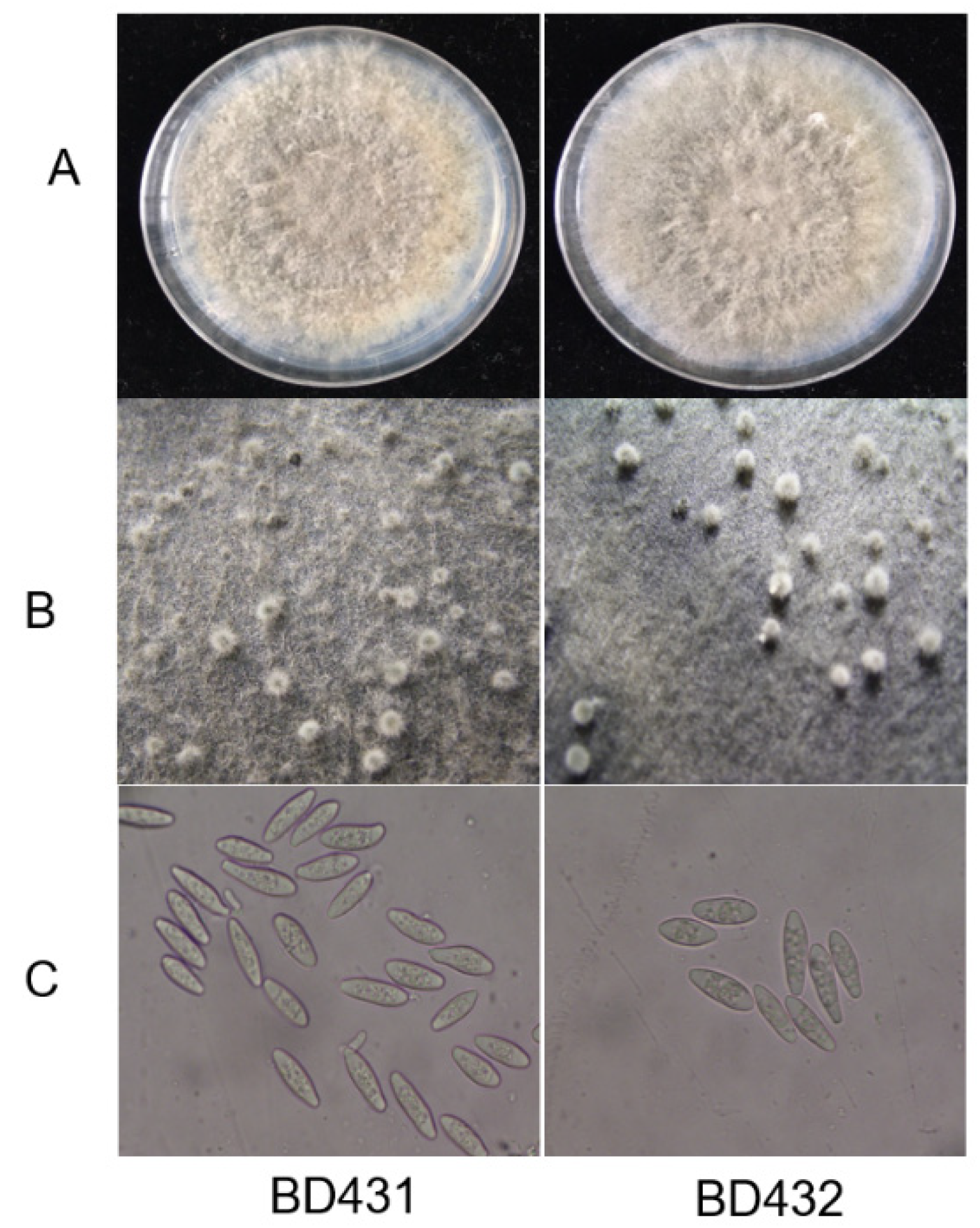
2.1. Isolate Characterization
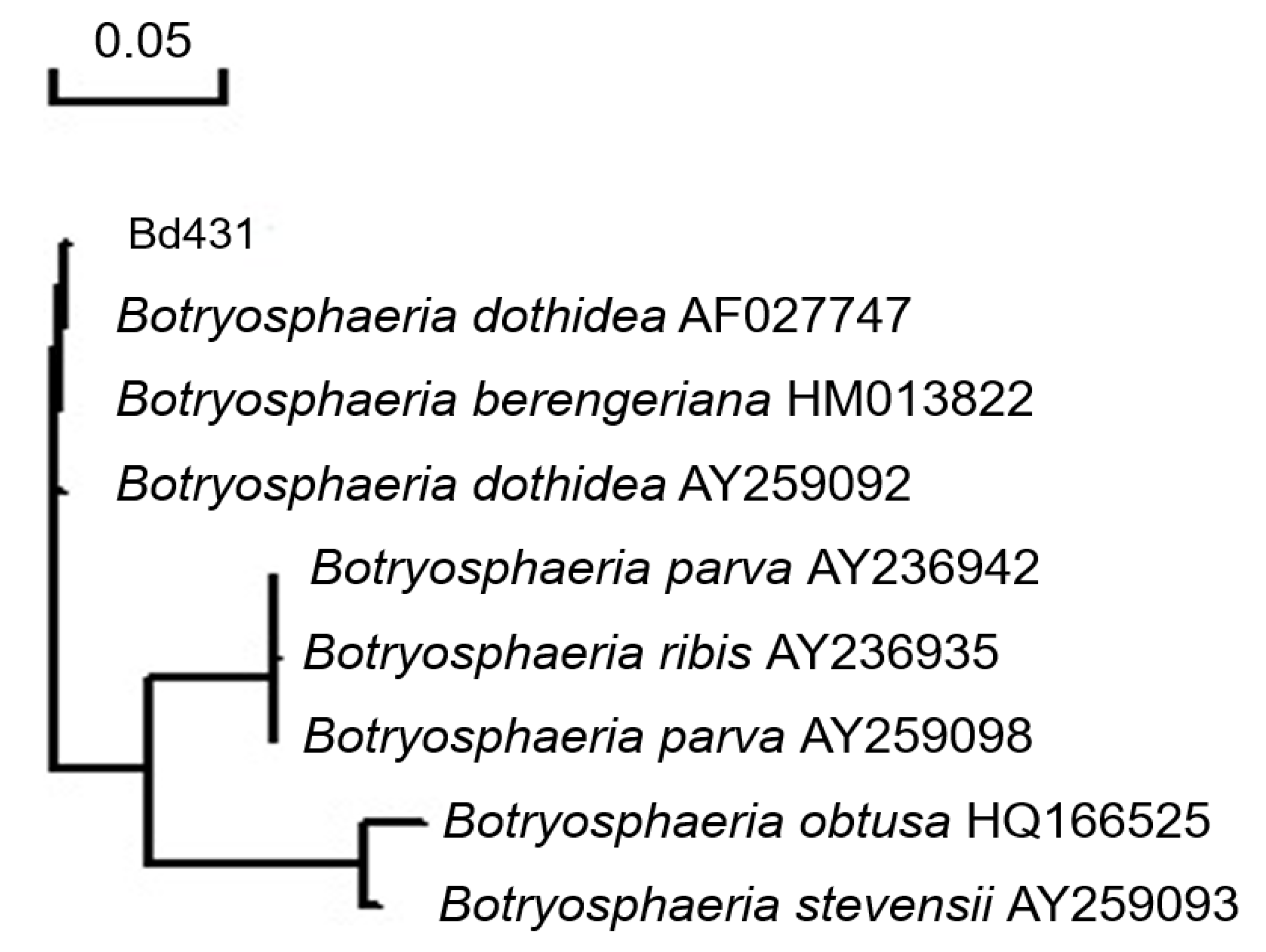
2.2. Molecular Identification
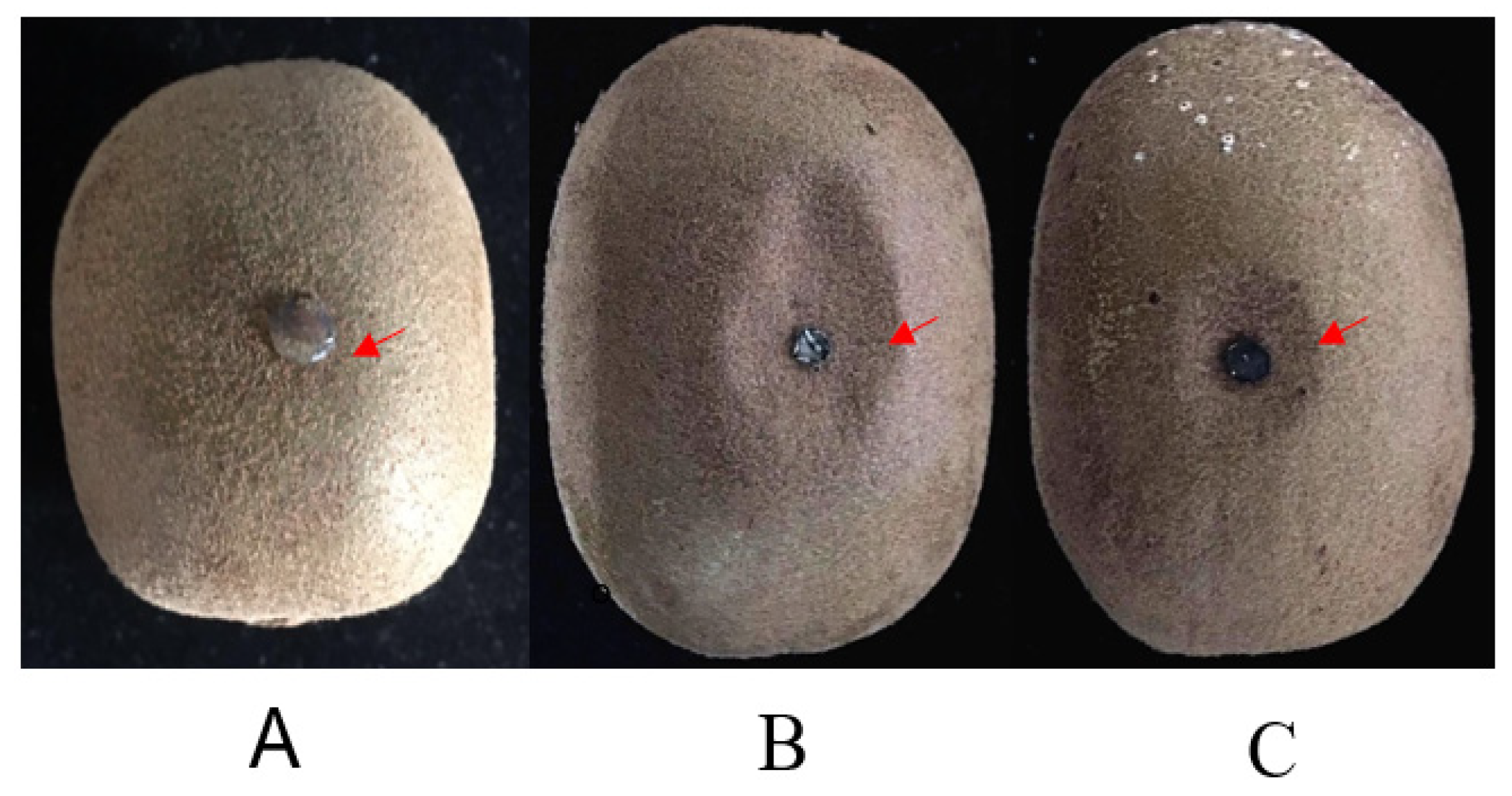
2.3. Pathogenicity Testing
2.4. Reisolation and Identification of Infected Fungi
2.5. LAMP Primers Selection
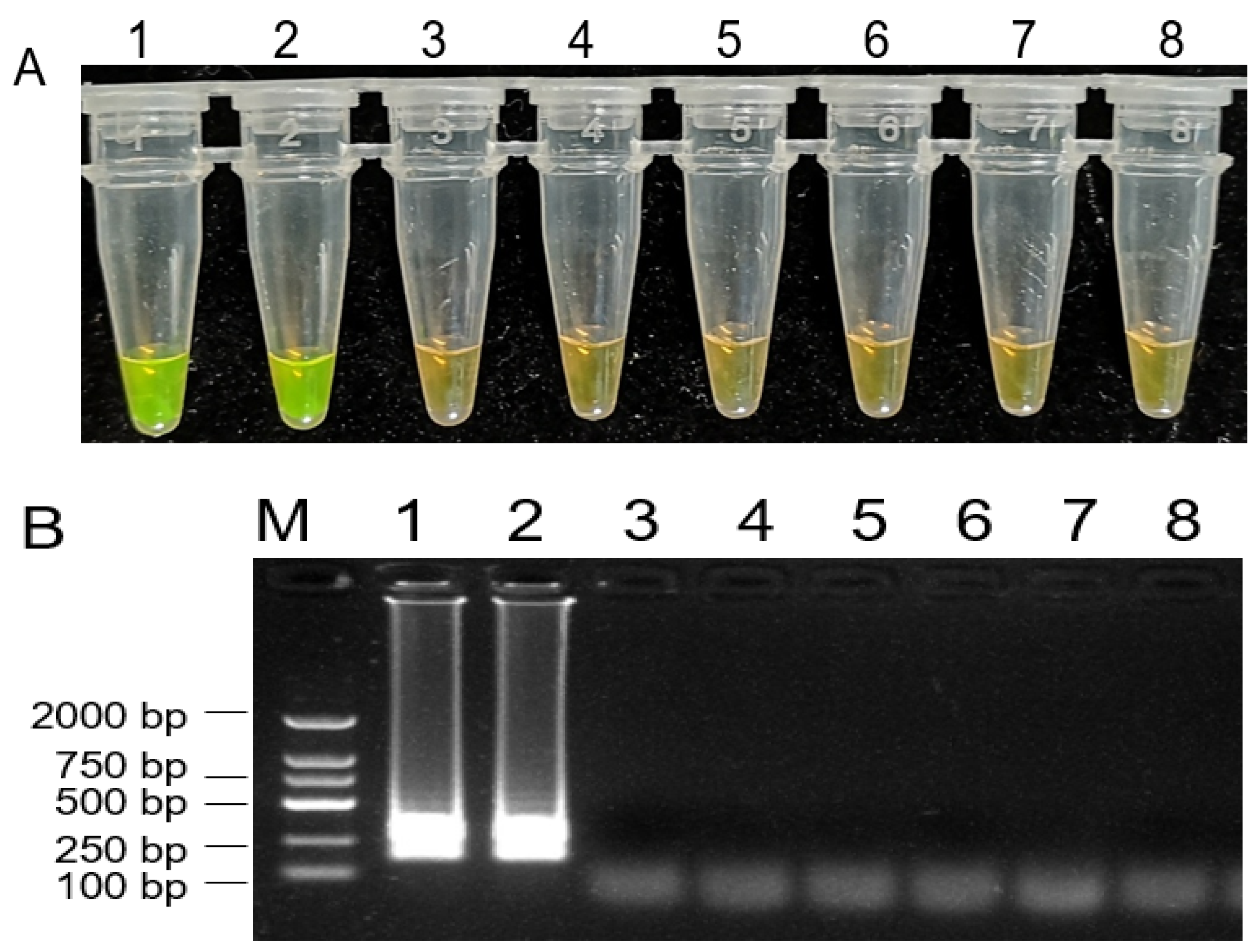
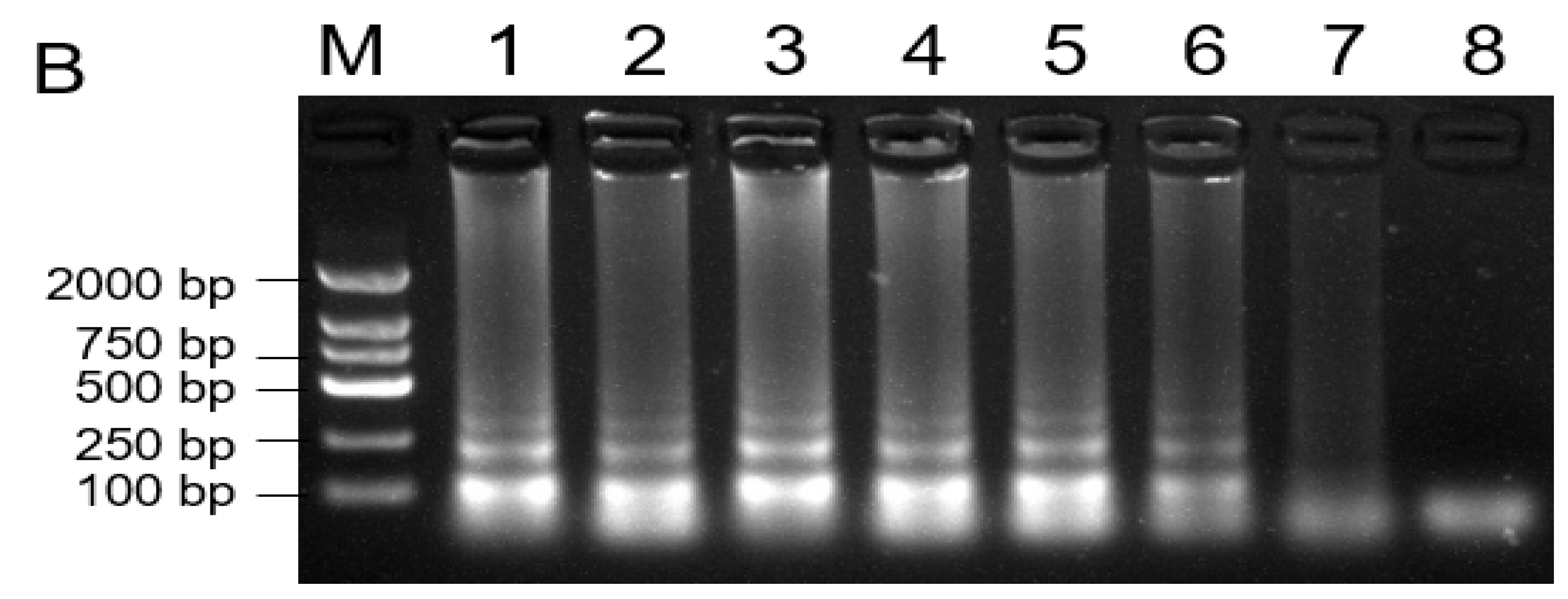
2.6. Assessment of LAMP Assay Sensitivity
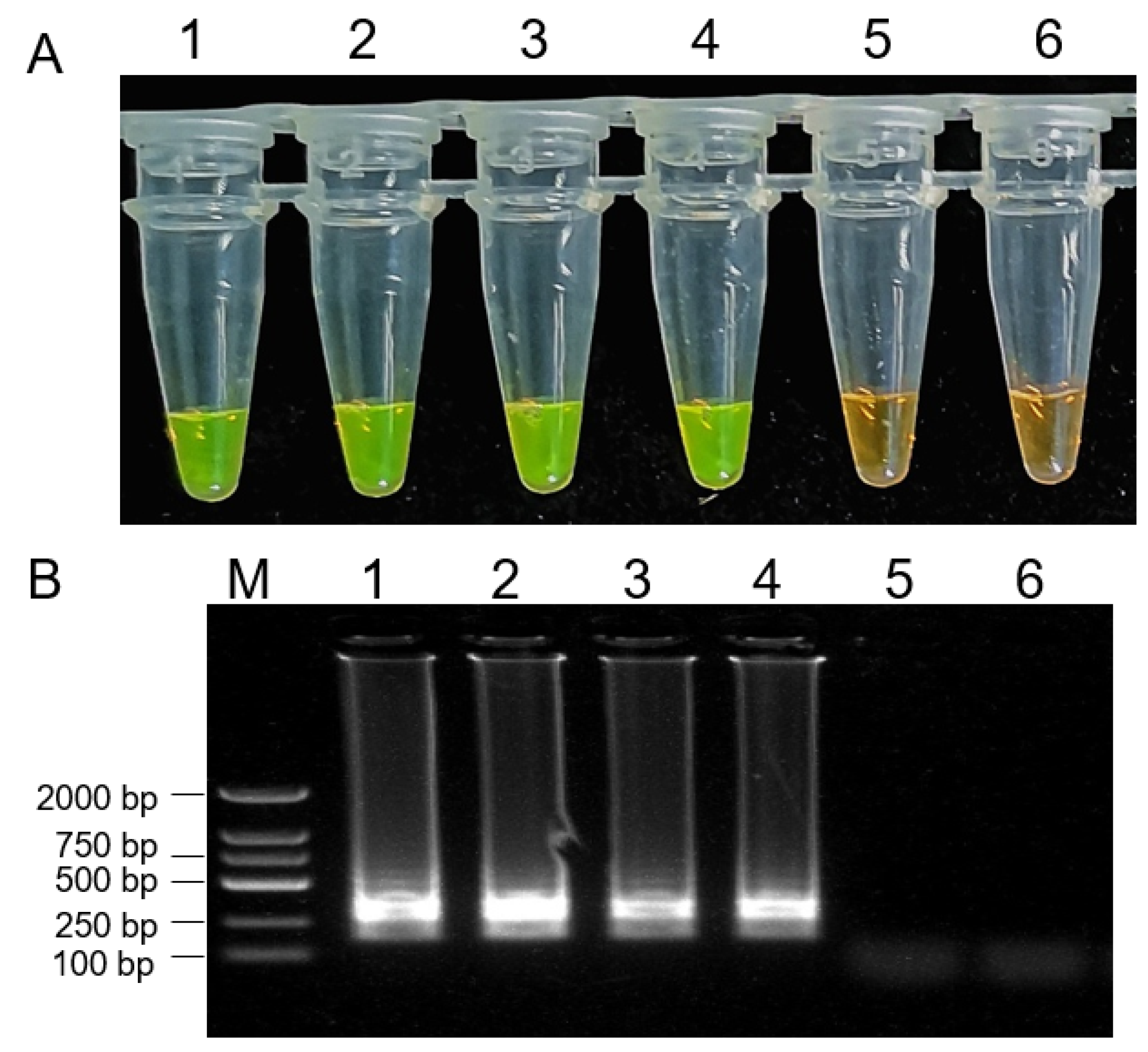
2.7. LAMP-Mediated Detection of B. dothidea in Fruit Samples
3. Discussion
4. Materials and Methods
4.1. Isolates Used and Morphological Observation
4.2. DNA Extraction, Sequencing, and Analysis
4.3. Pathogenicity Testing
4.4. Reisolation and Identification of Infected Fungi
4.5. LAMP Primers Design for the Detection of Botryosphaeria dothidea
4.6. Assessment of LAMP Assay Sensitivity
4.7. LAMP-Mediated Detection of B. dothidea in Fruit Samples
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, H.W. Huang History of 100 Years of Domestication and Improvement of Kiwifruit and Gene Discovery from Genetic Introgressed Populations in the Wild. Chin. Bull. Bot. 2009, 44, 127–142. [Google Scholar]
- Eisa, N.; Mansoureh, M.; Samira, P.A.; Farid, B. Characterization of Botryosphaeria dothidea as new pathogen of kiwifruit in Iran. J. Plant Prot. Res. 2019, 59, 134–137. [Google Scholar]
- Shihang, S.; Fang, J.; Lin, M.; Qi, X.; Chen, J.; Wang, R.; Li, Z.; Li, Y.; Muhammad, A. Freezing Tolerance and Expression of β-amylase Gene in Two Actinidia arguta Cultivars with Seasonal Changes. Plants 2020, 9, 515. [Google Scholar]
- Manning, M.; Burdon, J.; Silva, N.D.; Meier, X.; Pidakala, P.; Punter, M.; Billing, D. Maturity and postharvest temperature management affect rot expression in ‘Hort16A’ kiwifruit. Postharvest Biol. Technol. 2016, 113, 40–47. [Google Scholar] [CrossRef]
- Hawthorne, B.T.; Reid, M.S. Possibility for fungicidal control of kiwifruit fungal storage rots. N. Z. J. Exp. Agric. 1982, 10, 333–336. [Google Scholar] [CrossRef]
- Opgenorth, D.C. Storage rot of California-grown kiwifruit. Plant Dis. 1983, 67, 382–383. [Google Scholar] [CrossRef] [Green Version]
- Willetts, H.J.; Wong, J.A.L.; Kirst, G.D. The biology of Sclerotinia sclerotiorum, Sclerotinia trifoliorum and Sclerotinia minor with emphasis on specific nomenclature. Bot. Rev. 1980, 46, 101–165. [Google Scholar] [CrossRef]
- Manning, M.A.; Meier, X.; Olsen, T.L.; Johnston, P.R. Fungi associated with fruit rots of Actinidia chinensis ‘Hort16A’ in New Zealand. N. Z. J. Crop Hortic. Sci. 2003, 31, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Koh, Y.J.; Hur, J.S.; Jung, J.S. Postharvest fruit rot of kiwifruit (Actinidia deliciosa) in Korea. N. Z. J. Crop Hortic. Sci. 2005, 33, 303–310. [Google Scholar] [CrossRef]
- Luongo, L.; Santori, A.; Riccioni, L.; Belisario, A. Phomopsis sp. associated with post-harvest fruit rot of kiwifruit in Italy. J. Plant Pathol. 2011, 93, 205–209. [Google Scholar]
- Erper, I.; Turkkan, M.; Ozcan, M.; Luongo, L.; Belisario, A. Characterization of Diaporthe hongkongensis species causing stem-end rot on kiwifruit in Turkey. J. Plant Pathol. 2017, 99, 3. [Google Scholar]
- Marsberg, A.; Kemler, M.; Jami, F.; Nagel, J.H.; Postma-Smidt, A.; Naidoo, S.; Wingfield, M.J.; Crous, P.W.; Spatafora, J.W.; Hesse, C.N. Botryosphaeria dothidea: A latent pathogen of global importance to woody plant health. Mol. Plant Pathol. 2017, 18, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Pan, H.; Chen, M.; Zhang, S.; Zhong, C. Isolation and identification of pathogenic fungi causing postharvest fruit rot of kiwifruit (Actinidia chinensis) in China. J. Phytopathol. 2017, 165, 782–790. [Google Scholar] [CrossRef]
- Phillips, A.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Slippers, B.; Crous, P.W.; Denman, S.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Combined multiple gene genealogies and phenotypic characters differentiate several species previously identified as Botryosphaeria dothidea. Mycologia. 2004, 96, 83–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslam, S.; Tahir, A.; Aslam, M.F.; Alam, M.W.; Shedayi, A.A.; Sadia, S. Recent advances in molecular techniques for the identification of phytopathogenic fungia mini review. J. Plant Interact. 2017, 12, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Hameed, U.; Zhang, A.F.; Zang, H.Y.; Gu, C.Y.; Chen, Y.; Xu, Y.L. Development of a nested-PCR assay for the rapid detection of Pilidiella granati in pomegranate fruit. Sci. Rep. 2017, 7, 40954. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Zhang, H.; Chi, F.; Ji, Z.; Dong, Q.; Cao, K.; Zhou, Z. Species-specific PCR-based assays for identification and detection of Botryosphaeriaceae species causing stem blight on blueberry in China. J. Integr. Agric. 2016, 15, 573–579. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef] [Green Version]
- Goto, M.; Honda, E.; Ogura, A.; Nomoto, A.; Hanaki, K.I. Colorimetric detection of loop-mediated isothermal amplification reaction by using hydroxy naphthol blue. Biotechniques 2009, 46, 167–172. [Google Scholar] [CrossRef]
- Moradi, A.; Nasiri, J.; Abdollahi, H.; Almasi, M. Development and evaluation of a loop-mediated isothermal amplification assay for detection of Erwinia amylovora based on chromosomal DNA. Eur. J. Plant Pathol. 2012, 133, 609–620. [Google Scholar] [CrossRef]
- Niessen, L.; Vogel, R.F. Detection of Fusarium graminearum DNA using a loop-mediated isothermal amplification (LAMP) assay. Int. J. Food Microbiol. 2010, 140, 183–191. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, C.Q. q-LAMP assays for the detection of Botryosphaeria dothidea causing Chinese hickory canker in trunk, water, and air samples. Plant Dis. 2019, 103, 3142–3149. [Google Scholar] [CrossRef] [PubMed]
- Shaoli, W.; Henghua, Q.; Yingzi, W.; Peisong, W.; Binghui, L.; Guanhua, W. Development of a loop-mediated isothermal amplification assay for rapid detection of apple ring rot pathogen Botryosphaeria dothidea. Chin. J. Plant Prot. 2020, 47, 127–133. [Google Scholar]
- Gyoung, H.K.; Young, J.K. Diagnosis and Integrated Management of Major Fungal Fruit Rots on Kiwifruit in Korea. Res. Plant Dis. 2018, 24, 113–122. [Google Scholar]
- Michailides, T.J.; Elmer, P.A.G. Botrytis gray mold of kiwi-fruit caused by Botrytis cinerea in the United States and New Zealand. Plant Dis. 2000, 84, 208–223. [Google Scholar] [CrossRef] [Green Version]
- Pennycook, S.R. Sclerotinia rot of kiwifruit (Actinidia chinensis). Orchardist N. Z. 1982, 55, 407–408. [Google Scholar]
- Pennycook, S.R. Fungal fruit rots of Actinidia deliciosa (kiwifruit). N. Z. J. Exp. Agric. 1985, 13, 289–299. [Google Scholar]
- Li, G.; Huang, G.; Zhu, L.; Lv, D.; Cao, B.; Liao, F.; Luo, J. Loop-mediated isothermal amplification (LAMP) detection of Phytophthora hibernalis, P. syringae and P. cambivora. J. Plant Pathol. 2019, 101, 51–57. [Google Scholar] [CrossRef]
- Pan, W.; Shen, H.; Zhao, M. Development and application of the novel visual loop-mediated isothermal amplification of Omp25 sequence for rapid detection of Brucella sp. J. Anim. Vet. Adv. 2011, 10, 2120–2126. [Google Scholar] [CrossRef] [Green Version]
- Przewodowska, A.; Zacharzewska, B.; Chołuj, J.; Treder, K. A one-step, real-time reverse transcription loop mediated isothermal amplification assay to detect potato virus Y. Am. J. Potato Res. 2015, 92, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Najafzadeh, M.; Vicente, V.; Xi, L.; De, H.G. Rapid detection of pathogenic fungi using loop-mediated isothermal amplification, exemplified by fonsecaea agents of chromoblastomycosis. J. Microbiol. Methods. 2010, 80, 19–24. [Google Scholar] [CrossRef]
- Wang, S.; Ye, W.; Tian, Q.; Dong, S.; Zheng, X. Rapid detection of Colletotrichum gloeosporioides using a loop-mediated isothermal amplification assay. Australas. Plant Pathol. 2017, 46, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Fan, F.; Yin, W.; Li, G.; Lin, Y.; Luo, C. Development of a LAMP method for detecting SDHI fungicide resistance in Botrytis cinerea. Plant Dis. 2018, 102, 1612–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathur, K.; Thakur, R.P.; Rao, V.P. Comparison of single-spore and hyphal-tip isolates of Colletotrichum graminicola for morphological and pathogenic diversity. J. Mycol. Plant Pathol. 2000, 30, 359–365. [Google Scholar]
- Tang, W.; Ding, Z.; Zhou, Z.; Wang, Y.; Guo, L. Phylogenetic and pathogenic analyses show that the causal agent of apple ring rot in China is Botryosphaeria dothidea. Plant Dis. 2012, 96, 486–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möller, E.; Bahnweg, G.; Sandermann, H.; Geiger, H. A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucleic Acids Res. 1992, 20, 6115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, P.L.; Francesca, P.H.; Gubler, W.D.; Florent, P.T. Botryosphaeriaceae species associated with dieback and canker disease of bay laurel in northern California with the description of Dothiorella californica sp. nov. Fungal Biol. 2017, 121, 347–360. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing offungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press, Inc.: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- An, J.; Guo, L.; Zhu, X.; Song, Y. Susceptibility of Botryosphaeria dothidea to tebuconazole and thiophanate-methyl. Chin. Plant Pro. 2016, 42, 187–191. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.H.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, L.N.; Woodward, J.F. Koch’s postulates: An interventionist perspective. Stud. Hist. Philos. Biol. Biomed. Sci. 2016, 59, 35–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.; Yang, Y.; Wang, Y.; Pan, X.; Wu, J.; Cai, Y.; Li, T.; Zhao, D.; Wang, J.; Zhou, M. Loop-mediated isothermal amplification for the rapid detection of the F200Y mutant genotype of carbendazim-resistant isolates of Sclerotinia sclerotiorum. Plant Dis. 2016, 100, 976–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Hou, H.; Zhou, Z.; Tu, H.; Yuan, H. Identification and Detection of Botryosphaeria dothidea from Kiwifruit (Actinidia chinensis) in China. Plants 2021, 10, 401. https://doi.org/10.3390/plants10020401
Wang L, Hou H, Zhou Z, Tu H, Yuan H. Identification and Detection of Botryosphaeria dothidea from Kiwifruit (Actinidia chinensis) in China. Plants. 2021; 10(2):401. https://doi.org/10.3390/plants10020401
Chicago/Turabian StyleWang, Li, Hui Hou, Zengqiang Zhou, Hongtao Tu, and Hongbo Yuan. 2021. "Identification and Detection of Botryosphaeria dothidea from Kiwifruit (Actinidia chinensis) in China" Plants 10, no. 2: 401. https://doi.org/10.3390/plants10020401
APA StyleWang, L., Hou, H., Zhou, Z., Tu, H., & Yuan, H. (2021). Identification and Detection of Botryosphaeria dothidea from Kiwifruit (Actinidia chinensis) in China. Plants, 10(2), 401. https://doi.org/10.3390/plants10020401