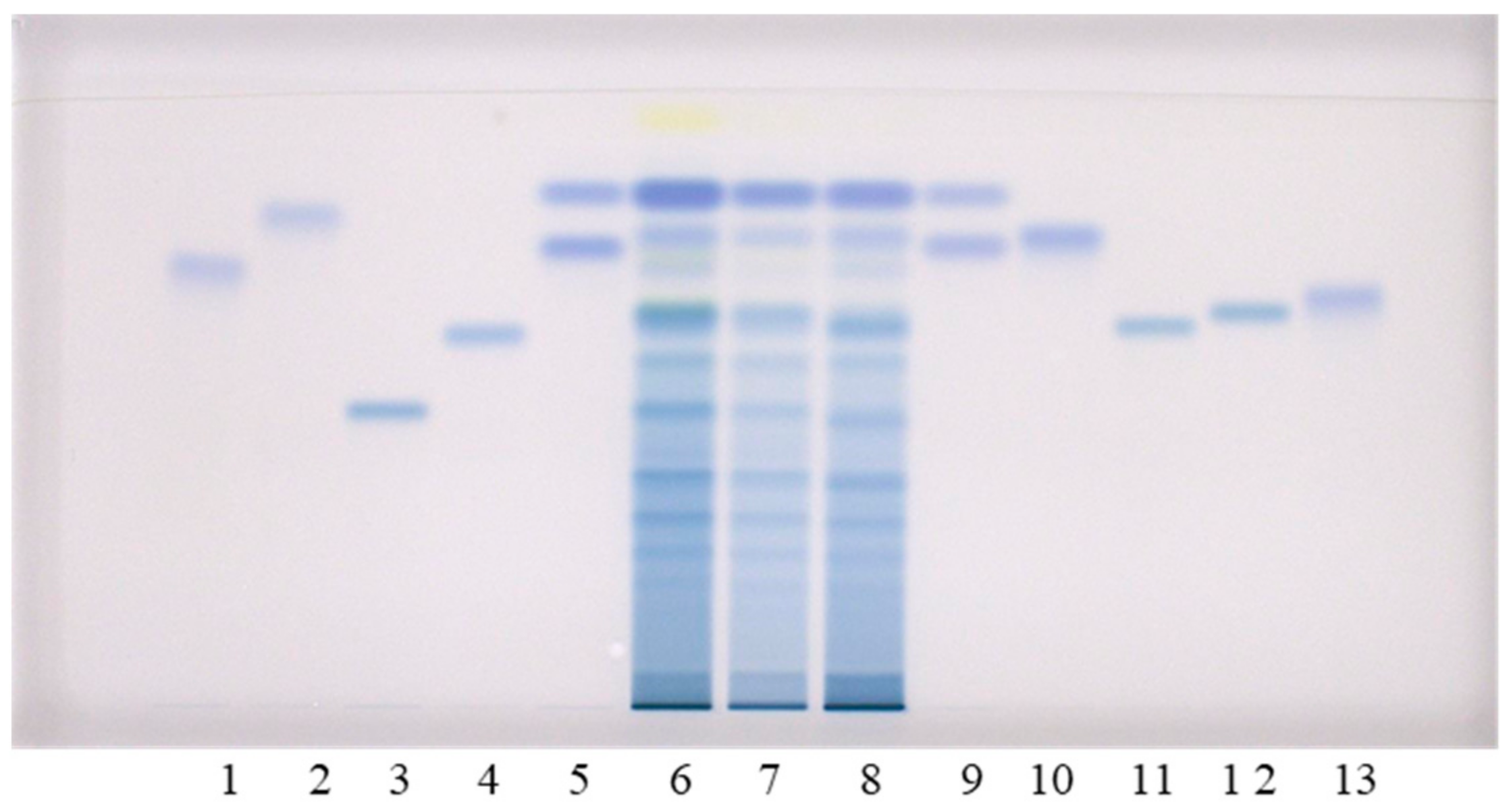
2.1. Qualitative Analyses of Flavan-3-ols and Proanthocyanidins with HPTLC
Qualitative analyses of flavan-3-ols and proanthocyanidins in the sample test solutions (STSs) prepared from rhizomes of Japanese, Bohemian and giant knotweed were performed using the high performance thin-layer chromatography (HPTLC) method on the HPTLC silica gel plates developed with toluene–acetone–formic acid (3:6:1,
v/v) [
9,
16]. Post-chromatographic derivatization with 4-dimethylaminocinnamaldehyde (DMACA) reagent [
17] enabled the detection of flavan-3-ols and proanthocyanidins, which was more selective than the detection before derivatization. We previously used this method in two studies: (1) an HPTLC/MS/MS study on flavan-3-ols and proanthocyanidins in rhizomes of Japanese knotweed [
9]; and (2) an HPTLC study on flavan-3-ols and proanthocyanidins in leaves of Japanese, Bohemian and giant knotweed [
18]. In this study, the following twelve standards were used for analyses: (+)-catechin, (−)-epicatechin, (−)-catechin gallate, (−)-epicatechin gallate, (−)-gallocatechin, (−)-epigallocatechin, (−)-gallocatechin gallate, (−)-epigallocatechin gallate, procyanidin B1, procyanidin B2, procyanidin B3 and procyanidin C1 (
Figure 1). Eight of these twelve standards ((−)-catechin gallate (track 2), (−)-epicatechin gallate (track 10), (−)-gallocatechin gallate (track 1), (−)-epigallocatechin gallate (track 13), procyanidin B1 (track 11), procyanidin B2 (track 12), procyanidin B3 (track 4) and procyanidin C1 (track 3)) were separated from each other (
Figure 1). The eight standards were also separated from the two pairs of unseparated standards (
Figure 1). The first of the two pairs of unseparated standards includes (+)-catechin (track 9, higher R
F) and (−)-epicatechin (track 5, higher R
F). The second pair includes (−)-gallocatechin (track 9, lower R
F) and (−)-epigallocatechin (track 5, lower R
F). As shown in
Figure 1, compounds (−)-epicatechin gallate (track 10), (−)-gallocatechin gallate (track 1), dimer procyanidin B2 (track 12) and trimer procyanidin C1 (track 3) were detected as intensive blue bands in STSs prepared from Japanese (track 6), Bohemian (track 7) and giant (track 8) knotweed rhizomes.
Additionally, intensive blue bands, present in all STSs from rhizomes (tracks 6–8), at the R
F of standards of monomers ((+)-catechin and (−)-epicatechin) confirmed the presence of monomers in rhizomes of all studied knotweeds (
Figure 1). Procyanidin B1 (track 11) and procyanidin B3 (track 4) were only detected in STS from giant knotweed (
Figure 1, tracks 6–8). Based on other standards (−)-catechin gallate (track 2), (−)-gallocatechin (track 9, lower R
F), (−)-epigallocatechin (track 5, lower R
F) and (−)-epigallocatechin gallate (track 13) were not detected in STSs of the three knotweed rhizomes (
Figure 1, tracks 6–8).
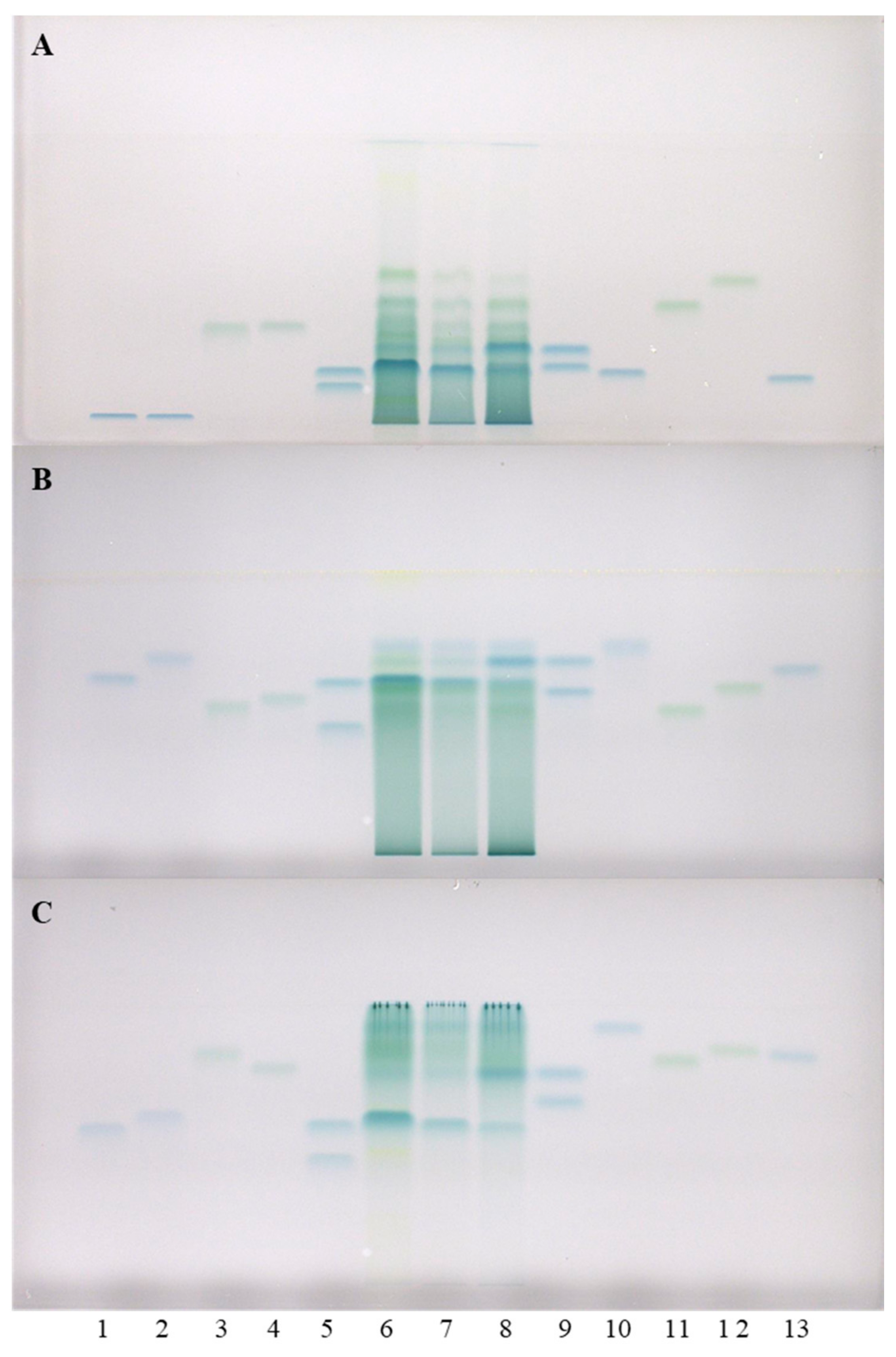
To further examine the obtained results, additional analyses of all three STSs from rhizomes were performed on HPTLC cellulose plates, using our previously developed methodology [
17,
19,
20]. This methodology includes three developing solvents (water [
17,
19,
20], 1-propanol–water–acetic acid (4:2:1,
v/v) [
17,
20] and 1-propanol–water–acetic acid (20:80:1,
v/v) [
20,
21,
22]) and post-chromatographic derivatization with DMACA reagent [
17]. This approach provided complementary results. The two pairs of standards (1st: (+)-catechin (track 9, higher R
F) and (−)-epicatechin (track 5, higher R
F); 2nd: (−)-gallocatechin (track 9, lower R
F) and (−)-epigallocatechin (track 5, lower R
F)) that were not separated on HPTLC silica gel plates, were separated on the HPTLC cellulose plate developed with water (
Figure 2). Furthermore (+)-catechin and (−)-epicatechin were separated from all other standards on the HPTLC cellulose plates which were developed with water (
Figure 2A) and 1-propanol–water–acetic acid (4:2:1 (
v/v),
Figure 2B). As shown in
Figure 2, (+)-catechin (track 9, higher R
F), (−)-epicatechin (track 5, higher R
F), (−)-epicatechin gallate (track 10) and procyanidin B2 (track 12) were detected in Japanese (track 6), Bohemian (track 7) and giant (track 8) knotweed on the HPTLC cellulose plates developed with all three developing solvents. (−)-catechin gallate (track 2) was detected in STSs of all three knotweeds only on the plates developed with 1-propanol–water–acetic acid (4:2:1 (
v/v),
Figure 2B), and in STS of Japanese knotweed on the plate developed with 1-propanol–water–acetic acid (20:80:1 (
v/v),
Figure 2C). (−)-Gallocatechin (
Figure 2A, track 9, lower R
F) was detected in all three knotweeds only on the plates developed with water. Procyanidin B3 (track 4) was also detected in all three knotweeds only on the plate developed with water (
Figure 2A), and was additionally detected in giant knotweed on the plate developed with 1-propanol–water–acetic acid (20:80:1 (
v/v),
Figure 2C). (−)-Epigallocatechin (
Figure 2, track 5, lower R
F) was not detected in any of the studied STSs from rhizomes. (−)-Gallocatechin gallate was detected in all three knotweeds, but only on the plates developed with the developing solvents containing 1-propanol–water–acetic acid (
Figure 2B,C) and not on the plate developed with water (
Figure 2A). (−)-Epigallocatechin gallate (track 13) was detected in all three knotweeds, but only on the plates developed with 1-propanol–water–acetic acid (20:80:1 (
v/v),
Figure 2C). Procyanidin B1 (track 11) was detected in all three knotweeds on the plates developed with water (
Figure 2A) and 1-propanol–water–acetic acid (4:2:1 (
v/v),
Figure 2B), and only detected in giant knotweed on the plate developed with 1-propanol–water–acetic acid (20:80:1 (
v/v),
Figure 2C). Procyanidin C1 (track 3) was detected in all three STSs on the HPTLC cellulose plates developed with 1-propanol–water–acetic acid (4:2:1 (
v/v),
Figure 2B and 20:80:1 (
v/v),
Figure 2C). Procyanidin C1 was also detected in Bohemian and giant knotweed on the plate developed with water (
Figure 2A).
Table 1 summarizes the results of the HPTLC analyses obtained with all four chromatographic systems involving two HPTLC stationary phases (silica gel and cellulose) in combination with the developing solvents used (toluene–acetone–formic acid (3:6:1,
v/v) for silica gel plates (
Figure 1); water and 1-propanol–water–acetic acid in different ratios (4:2:1,
v/v; 20:80:1,
v/v) for cellulose plates (
Figure 2)). Data from
Table 1 show that complementary methods are crucial to avoid reporting false positive results. In certain cases, as illustrated in
Table 1, different combinations of the stationary phase and the developing solvent gave different results. The final confirmation of compounds in STSs was only given to those compounds which were detected using both stationary phases (column “Both” in
Table 1) and all developing solvents. Based on these criteria we confirmed the presence of (+)-catechin, (−)-epicatechin, (−)-epicatechin gallate and procyanidin B2 (dimer) in rhizomes of Japanese, Bohemian and giant knotweed, and the presence of procyanidin B1 (dimer) and procyanidin C1 (trimer) in rhizomes of giant knotweed. (+)-Catechin and (−)-epicatechin were previously found in Japanese [
8,
15] and giant knotweed rhizomes [
8]. In our recent study we isolated (+)-catechin, (−)-epicatechin, (−)-epicatechin gallate, procyanidin B1, procyanidin B2, procyanidin B3, proanthocyanidin B dimer gallate and some other compounds from rhizomes and bark of the rhizomes of Japanese knotweed [
15].
To the best of our knowledge this is the first report on (+)-catechin, (−)-epicatechin, (−)-epicatechin gallate and procyanidin B2 (dimer) in Bohemian knotweed rhizomes. This is also the first report on (−)-epicatechin gallate, procyanidin B1 (dimer), procyanidin B2 (dimer) and procyanidin C1 (trimer) in giant knotweed rhizomes.
2.2. Chromatographic Fingerprinting (HPTLC) of Proanthocyanidins with Densitometry and Image Analysis
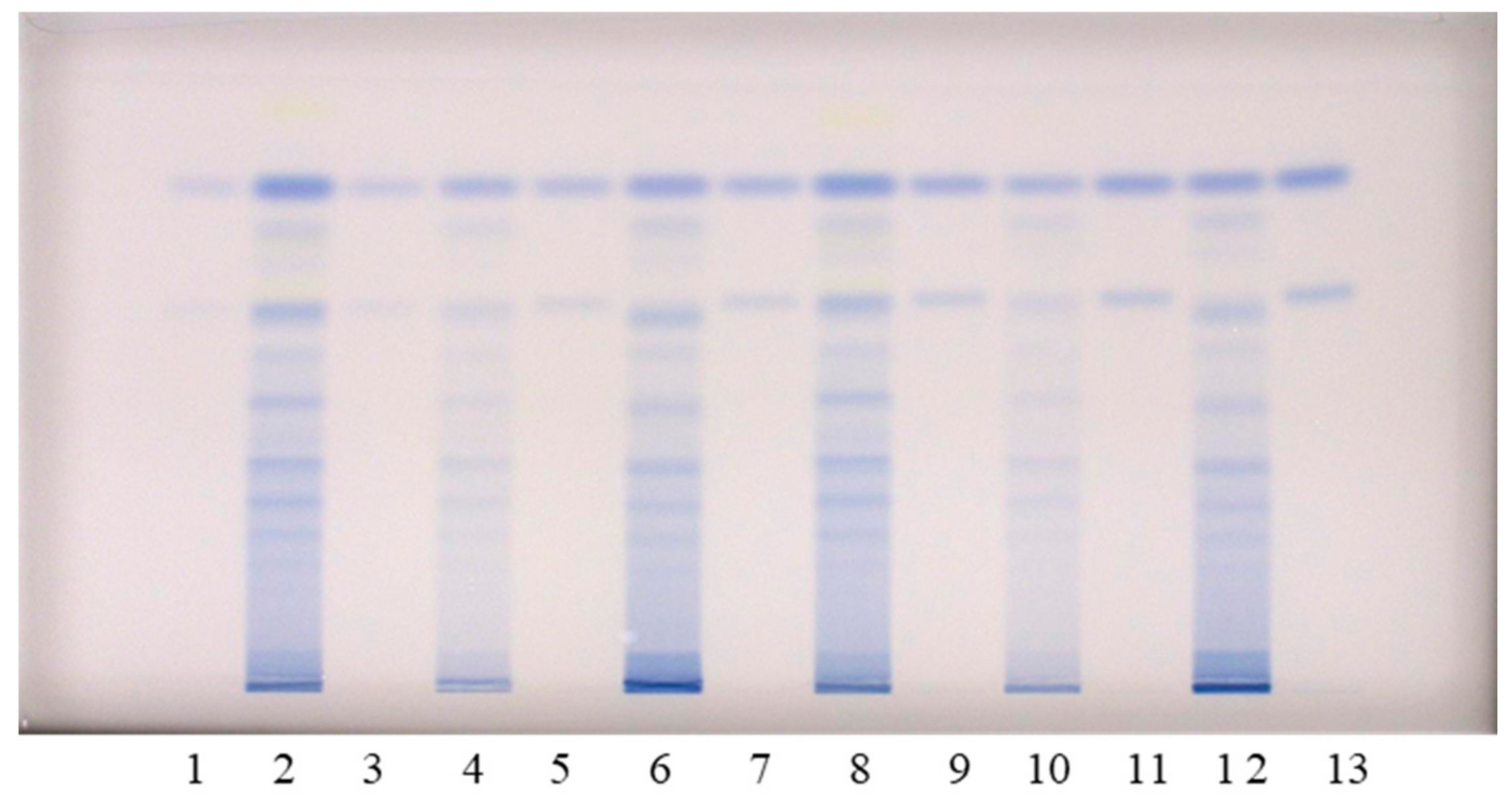
The chromatographic fingerprinting of proanthocyanidins in STSs from rhizomes of Japanese, Bohemian and giant knotweed on HPTLC silica gel plate was performed with densitometry and image analysis. Only densitometry was applied for the chromatographic fingerprinting of proanthocyanidins on the developed plates (before post-chromatographic derivatization). Additionally, densitometry and image analysis were used for fingerprinting after post-chromatographic derivatization with DMACA reagent (
Figure 3). To detect underivatized proanthocyanidins and their derivatives, formed when proanthocyanidins react with DMACA reagent, densitometric scanning was performed in the absorption/reflectance mode at two wavelengths 280 nm (
Figure 4A) and 655 nm (
Figure 4B), re spectively. As evident from the densitograms of (−)-epicatechin and procyanidin B2 standards before (
Figure 4A) and after (
Figure 4B) post-chromatographic derivatization, derivatization with DMACA reagent significantly enhanced the sensitivity of the HPTLC method. This enhancement of the sensitivity is shown in a considerable increase in the heights of the peaks in the densitograms scanned after the derivatization for both standards and all STSs at R
F values for monomers (R
F of (−)-epicatechin) and dimers (R
F of procyanidin B2) of B-type proanthocyanidins (
Figure 4B). The densitograms of all STSs contain two peaks at the same R
F values as the standard solutions of (−)-epicatechin and procyanidin B2. These peaks confirm that Japanese, Bohemian and giant knotweed contain monomers and dimers of B-type proanthocyanidins (
Figure 4B). Comparisons of the densitograms of each of the three STSs from rhizomes recorded on the same plate before (
Figure 4A) and after (
Figure 4B) derivatization show differences in the chromatographic fingerprints. These differences can be explained by the fact that at 280 nm the light was also absorbed by other compounds (e.g., phenolic acids, other flavonoids) which are present together with flavan-3-ols and proanthocyanidins in all three knotweed STSs. Therefore, peaks at other R
F values are also visible.
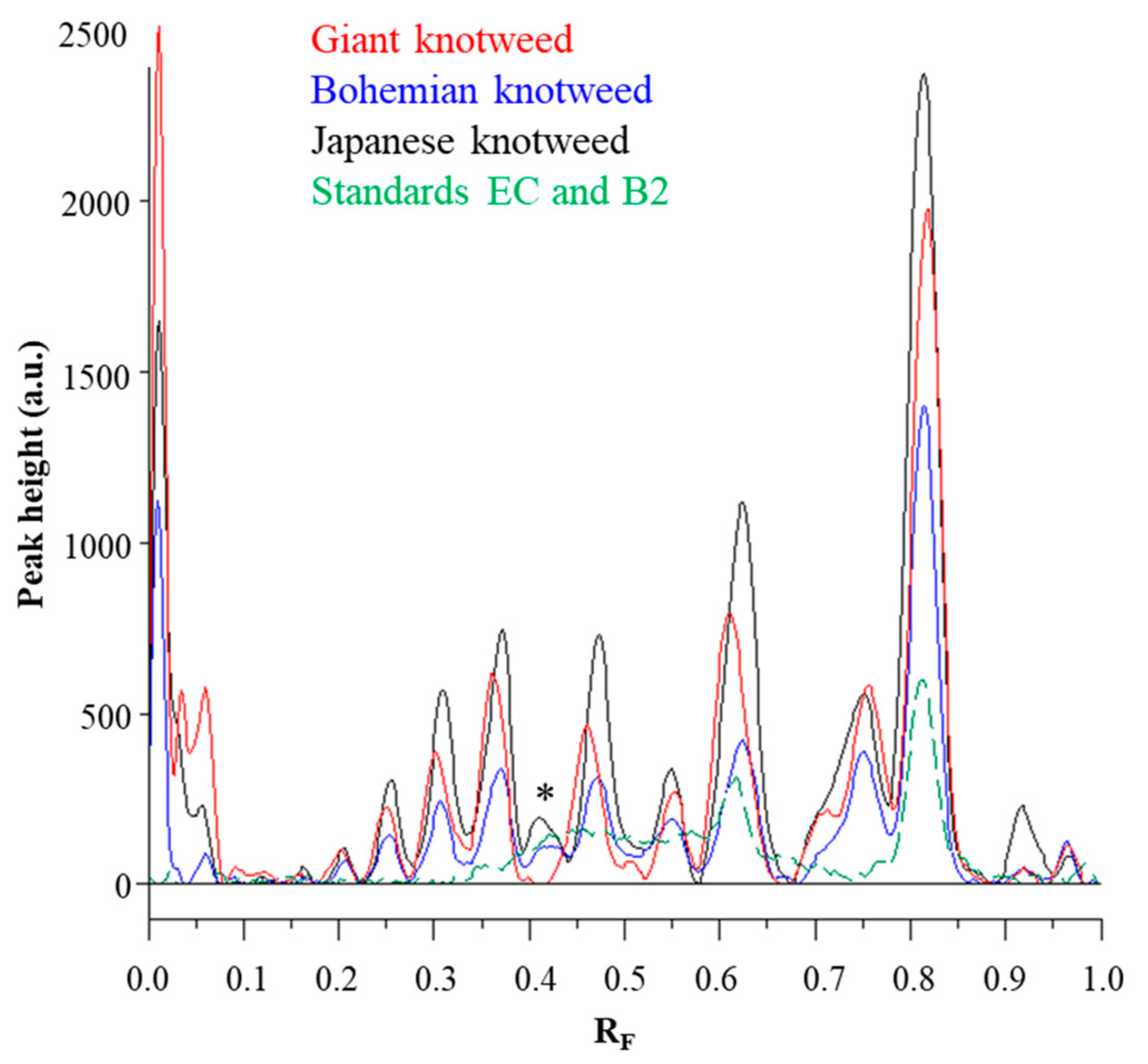
Chromatographic fingerprinting of proanthocyanidins with image analysis was performed at white light after post-chromatographic derivatization with DMACA reagent. Image analysis of the underivatized proanthocyanidins could not be performed at 280 nm due to the technical limitations of the DigiStore 2 documentation system, which in the UV zone only permits the use of wavelengths of 366 and 254 nm. Therefore, the chromatographic fingerprinting of proanthocyanidins with image analysis was performed only after post-chromatographic derivatization with DMACA reagent. After post-chromatographic derivatization the images of the HPTLC silica gel plates were captured at white light illumination. The images (
Figure 3) were then converted to videodensitograms (
Figure 5 and
Figure 6) in absorption mode. The videodensitograms of STSs from rhizomes of all three knotweeds show peaks at R
F values of (−)-epicatechin and procyanidin B2 standards, which confirms the presence of monomers and dimers in all knotweed STSs (
Figure 5). Comparisons of the videodensitograms of STSs from rhizomes showed that Japanese and Bohemian knotweed have equal qualitative profiles, which also included the peak at R
F 0.41, which was not detected in giant knotweed (
Figure 5). Additionally, from the peak heights in vid eodensitograms of all STSs it is evident that Japanese knotweed is the richest source of proanthocyanidins (
Figure 5). The chromatographic fingerprints of rhizomes of Japanese, Bohemian and giant knotweed were also compared with the chromatographic fingerprints of leaves obtained from the respective plant species and collected at the same locations [
18] as the rhizomes. The STSs of leaves were prepared as described in our previous study [
18].
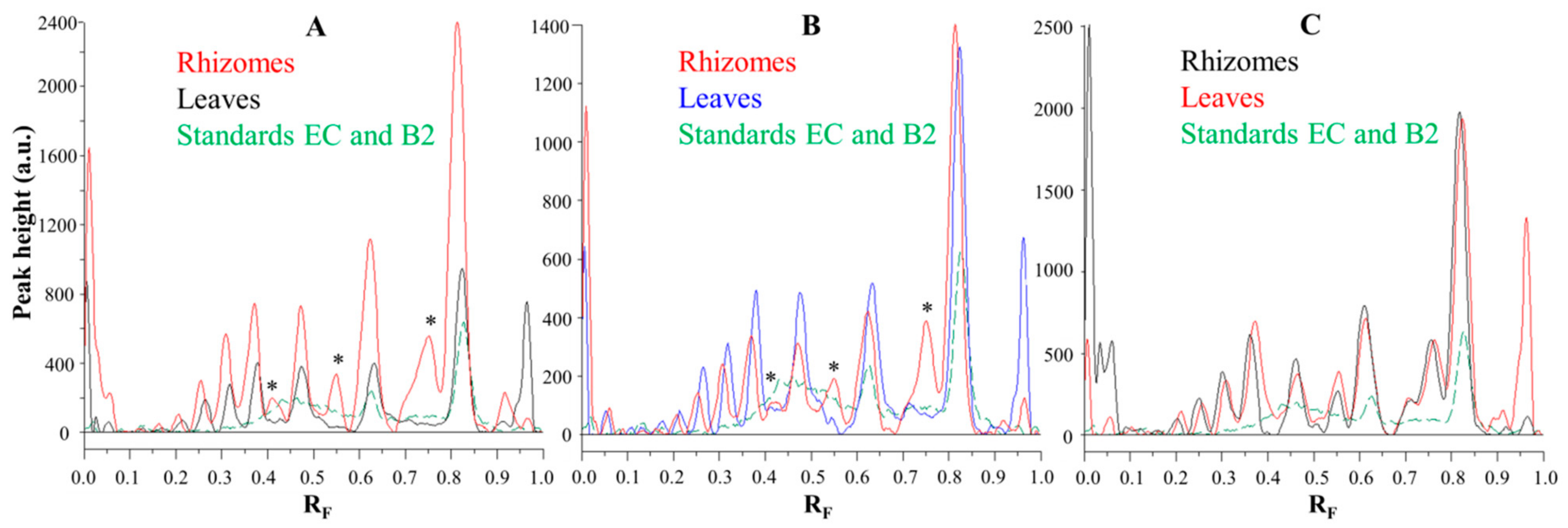
Comparisons of the chromatographic fingerprints of the rhizomes and leaves of Japanese (
Figure 6A), Bohemian (
Figure 6B) and giant (
Figure 6C) showed that within the individual knotweed species giant knotweed rhizomes and leaves have the most similar fingerprints (
Figure 6C), while the fingerprints of Japanese (
Figure 6A) and Bohemian (
Figure 6B) knotweed rhizomes and leaves differ in the peaks at R
F 0.41, 0.55, and 0.75 which are only present in rhizomes (
Figure 6A,B). The smallest differences in the heights of peaks at R
F values of (−)-epicatechin and procyanidin B2 standards were observed between the videodensitograms of rhizomes and leaves of giant knotweed (
Figure 6C). The biggest differences at these R
Fs were observed in the heights of peaks between rhizomes and leaves of Japanese knotweed (
Figure 6A), with significantly higher peaks for rhizomes. In the case of Bohemian knotweed the differences between the peak heights of rhizomes and leaves are less pronounced. However, for rhizomes the peak at R
F of (−)-epicatechin is higher than for leaves, and the peak at R
F of procyanidin B2 standard is lower than for leaves.
2.3. Quantification of Proanthocyanidins
HPTLC combined with image analysis was applied for the quantitative determination of proanthocyanidins with the focus on: (1) differences in the contents of monomers and dimers in rhizomes of all three knotweeds; (2) total proanthocyanidins content. Im ages of the chromatograms were converted to videodensitograms, which were integrated using VideoScan software. Limits of detection (LOD), limits of quantification (LOQ) and the stability of the derivatized standards of (−)-epicatechin (100 ng) and procyanidin B2 (100 ng) on the HPTLC silica gel plates were described in our previous study [
18]. STSs from rhizomes were analyzed on an HPTLC silica gel plate (
Figure 4) using equal amounts (30, 40, 60, 80, 100, 120 and 150 ng per plate) of (−)-epicatechin and procyanidin B2 standards for the calibration curves. Equal volumes of each STS were applied twice on the plate (once on the left and once on the right side of the plate). For quantification the mean value of the peak areas from both applications (at the same R
F) was cal culated (data-pair technique), which reduced the effect of inequity of the stationary phase. The equations of the calibration curves based on peak areas of standards (−)-epicatechin and procyanidin B2 were used to calculate the contents of monomers (flavan-3-ols) and procyanidin dimers, respectively, in rhizomes of Japanese, Bohemian and giant knotweed (
Table 2). This is the first report about the quantification of the total content of monomers and dimers of proanthocyanidins in rhizomes of Japanese, Bohemian and giant knotweed. Other authors reported only the contents of separate monomers ((+)-catechin and (−)-epicatechin) [
7,
8,
23] and B-type dimers (specified as “procyanidin dimer B”) [
7,
8] in the rhizomes of Japanese [
7,
8,
23] and giant [
7,
8] knotweed. The total contents of monomers and dimers expressed per dry mass of the plant material (DM) for all STSs were the highest in Japanese knotweed rhizomes (5.80 kg/t DM) and the lowest in Bohemian knotweed rhizomes (2.61 kg/t DM), while it was 4.53 kg/t DM in gi ant knotweed rhizomes (
Table 2). It was determined that the content of monomers (2.36 kg/t DM) in giant knotweed rhizomes was equal to that reported for its leaves in our previous study [
18]. The contents of dimers in rhizomes (2.17 kg/t DM) and leaves (2.06 kg/t DM) [
18] of giant knotweed were also comparable. The content of monomers ob tained in rhizomes (1.52 kg/t DM) was slightly higher than in leaves (1.39 kg/t DM) [
18] of Bohemian knotweed. Even more pronounced differences were observed between the contents of dimers in rhizomes (1.09 kg/t DM) and leaves (1.40 kg/t DM) [
18] of Bohemian knotweed. The differences between Japanese knotweed rhizomes and leaves were significant. Rhizomes are more than three times richer in monomers (2.99 kg/t DM) than leaves (0.84 kg/t DM) [
18] and almost three times richer in dimers (2.81 kg/t DM) than leaves (0.99 kg/t DM) [
18].
Two other groups of authors reported separate contents of two monomers ((+)-catechin, (−)-epicatechin) [
7,
8,
23]) and B-type dimers (specified as “procyanidin di mer B” [
7,
8]) per 100 g DM. Using their data we calculated contents of monomers and dimers in their samples of rhizomes [
7,
8,
23] and leaves [
7,
8] of Japanese [
7,
8,
23] and giant [
7,
8] knotweed. The first of these two groups reported 118 to 215 mg catechin/100 g DM and from 212 to 265 mg of epicatechin/100 g DM of the samples of Japanese knotweed rhizomes [
23]. Since both compounds are monomers, we calculated that they determined from 330 to 480 mg of monomers/100 g DM of Japanese knotweed rhizomes, which is slightly higher than was determined in our study (
Table 2).
The second group of authors determined 82 mg of monomers (8 mg (+)-catechin, 74 mg (−)-epicatechin) per 100 g DM of rhizomes, and 475 mg of monomers (145 mg (+)-catechin, 330 mg (−)-epicatechin) per 100 g DM of leaves of Japanese knotweed [
8]. The monomers content in rhizomes was 3.6 times lower than in our study (
Table 2), while the monomers content in leaves was 5.6 times higher than in our previous study on Japanese knotweed leaves [
18]. In giant knotweed they found 190 mg of monomers (21 mg (+)-catechin, 169 mg (−)-epicatechin) per 100 g DM of rhizomes and 449 mg of monomers (111 mg (+)-catechin, 338 mg (−)-epicatechin) per 100 g DM of leaves [
8]. The monomers content in giant knotweed rhizomes was 1.2 times lower than in our study (
Table 2), while the monomers content in leaves was 1.9 times higher than in our study on giant knotweed leaves [
18]. Separate contents of B-type dimers (specified as “procyanidin dimer B”) were also determined in rhizomes and leaves of Japanese and giant knotweed [
8]. Using the contents of the two dimers in rhizomes and the three dimers in leaves of both species we calculated the following dimers contents in 100 g of DM: 184 mg in rhi zomes and 1251 mg in leaves of Japanese knotweed; 206 mg in rhizomes and 1125 mg in leaves of giant knotweed [
8]. The dimers content in Japanese knotweed rhizomes was 1.5 times lower than in our study (
Table 2), while the dimers content in leaves was 12.6 times higher than in our study on Japanese knotweed leaves [
18]. The dimers content in giant knotweed rhizomes was comparable with that obtained in our study (
Table 2), while the dimers content in leaves was 5.5 times higher than in our previous study on leaves [
18].
The second group of authors performed an additional study on Japanese and giant knotweed [
7], in which they used the same analytical method as in [
8], and reported separate contents of (+)-catechin, (−)-epicatechin and three B-type dimers (specified as “procyanidin dimer B”) in rhizomes and leaves of both species. The contents in 100 g of dry Japanese knotweed materials were as follows: 360 mg of monomers (90 mg (+)-catechin, 270 mg (−)-epicatechin) and 970 mg of dimers in rhizomes, and 320 mg of monomers (190 mg (+)-catechin, 130 mg (−)-epicatechin) and 520 mg of dimers in leaves [
7]. The contents in 100 g of dry giant knotweed materials were: 840 mg of monomers (220 mg (+)-catechin, 620 mg (−)-epicatechin) and 730 mg of dimers in rhizomes, and 270 mg of monomers (140 mg (+)-catechin, 130 mg (−)-epicatechin) and 450 mg of dimers in leaves of giant knotweed [
7]. The content of monomers in Japanese knotweed rhizomes was 4.4 times higher than in [
8] and 1.2 times higher than in our study (
Table 2), while the monomers content in leaves was 1.5 times lower than in [
8] and 3.8 times higher than in our study on leaves [
18]. The dimers content in Japanese knotweed rhi zomes was 5.3 times higher than in [
8] and 3.5 times higher than in our study (
Table 2), while the monomers content in leaves was 2.4 times lower than in [
8] and 5.3 times higher than in our study on leaves [
18]. The monomers content in giant knot weed rhizomes was 4.4 times higher than in [
8] and 3.6 times higher than in our study (
Table 2), while the monomers content in leaves was comparable to that obtained in our study on leaves [
18] and 1.7 times lower than in [
8]. The dimers content in giant knotweed rhizomes was 3.5 times higher than in [
8] and 3.4 times higher than in our study (
Table 2), while the dimers content in leaves was 2.5 times lower than in [
8] and 2.2 times higher than in our study on leaves [
18]. The differences in the contents obtained by the various studies arise from numerous factors, which influence the biosynthesis of these natural compounds. These factors include: the phenological stage of the plant, the position on the plant (for example leaves), the abiotic and biotic factors such as the growth conditions (temperature, soil nutrition supply, water supply, light intensity), infections with pests or diseases, attacks by herbivores and others.
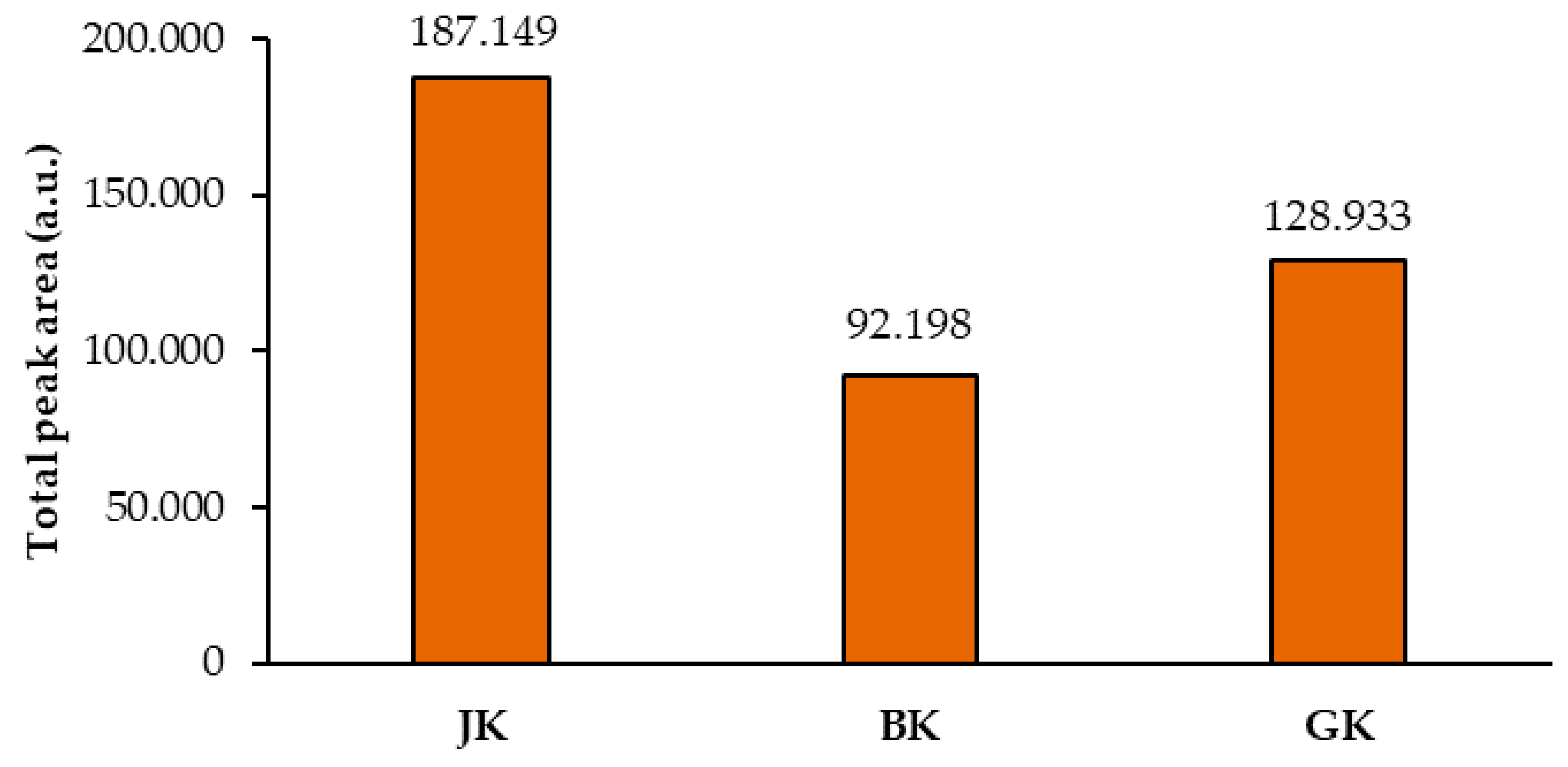
Total peak areas of the videodensitograms of the two equal applications of the same STS on HPTLC silica gel plates were used to calculate the mean of the total peak areas of proanthocyanidins (all blue bands in chromatograms). This provided additional information about proanthocyanidins in all STSs (
Figure 7). The comparison of the mean values of the total peak areas of proanthocyanidins proved the highest contents of proanthocyanidins in Japanese knotweed and the lowest in Bohemian knotweed rhizomes (
Figure 7), with the difference between both for the factor 2, and the difference between Japanese and giant knotweed for the factor 1.5. Similar differences were also obtained in our previous study of leaves of all three species, where the highest content determined in giant knotweed leaves was 1.8 and 2.2 times higher than in Bohemian and Japanese knotweed, respectively [
18].
2.4. HPTLC–MS/MS Characterization of Flavan-3-ols and Proanthocyanidins
HPTLC–MS/MS analyses of flavan-3-ols and proanthocyanidins in STSs from Bohemian and giant knotweed rhizomes were performed using the same method as in our previous studies of flavan-3-ols and proanthocyanidins in Japanese knotweed rhizomes [
10] and Japanese, Bohemian and giant knotweed leaves [
18]. Acetonitrile was used for the pre-development and development of the HPTLC diol F
254S plates. The left incised (but not cut) edge of each developed plate was broken off and derivatized with DMACA reagent, which was done to detect proanthocyanidins bands (not visible on the developed plate) and to prevent their possible degradation (due to heating) before de tection with MS. This derivatized part of the plate was then put next to the underivatized part of the plate. The blue-colored proanthocyanidins zones were then used for the proper positioning of the elution head of the TLC–MS interface on the parallel zones with the same R
F on the underivatized part of the plate. Only the underivatized zones were eluted from the plate and transferred to the MS detector for acquisition of the MS, MS
2 and MS
3 spectra.
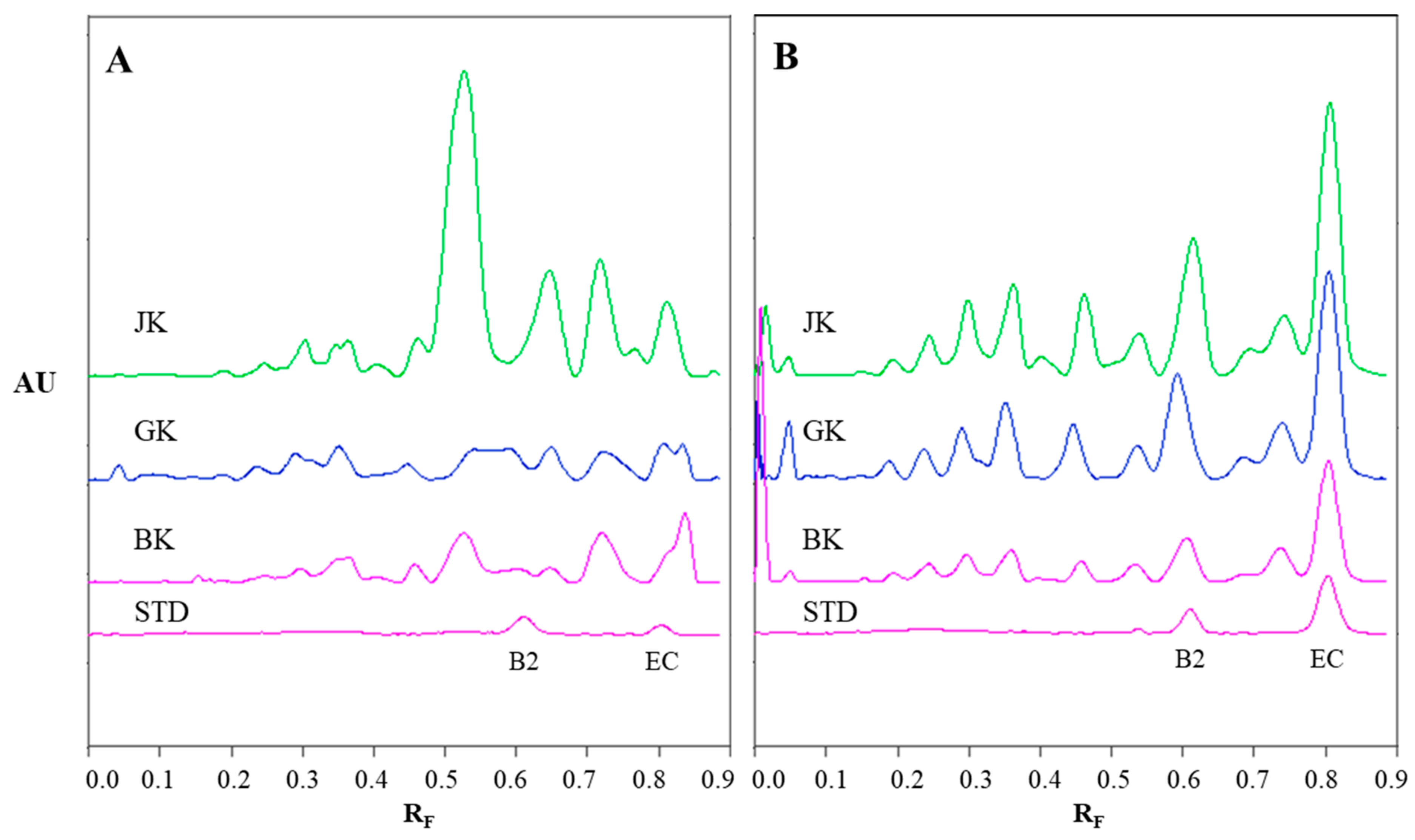
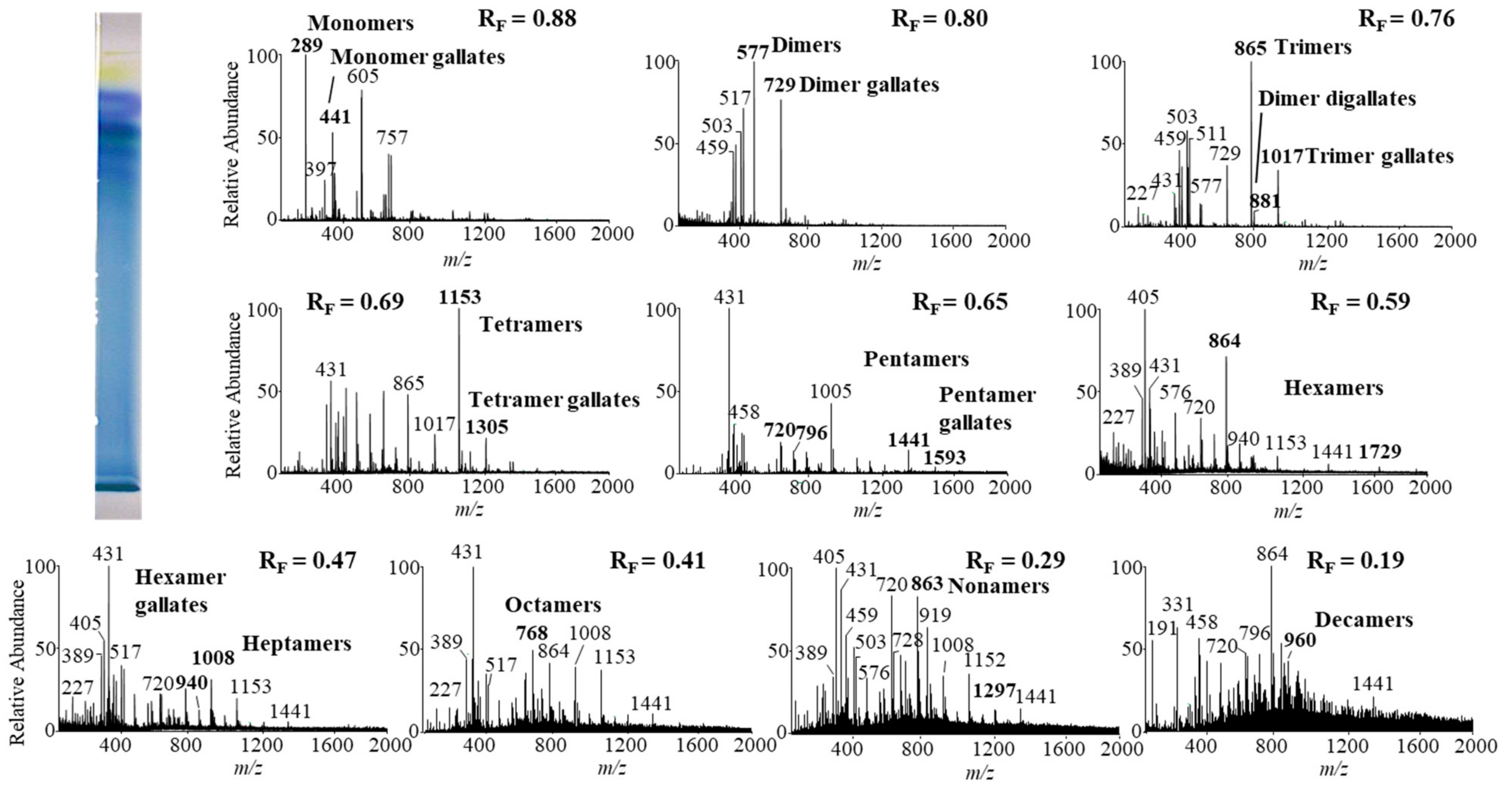
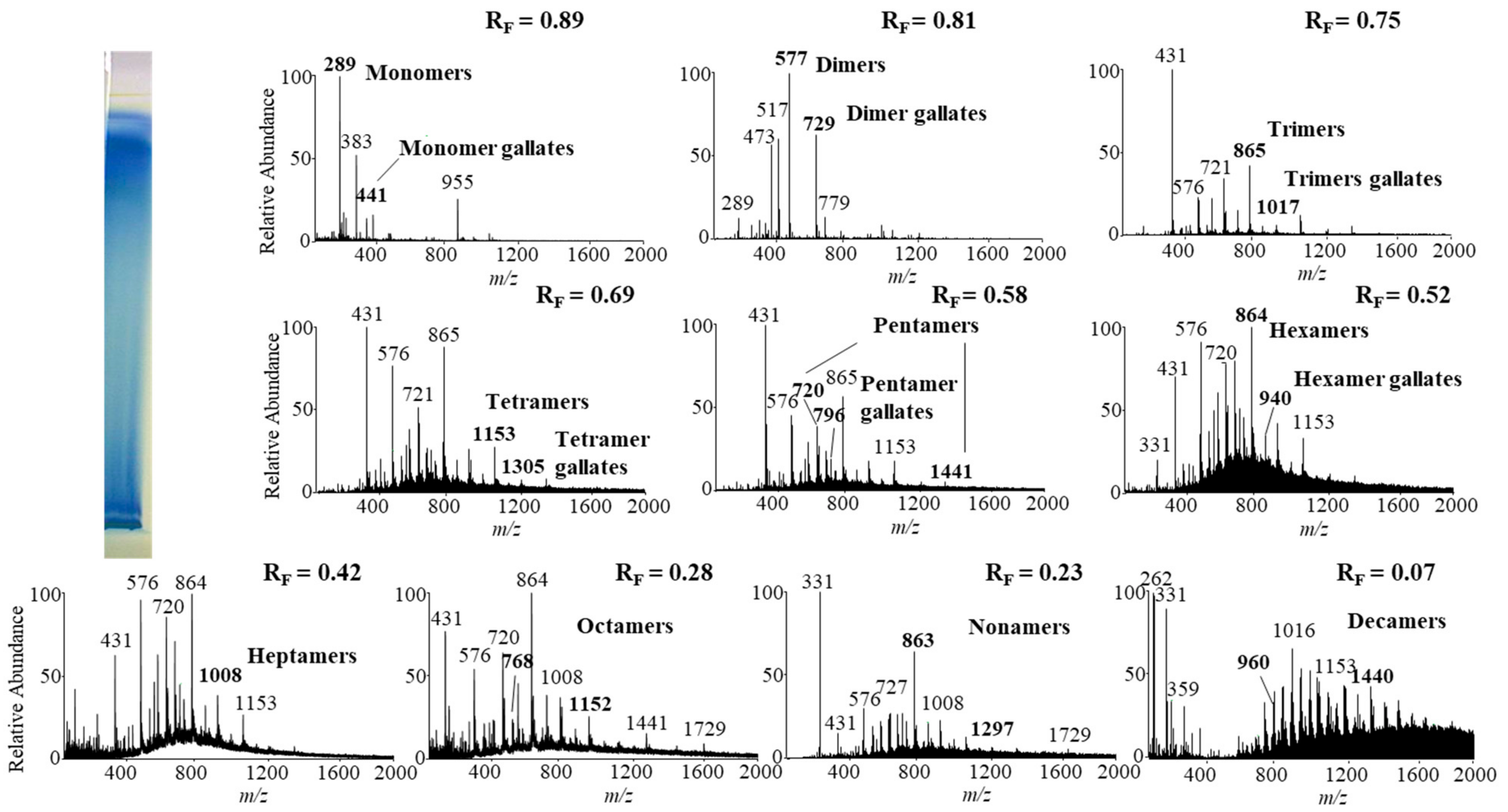
Monomers, monomer gallates, dimers and trimers were identified by comparing the fragmentation patterns with those of their respective commercial standards (−)-epicatechin, (−)-epicatechin gallate, procyanidin B2 and procyanidin C1. Standards of oligomers with higher degrees of polymerization (tetramers up to decamers), their gal lates and digallates are not commercially available. Therefore, tetramers to decamers, as well as their gallates and digallates, were tentatively identified based on their MS spectra (
Figure 8 and
Figure 9,
Table 3) and comparisons of the obtained and published frag mentation patterns. Pentamers to decamers and their gallates were identified as double- or triple-charged deprotonated molecular ions due to the limitations of the mass range of the MS detector.
Table 3 compares data from this study with literature data about proanthocyanidins identified in rhizomes [
7,
8,
9,
10,
12,
13,
14] and leaves [
7,
8,
18] of Japanese [
7,
8,
9,
10,
12,
13,
14,
18], Bohemian [
12,
14,
18] and giant [
7,
8,
12,
13,
14,
18] knotweed by using HPTLC–MS/MS [
9,
10,
18] and HPLC–MS methods [
7,
8,
12,
13,
14]. In this study we identified all B-type pro anthocyanidins from monomers to decamers (monomers, dimers, trimers, tetramers, pentamers, hexamers, heptamers, octamers, nonamers and decamers), as well as some of their gal lates (monomer gallates, dimer gallates, dimer digallates, trimer gallates, te tramer gallates, pentamer gallates and hexamer gallates) in rhizomes of Bohemian (
Figure 8,
Table 3) and giant knotweed (
Figure 9,
Table 3). To the best of our knowledge we were the first to identify pentamer gallates, hexamers, hexamer gallates, nonamers and decamers in rhizomes of Bohemian and giant knotweed. All these proanthocyanidins and their gallates were previously reported in our studies of Japanese knotweed rhi zomes by using the same HPTLC–MS/MS method [
10] and by HPTLC–MS/MS method on HPTLC silica gel plates [
9]. Rhizomes of all three knotweeds have the same chemical profile of proanthocyanidins with respect to the degree of polymerization and also with respect to gallates (
Table 3). Rhizomes of all three knotweeds also have the same chemical profile of proanthocyanidins with respect to the degree of polymerization as was discovered for leaves of Japanese, Bohemian and giant knotweed in our previous study [
18] (
Table 3). Additionally, rhizomes of all three knotweeds and leaves of giant knotweed have the same chemical profile of proanthocyanidins with respect to gallates (
Table 3). Only a few gallates are present in the profiles of leaves of Japanese knotweed (monomer gallates and dimer gallates) and Bohemian knotweed (monomer gallates, di mer gallates and dimer digallates) (
Table 3). Within the individual species of the stud ied knotweeds, only giant knotweed rhizomes and leaves have the same chemical profile of proanthocyanidins with respect to the degree of polymerization and also with respect to gallates (
Table 3).
The investigations on proanthocyanidins in knotweeds were performed on Japanese knotweed rhizomes [
7,
8,
9,
10,
12,
13,
14] using HPLC–MS [
7,
8,
12,
13,
14] or HPTLC–MS [
9,
10] methods (
Table 3). Literature data about proanthocyanidins identified (by using mass spectrometry) in Bohemian knotweed rhizomes are rather scarce (
Table 3). There are only two publications. One of the publications provides data only about monomers and monomer gallate [
12], the other in addition to monomers also reports dimers, dimer gal lates, dimer digallates, trimers, trimer gallates, trimer digallates, tetramer, tetramer gallates, pentamers, heptamers and octamers [
14]. Both publications [
12,
14] report the same proanthocyanidins and their gallates in giant knotweed. Additional publications reported monomers [
7,
8,
13] and monomer gallate [
7,
8,
13], dimers [
7,
8], dimer gallates [
13], dimer digallates [
13] in giant knotweed (
Table 3).













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}