
A Root Tip-Specific Expressing Anthocyanin Marker for Direct Identification of Transgenic Tissues by the Naked Eye in Symbiotic Studies



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Anthocyanin Markers Allow the Easy Visual Screening of Transgenic Roots
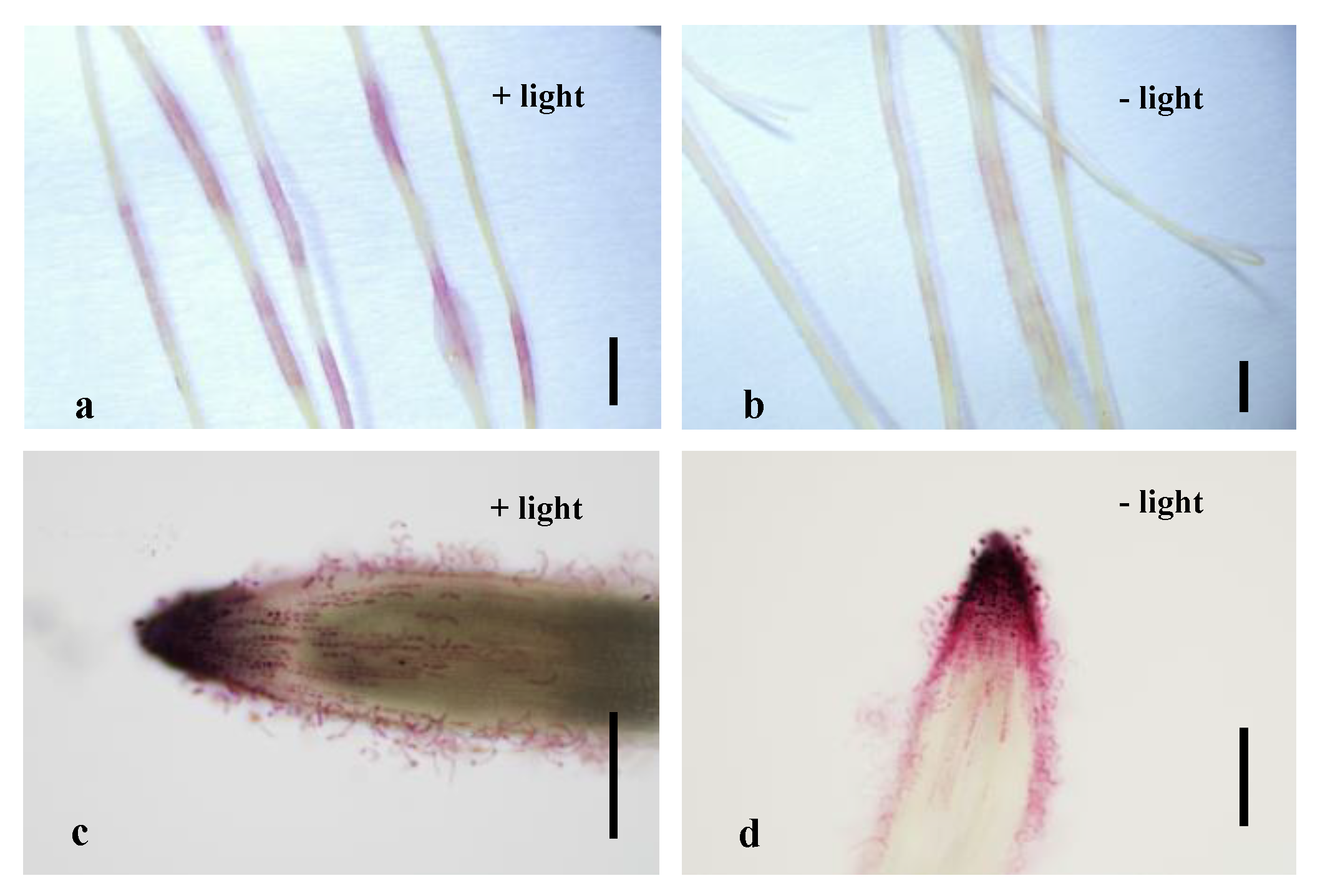
2.2. Light and Sucrose Induce Endogenous Anthocyanin Production in Roots but Do Not Affect Visual Screening of Transgenic Roots Expressing pMtRC-MtLAP1
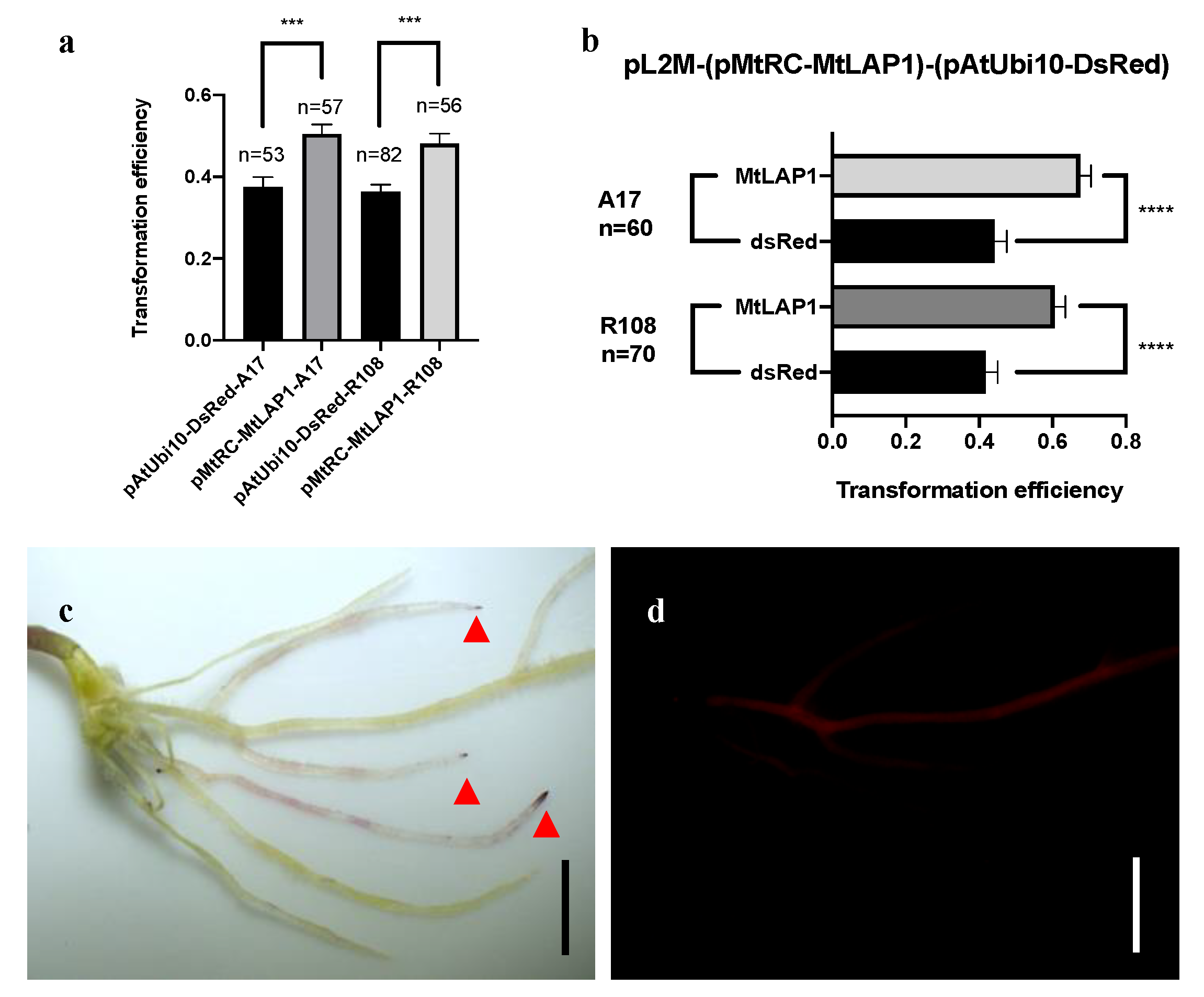
2.3. The Apparent Transformation Efficiency of pMtRC-MtLAP1 Is Higher than pAtUbi10-DsRed
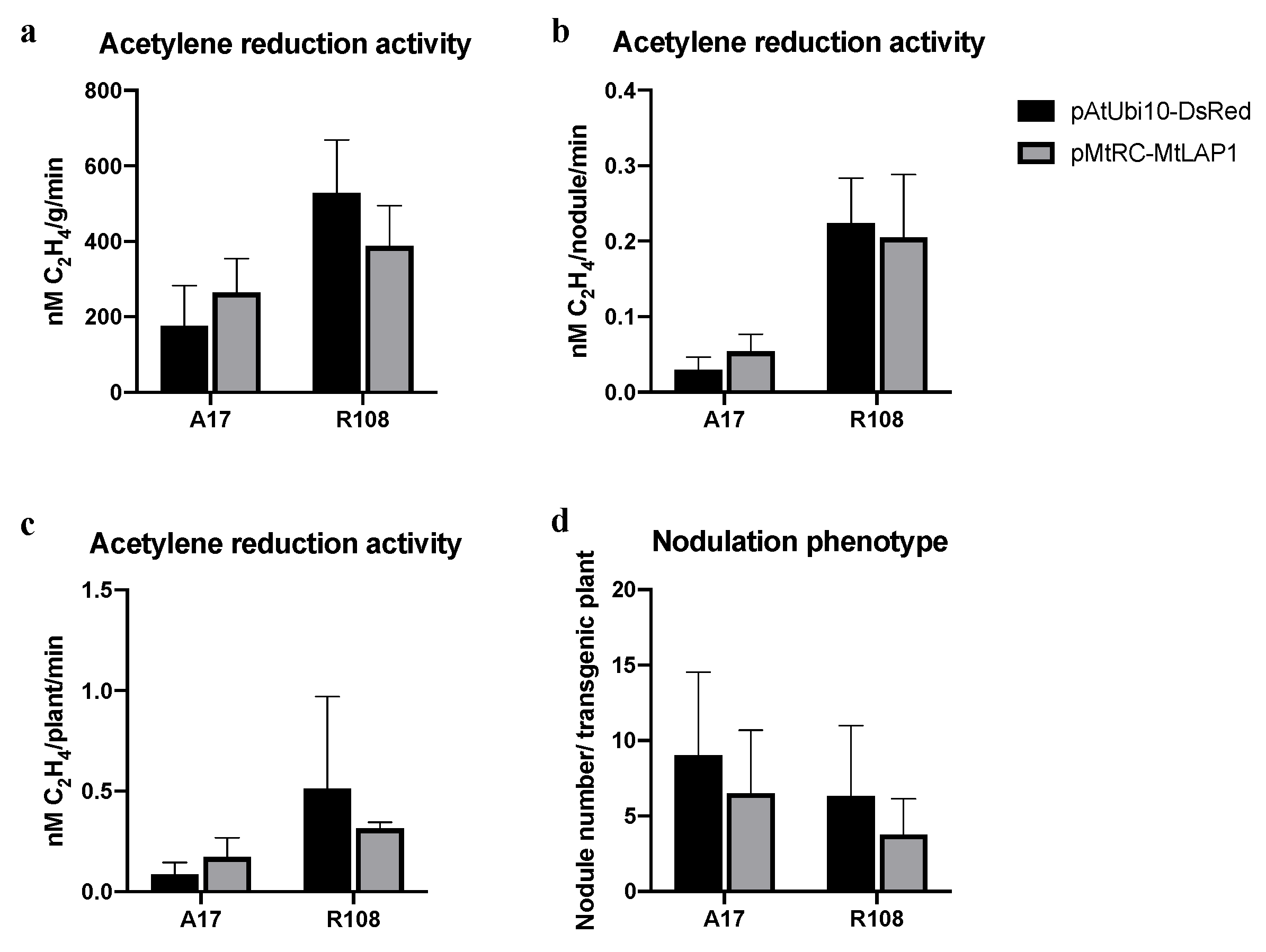
2.4. The MtLAP1 Screening Marker Does Not Affect Nodulation
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain, Plant Materials and Growth Conditions
4.2. pAtE47, pMtRC, and pMtLAP1 Vector Construction
4.3. A. Rhizogenes-Mediated Hairy Root Transformation
4.4. The Acetylene Reduction Assay
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Oldroyd, G.E. Speak, friend, and enter: Signalling systems that promote beneficial symbiotic associations in plants. Nat. Rev. Microbiol. 2013, 11, 252–263. [Google Scholar] [CrossRef]
- Roy, S.; Liu, W.; Nandety, R.S.; Crook, A.; Mysore, K.S.; Pislariu, C.I.; Frugoli, J.; Dickstein, R.; Udvardi, M.K. Celebrating 20 Years of Genetic Discoveries in Legume Nodulation and Symbiotic Nitrogen Fixation. Plant Cell 2020, 32, 15–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burks, D.; Azad, R.; Wen, J.; Dickstein, R. The Medicago truncatula Genome: Genomic Data Availability. Methods Mol. Biol. 2018, 1822, 39–59. [Google Scholar] [CrossRef] [PubMed]
- Proust, H.; Hartmann, C.; Crespi, M.; Lelandais-Brière, C. Root Development in Medicago truncatula: Lessons from Genetics to Functional Genomics. In Functional Genomics in Medicago truncatula: Methods and Protocols; Cañas, L.A., Beltrán, J.P., Eds.; Springer: New York, NY, USA, 2018; pp. 205–239. [Google Scholar]
- de Bang, T.C.; Lundquist, P.K.; Dai, X.; Boschiero, C.; Zhuang, Z.; Pant, P.; Torres-Jerez, I.; Roy, S.; Nogales, J.; Veerappan, V.; et al. Genome-Wide Identification of Medicago Peptides Involved in Macronutrient Responses and Nodulation. Plant Physiol. 2017, 175, 1669–1689. [Google Scholar] [CrossRef] [Green Version]
- Boisson-Dernier, A.; Chabaud, M.; Garcia, F.; Bécard, G.; Rosenberg, C.; Barker, D.G. Agrobacterium rhizogenes-transformed roots of Medicago truncatula for the study of nitrogen-fixing and endomycorrhizal symbiotic associations. Mol. Plant Microbe Interact. 2001, 14, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Lin, M.-H.; Gresshoff, P.M.; Indrasumunar, A.; Ferguson, B.J. pHairyRed: A Novel Binary Vector Containing the DsRed2 Reporter Gene for Visual Selection of Transgenic Hairy Roots. Mol. Plant 2011, 4, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, T.; Kusakabe, M.; Shimoda, Y.; Sato, S.; Tabata, S.; Murooka, Y.; Hayashi, M. Polyubiquitin Promoter-Based Binary Vectors for Overexpression and Gene Silencing in Lotus japonicus. Mol. Plant Microbe Interact. 2008, 21, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Kondorosi, É.; Kereszt, A. An anthocyanin marker for direct visualization of plant transformation and its use to study nitrogen-fixing nodule development. J. Plant Res. 2019, 132, 695–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Wang, X.; Li, H.; Liu, S.; Jin, L.; Lyu, Y.; Shi, M.; Liu, S.; Yang, X.; Lyu, S. Anthocyanin, a novel and user-friendly reporter for convenient, non-destructive, low cost, directly visual selection of transgenic hairy roots in the study of rhizobia-legume symbiosis. Plant Methods 2020, 16, 94. [Google Scholar] [CrossRef]
- Peel, G.J.; Pang, Y.; Modolo, L.V.; Dixon, R.A. The LAP1 MYB transcription factor orchestrates anthocyanidin biosynthesis and glycosylation in Medicago. Plant J. 2009, 59, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Zhang, J.; Subramanian, S.; Stacey, G.; Yu, O. Flavones and flavonols play distinct critical roles during nodulation of Medicago truncatula by Sinorhizobium meliloti. Plant J. 2009, 57, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Solfanelli, C.; Poggi, A.; Loreti, E.; Alpi, A.; Perata, P. Sucrose-specific induction of the anthocyanin biosynthetic pathway in Arabidopsis. Plant Physiol. 2006, 140, 637–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, L.S.; Liu, A. Light signaling induces anthocyanin biosynthesis via AN3 mediated COP1 expression. Plant Signal. Behav. 2015, 10, e1001223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, E.; Engler, C.; Gruetzner, R.; Werner, S.; Marillonnet, S. A Modular Cloning System for Standardized Assembly of Multigene Constructs. PLoS ONE 2011, 6, e16765. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.W.; Breakspear, A.; Stacey, N.; Findlay, K.; Nakashima, J.; Ramakrishnan, K.; Liu, M.; Xie, F.; Endre, G.; de Carvalho-Niebel, F.; et al. A protein complex required for polar growth of rhizobial infection threads. Nat. Commun. 2019, 10, 2848. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.W.; Breakspear, A.; Guan, D.; Cerri, M.R.; Jackson, K.; Jiang, S.; Robson, F.; Radhakrishnan, G.V.; Roy, S.; Bone, C.; et al. NIN Acts as a Network Hub Controlling a Growth Module Required for Rhizobial Infection. Plant Physiol. 2019, 179, 1704–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bovy, A.; Schijlen, E.; Hall, R.D. Metabolic engineering of flavonoids in tomato (Solanum lycopersicum): The potential for metabolomics. Metabolomics 2007, 3, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, A.; Takeda, K.; Ohnishi, T. Light-Induced Anthocyanin Reduces the Extent of Damage to DNA in UV-Irradiated Centaurea cyanus Cells in Culture. Plant Cell Physiol. 1991, 32, 541–547. [Google Scholar] [CrossRef]
- Baskar, V.; Venkatesh, R.; Ramalingam, S. Flavonoids (Antioxidants Systems) in Higher Plants and Their Response to Stresses. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D.K., Palma, J.M., Corpas, F.J., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 253–268. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruan, Y.; Chen, K.; Su, Y.; Jiang, S.; Xu, P.; Murray, J.D. A Root Tip-Specific Expressing Anthocyanin Marker for Direct Identification of Transgenic Tissues by the Naked Eye in Symbiotic Studies. Plants 2021, 10, 605. https://doi.org/10.3390/plants10030605
Ruan Y, Chen K, Su Y, Jiang S, Xu P, Murray JD. A Root Tip-Specific Expressing Anthocyanin Marker for Direct Identification of Transgenic Tissues by the Naked Eye in Symbiotic Studies. Plants. 2021; 10(3):605. https://doi.org/10.3390/plants10030605
Chicago/Turabian StyleRuan, Yiting, Ke Chen, Yangyang Su, Suyu Jiang, Ping Xu, and Jeremy D. Murray. 2021. "A Root Tip-Specific Expressing Anthocyanin Marker for Direct Identification of Transgenic Tissues by the Naked Eye in Symbiotic Studies" Plants 10, no. 3: 605. https://doi.org/10.3390/plants10030605
APA StyleRuan, Y., Chen, K., Su, Y., Jiang, S., Xu, P., & Murray, J. D. (2021). A Root Tip-Specific Expressing Anthocyanin Marker for Direct Identification of Transgenic Tissues by the Naked Eye in Symbiotic Studies. Plants, 10(3), 605. https://doi.org/10.3390/plants10030605