
The Essential Oil Composition of Trachymene incisa Rudge subsp. incisa Rudge from Australia †

Abstract
:1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. Plant Material
3.2. Isolation of Volatile Oils
3.3. Identification of Components
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hart, J.M.; Henwood, M.J. A revision of Australian Trachymene (Apiaceae: Hydrocotyloideae). Aust. Syst. Bot. 2006, 19, 11–57. [Google Scholar] [CrossRef]
- Keighery, G.J.; Rye, B.L. A taxonomic revision of Trachymene sect. Dimetopia (Apiaceae). Nuytsia 1999, 13, 33–59. [Google Scholar]
- Powell, J.M. Apiaceae. In Flora of New South Wales; Harden, G.J., Ed.; New South Wales University Press: Sydney, Australia, 1992; Volume 3, pp. 87–116. [Google Scholar]
- Benson, D.; McDougall, L. Ecology of Sydney plant species: Part 1: Ferns, fern-allies. cycads, conifers and dicotyledon families Acanthaceae to Asclepiadaceae. Cunninghamia 1993, 3, 257–422. [Google Scholar]
- Davila, Y.C. Reproductive Ecology of Trachymene incisa (Apiaceae). Bachelor’s Thesis, The University of Sydney, Sydney, Australia, 1999. [Google Scholar]
- Watson, C. Root Demography of an Australian herb: Trachymene incisa. Bachelor’s Thesis, University of Sydney, Sydney, Australia, 2000. [Google Scholar]
- Gill, K. Seed Dispersal in Trachymene incisa (Apiaceae): The Process and Consequences for the Population. Bachelor’s Thesis, University of Sydney, Sydney, Australia, 2000. [Google Scholar]
- Davila, Y.C.; Wardle, G.M. Reproductive ecology of the Australian herb Trachymene incisa subsp. incisa (Apiaceae). Aust. J. Bot. 2002, 50, 619–626. [Google Scholar] [CrossRef] [Green Version]
- Wardle, G.M. Experimental determination of seed emergence and carry-over in the soil seed bank of the herbaceous perennial, Trachymene incisa (Apiaceae). Austral Ecol. 2003, 28, 161–172. [Google Scholar] [CrossRef]
- Edgar, G.; Ropert, J. The toxicity of Didiscus glaucifolius, ‘wild parsnip’, for sheep. Aust. Vet. J. 1942, 18, 125–126. [Google Scholar] [CrossRef]
- Clark, L.; Carlisle, C.H.; Beasley, P.S. Observations on the pathology of bent leg of lambs in south-western Queensland. Aust. Vet. J. 1975, 51, 4–10. [Google Scholar] [CrossRef]
- Clark, L. Trachymene spp and inferitility in sheep. Aust. Vet. J. 1977, 53, 249–250. [Google Scholar] [CrossRef] [PubMed]
- Philbey, A.W. Trachymene glaucifolia associated with bentleg in lambs. Aust. Vet. J. 1990, 67, 468. [Google Scholar] [CrossRef] [PubMed]
- Bourke, C.A. The clinical-differentiation of nervous and muscular locomotor disorders of sheep in Australia. Aust. Vet. J. 1995, 72, 228–234. [Google Scholar] [CrossRef]
- Alamgir, A.N.M. Medicinal, Non-medicinal, Biopesticides, Color- and Dye-Yielding Plants; Secondary Metabolites and Drug Principles; Significance of Medicinal Plants; Use of Medicinal Plants in the Systems of Traditional and Complementary and Alternative Medicines (CAMs). In Therapeutic Use of Medicinal Plants and Their Extracts; Progress in Drug Research; Springer: Cham, Switzerland, 2017; Volume 73, pp. 61–104. [Google Scholar] [CrossRef]
- Staples, T.L.; Dwyer, J.M.; Loy, X.; Mayfield, M.M. Potential mechanisms of coexistence in closely related forbs. Oikos 2016, 125, 1812–1823. [Google Scholar] [CrossRef]
- Wick, R.L.; Rane, K.K.; Sutton, D.P. 2 New ornamental hosts for Phytophthora cactorum—Trachymene caerulea and Gypsophila paniculata. Plant Dis. 1987, 71, 281. [Google Scholar] [CrossRef]
- Crone, M.; McComb, J.A.; O’Brien, P.A.; Hardy, G.E.S.J. Annual and herbaceous perennial native Australian plant species are symptomless hosts of Phytophthora cinnamomi in the Eucalyptus marginata (jarrah) forest of Western Australia. Plant Pathol. 2013, 62, 1057–1062. [Google Scholar] [CrossRef]
- Crone, M.; McComb, J.A.; O’Brien, P.A.; Hardy, G.E.S.J. Assessment of Australian native annual/herbaceous perennial plant species as asymptomatic or symptomatic hosts of Phytophthora cinnamomi under controlled conditions. For. Pathol. 2013, 43, 245–251. [Google Scholar] [CrossRef]
- Crone, M.; McComb, J.A.; O’Brien, P.A.; Hardy, G.E.S.J. Survival of Phytophthora cinnamomi as oospores, stromata, and thick-walled chlamydospores in roots of symptomatic and asymptomatic annual and herbaceous perennial plant species. Fungal Biol. 2013, 117, 112–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audran, J.C. Degeneration of Trachymene-pilosa exine by osmium-tetroxide used in impregnation technique. Planta 1981, 152, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Ball, E.A. Regeneration from isolated portions of the shoot apex of Trachymene coerulea RC Grah. Ann. Bot. 1980, 45, 103–112. [Google Scholar] [CrossRef]
- Batianoff, G.N. Coral cay terrestrial flora changes at Lady Elliot Island, Great Barrier Reef, Australia. Proc. R. Soc. Qld. 1998, 107, 5–14. [Google Scholar]
- Batianoff, G.N.; Naylor, G.C.; Old, J.A.; Neldner, V.J. Climate and vegetation changes at coringa-herald national nature reserve, coral Sea islands, Australia. Pac. Sci. 2010, 64, 73–92. [Google Scholar] [CrossRef] [Green Version]
- New South Wales Scientific Committee. Agnes BanksWoodland in the Sydney Basin Bioregion—Endangered ecological community Listing. New South Wales: National Parks and Wildlife Service. 2000. Available online: https://www.environment.nsw.gov.au/determinations/AgnesBanksWoodlandSydneyEndComListing.htm (accessed on 14 February 2021).
- Davila, Y.C.; Wardle, G.M. Bee boys and fly girls: Do pollinators prefer male or female umbels in protandrous parsnip, Trachymene incisa (Apiaceae)? Austral Ecol. 2007, 32, 798–807. [Google Scholar] [CrossRef]
- Davila, Y.C.; Wardle, G.M. Variation in native pollinators in the absence of honeybees: Implications for reproductive success of an Australian generalist-pollinated herb Trachymene incisa (Apiaceae). Bot. J. Linn. Soc. 2008, 156, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Peter, K.V. Handbook of Herbs and Spices Vol. 2; Woodhead Publishing: Abington, UK, 2004. [Google Scholar]
- Chizzola, R. Essential Oil Composition of Wild Growing Apiaceae from Europe and the Mediterranean. Nat. Prod. Commun. 2010, 5, 1477–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habtemariam, S. Chapter 6—Introduction to plant secondary metabolites—From biosynthesis to chemistry and antidiabetic action. In Medicinal Foods as Potential Therapies for Type-2 Diabetes and Associated Diseases; Habtemariam, S., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 109–132. [Google Scholar]
- Hampel, D.; Mosandl, A.; Wüst, M. Biosynthesis of mono- and sesquiterpenes in carrot roots and leaves (Daucus carota L.): Metabolic cross talk of cytosolic mevalonate and plastidial methylerythritol phosphate pathways. Phytochemistry 2005, 66, 305–311. [Google Scholar] [CrossRef]
- Palá-Paúl, J.; Brophy, J.J.; Goldsack, R.J.; Copeland, L.M.; Pérez-Alonso, M.J.; Velasco-Negueruela, A. Essential oil composition of the seasonal heterophyllous leaves of Eryngium vesiculosum from Australia. Aust. J. Bot. 2003, 51, 497–501. [Google Scholar] [CrossRef]
- Palá-Paúl, J.; Pérez-Alonso, M.J.; Velasco-Negueruela, A.; Varadé, J.; Villa, A.M.; Sanz, J.; Brophy, J.J. Essential oil composition of the different parts of Eryngium bourgatii Gouan from Spain. J. Chromatogr. A 2005, 1074, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Palá-Paúl, J.; Pérez-Alonso, M.J.; Velasco-Negueruela, A.; Varadé, J.; Villa, A.M.; Sanz, J.; Brophy, J.J. Analysis of the essential oil composition of the different parts of Eryngium glaciale Boiss. from Spain. J. Chromatogr. A 2005, 1094, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Ballhorn, D.J.; Godschalx, A.L.; Smart, S.M.; Kautz, S.; Schadler, M. Chemical defense lowers plant competitiveness. Oecologia 2014, 176, 811–824. [Google Scholar] [CrossRef]
- Fernandez, C.; Monnier, Y.; Santonja, M.; Gallet, C.; Weston, L.A.; Prévosto, B.; Saunier, A.; Baldy, V.; Bousquet-Mélou, A. The impact of competition and allelopathy on the trade-off between plant defense and growth in two contrasting tree species. Front. Plant Sci. 2016, 7, 594. [Google Scholar] [CrossRef] [Green Version]
- Mirmostafaee, S.; Azizi, M.; Fujii, Y. Study of allelopathic interaction of essential oils from medicinal and aromatic plants on seed germination and seedling growth of lettuce. Agronomy 2020, 10, 163. [Google Scholar] [CrossRef] [Green Version]
- Burns, J.H.; Strauss, S.Y. Effects of competition on phylogenetic signal and phenotypic plasticity in plant functional traits. Ecol. 2012, 93, 126–127. [Google Scholar] [CrossRef]
- Cabal, C.; Martínez-García, R.; de Castro Aguilar, A.; Valladares, F.; Pacala, S.W. The exploitative segregation of plant roots. Science 2020, 4, 1197–1199. [Google Scholar] [CrossRef] [PubMed]
- Brophy, J.J.; House, A.P.N.; Boland, D.J. Eucalyptus Leaf Oils-Use, Chemistry, Distillation and Marketing; Inkata Press: Melbourne, Auatralia; Sydney, Auatralia, 1991. [Google Scholar]
- Stenhagen, E.; Abrahamsson, S.; McLafferty, F.W. Registry of Mass Spectral Data; Wiley: New York, NY, USA, 1974. [Google Scholar]
- Heller, S.R.; Milne, G.W.A. EPA/NIH Mass Spectral Data Base; US Government Printing Office: Washington, DC, USA, 1978.
- Swigar, A.A.; Silverstein, R.M. Monoterpenes; Aldrich: Milwaukee, WI, USA, 1981. [Google Scholar]
- Adams, R.P. Identification of Essential oils Components by Gas Chromatography/Mass Spectroscopy; Allured Publishing Corporation: Carol Stream, IL, USA, 1995. [Google Scholar]
- Joulain, D.; König, W.A. The Atlas of Spectral Data of Sesquiterpene Hydrocarbons; E. B.-Verlag: Hamburg, Germany, 1998. [Google Scholar]
- Adams, R.P. Identification of Essential Oils Components by Gas Chromatography/Quadrupole Mass Spectroscopy; Allured Publishing Corporation: Carol Stream, IL, USA, 2001. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography—Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Babushok, V.I.; Linstrom, P.J.; Zenkevich, I.G. Retention Indices for Frequently Reported Compounds of Plant Essential Oils. J. Phys. Chem. Ref. Data 2011, 40, 043101–043147. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| KIa. | KI1 | KIb | KI2 | Compound | C. Type | T.in.I1 | T.in.I2 | T.in.I3 | T.in.II4 | T.in.II5 | I.M. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1007 | 1025 | 932 | 936 | α-pinene | MH | 7.2 | 19.2 | t | 0.1 | KI, MS | |
| 1034 | 1061 | 945 | 949 | α-fenchene | MH | 0.8 | KI, MS | ||||
| 1041 | 1068 | 946 | 950 | camphene | MH | t | KI, MS | ||||
| 1084 | 1110 | 974 | 978 | β-pinene | MH | 0.4 | 2.2 | t | 0.1 | KI, MS | |
| 1105 | 1122 | 969 | 973 | sabinene | MH | KI, MS | |||||
| 1127 | 1146 | 1008 | 1011 | δ-3-carene | MH | 1.8 | KI, MS | ||||
| 1139 | 1167 | 1002 | 1004 | α-phellandrene | MH | 1.2 | KI, MS | ||||
| 1141 | 1160 | 988 | 989 | myrcene | MH | t | KI, MS | ||||
| 1157 | 1177 | 1014 | 1017 | α-terpinene | MH | KI, MS | |||||
| 1176 | 1198 | 1024 | 1029 | limonene | MH | t | 0.8 | KI, MS | |||
| 1178 | 1209 | 1025 | 1030 | β-phellandrene | MH | t | KI, MS | ||||
| 1181 | 1211 | 1026 | 1031 | 1,8-cineole | OM | t | 9.0 | KI, MS | |||
| 1218 | 1234 | 1032 | 1038 | Z-β-ocimene | MH | t | KI, MS | ||||
| 1224 | 1245 | 1054 | 1059 | γ-terpinene | MH | t | KI, MS | ||||
| 1226 | 1250 | 1044 | 1048 | E-β-ocimene | MH | t | KI, MS | ||||
| 1249 | 1270 | 1020 | 1024 | p-cymene | MH | t | t | 0.6 | t | t | KI, MS |
| 1260 | 1282 | 1086 | 1086 | terpinolene | MH | t | 0.4 | 0.9 | KI, MS | ||
| 1320 | 1237 | 989 | 986 | 6-methylhept-5-en-2-one | O | t | 0.2 | t | t | KI, MS | |
| 1459 | 1469 | 1335 | 1337 | δ-elemene | SH | 0.3 | 0.3 | t | KI, MS | ||
| 1462 | 1491 | 1374 | 1376 | α-copaene | SH | t | 3.4 | t | KI, MS | ||
| 1510 | 1541 | 1387 | 1387 | β-cubebene | SH | 0.6 | t | KI, MS | |||
| 1523 | 1559 | 1411 | 1414 | α-cis-bergamotene | SH | 0.6 | 0.4 | KI, MS | |||
| 1543 | 1575 | 1432 | 1435 | α-trans-bergamotene | SH | 0.2 | 2.1 | t | KI, MS | ||
| 1560 | 1590 | 1389 | 1390 | β-elemene | SH | 0.8 | 2.9 | 0.4 | t | KI, MS | |
| 1563 | 1598 | 1417 | 1420 | β-caryophyllene | SH | 2.5 | 5.4 | 1.2 | 10.8 | 10.4 | KI, MS |
| 1570 | 1620 | 1439 | 1441 | aromadendrene | SH | 1.5 | 0.6 | 0.5 | 5.6 | 5.0 | KI, MS |
| 1572 | 1629 | 1509 | 1504 | α-bulnesene | SH | 0.5 | t | 1.2 | 0.9 | KI, MS | |
| 1632 | 1649 | 1458 | 1460 | allo-aromadendrene | SH | 0.2 | 0.4 | 0.4 | 0.4 | 0.4 | KI, MS |
| 1644 | 1664 | 1454 | 1456 | (E)-β-farnesene | SH | 0.3 | 1.4 | 0.6 | KI, MS | ||
| 1653 | 1666 | 1452 | 1453 | α-humulene | SH | 0.5 | 1.5 | 0.3 | 1.8 | 1.8 | KI, MS |
| 1658 | C15H24 | SH | 0.4 | 0.3 | 0.7 | KI, MS | |||||
| 1661 | 1696 | 1496 | 1492 | viridiflorene | SH | 1.9 | 0.8 | 0.6 | 5.4 | 4.7 | KI, MS |
| 1670 | 1708 | 1484 | 1480 | germacrene-D | SH | 1.4 | 2.4 | 1.5 | 0.5 | KI, MS | |
| 1680 | 1705 | 1505 | 1512 | γ-bisabolene | SH | 27.4 | KI, MS | ||||
| 1697 | 1717 | 1489 | 1486 | β-selinene | SH | 36.7 | 11.9 | 0.5 | 1.8 | KI, MS | |
| 1706 | 1734 | 1500 | 1494 | bicyclogermacrene | SH | 28.0 | 21.5 | 7.5 | 34.7 | 24.4 | KI, MS |
| 1722 | 1763 | 1513 | 1513 | γ-cadinene | SH | t | KI, MS | ||||
| 1724 | 1755 | 1522 | 1521 | δ-cadinene | SH | t | 9.0 | 0.5 | 1.5 | 1.7 | KI, MS |
| 1727 | 1788 | 1495 | 1531 | cis-cadina-1,4-diene | SH | 0.3 | t | KI, MS | |||
| 1734 | 1744 | 1505 | 1504 | (E,E)-α-farnesene | SH | t | 1.4 | 2.6 | 1.6 | KI, MS | |
| 1749 | 1773 | 1479 | 1482 | ar-curcumene | SH | t | t | 0.7 | 1.1 | KI, MS | |
| 1886 | 1986 | 1582 | 1580 | caryophyllene oxide | OS | 1.6 | 2.3 | 0.2 | 1.0 | 12.8 | KI, MS |
| 1958 | 2039 | 1602 | 1582 | ledol | OS | 0.2 | 0.2 | 0.1 | 0.5 | t | KI, MS |
| 2016 | 2067 | 1645 | 1636 | cubenol | OS | t | 0.9 | 0.2 | 0.8 | 1.5 | KI, MS |
| 2018 | 2074 | 1595 | 1588 | cubeban-11-ol | OS | 0.7 | 0.4 | 0.4 | 1.6 | 1.1 | KI, MS |
| 2034 | 2082 | 1590 | 1582 | globulol | OS | 2.0 | 1.5 | 1.3 | 7.0 | 6.9 | KI, MS |
| 2044 | 2090 | 1592 | 1591 | viridiflorol | OS | 1.5 | 1.1 | 0.8 | 3.9 | 3.5 | KI, MS |
| 2066 | C15H26O | OS | 0.5 | 0.3 | 0.3 | 2.2 | 2.0 | KI, MS | |||
| 2074 | C15H26O | OS | 0.4 | 0.6 | 0.3 | 1.3 | 1.0 | KI, MS | |||
| 2078 | C15H26O | OS | 0.2 | 0.3 | KI, MS | ||||||
| 2086 | 2127 | 1577 | 1576 | spathulenol | OS | 4.2 | 11.2 | 2.9 | 6.7 | 7.5 | KI, MS |
| Total | 93.5 | 81.7 | 91.0 | 91.1 | 88.4 | ||||||
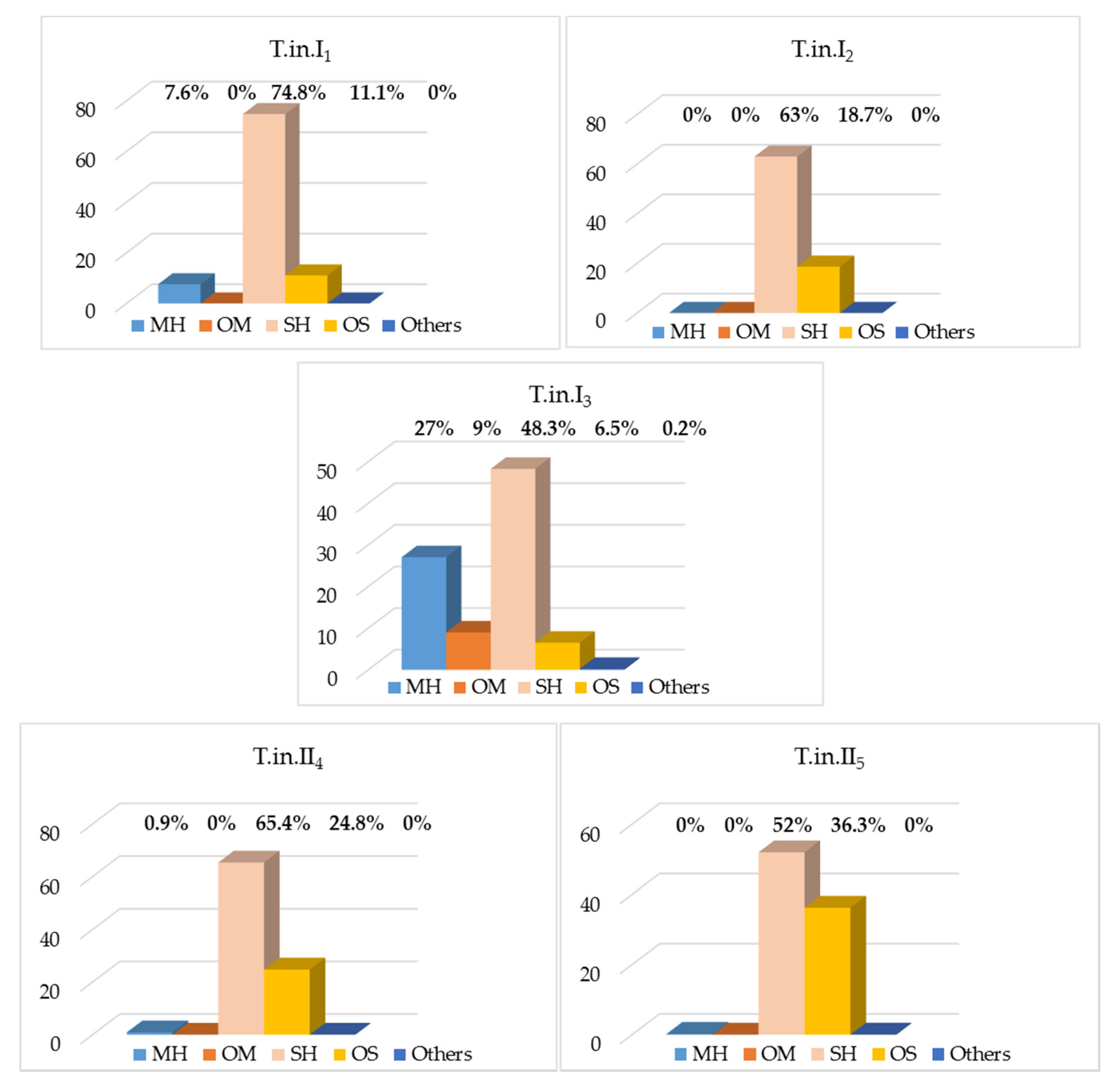
| Monoterpene Hydrocarbon (MH) | 16 | 7.6 | 0 | 27.0 | 0.9 | 0 | |||||
| Oxygenated Monoterpene (OM) | 1 | 0 | 0 | 9.0 | 0 | 0 | |||||
| Sesquiterpene Hydrocarbon (SH) | 23 | 74.8 | 63.0 | 48.3 | 65.4 | 52.0 | |||||
| Oxygenated Sesquiterpene (OS) | 10 | 11.1 | 18.7 | 6.5 | 24.8 | 36.3 | |||||
| Other (O) | 1 | 0 | 0 | 0.2 | 0 | 0 | |||||
| Total | 51.0 | 93.5 | 81.7 | 91.0 | 91.1 | 88.2 | |||||
| Sample | Voucher n° | Location | Yield |
|---|---|---|---|
| T.in.I1 | LMC4175 | Single National Park: (NSW, Australia) 20 km NW of Guyra (24 March 2007) | 0.05% |
| T.in.I2 | LMC4176 | 0.17% | |
| T.in.I3 | LMC4211 | 0.05% | |
| T.in.II4 | LMC4210 | 0.05% | |
| T.in.II5 | LMC4234 | Dumaresq Dam: (NSW, Australia) Along southwestern side of Dumaresq Dam, c. 10 km NW of Armidale (14 April 2007) | 0.1% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palá-Paúl, J.; Copeland, L.M.; Brophy, J.J. The Essential Oil Composition of Trachymene incisa Rudge subsp. incisa Rudge from Australia . Plants 2021, 10, 601. https://doi.org/10.3390/plants10030601
Palá-Paúl J, Copeland LM, Brophy JJ. The Essential Oil Composition of Trachymene incisa Rudge subsp. incisa Rudge from Australia . Plants. 2021; 10(3):601. https://doi.org/10.3390/plants10030601
Chicago/Turabian StylePalá-Paúl, Jesús, Lachlan M. Copeland, and Joseph J. Brophy. 2021. "The Essential Oil Composition of Trachymene incisa Rudge subsp. incisa Rudge from Australia " Plants 10, no. 3: 601. https://doi.org/10.3390/plants10030601
APA StylePalá-Paúl, J., Copeland, L. M., & Brophy, J. J. (2021). The Essential Oil Composition of Trachymene incisa Rudge subsp. incisa Rudge from Australia . Plants, 10(3), 601. https://doi.org/10.3390/plants10030601