
Differences in the Inhibitory Specificity Distinguish the Efficacy of Plant Protease Inhibitors on Mouse Fibrosarcoma



Abstract
:1. Introduction
2. Results
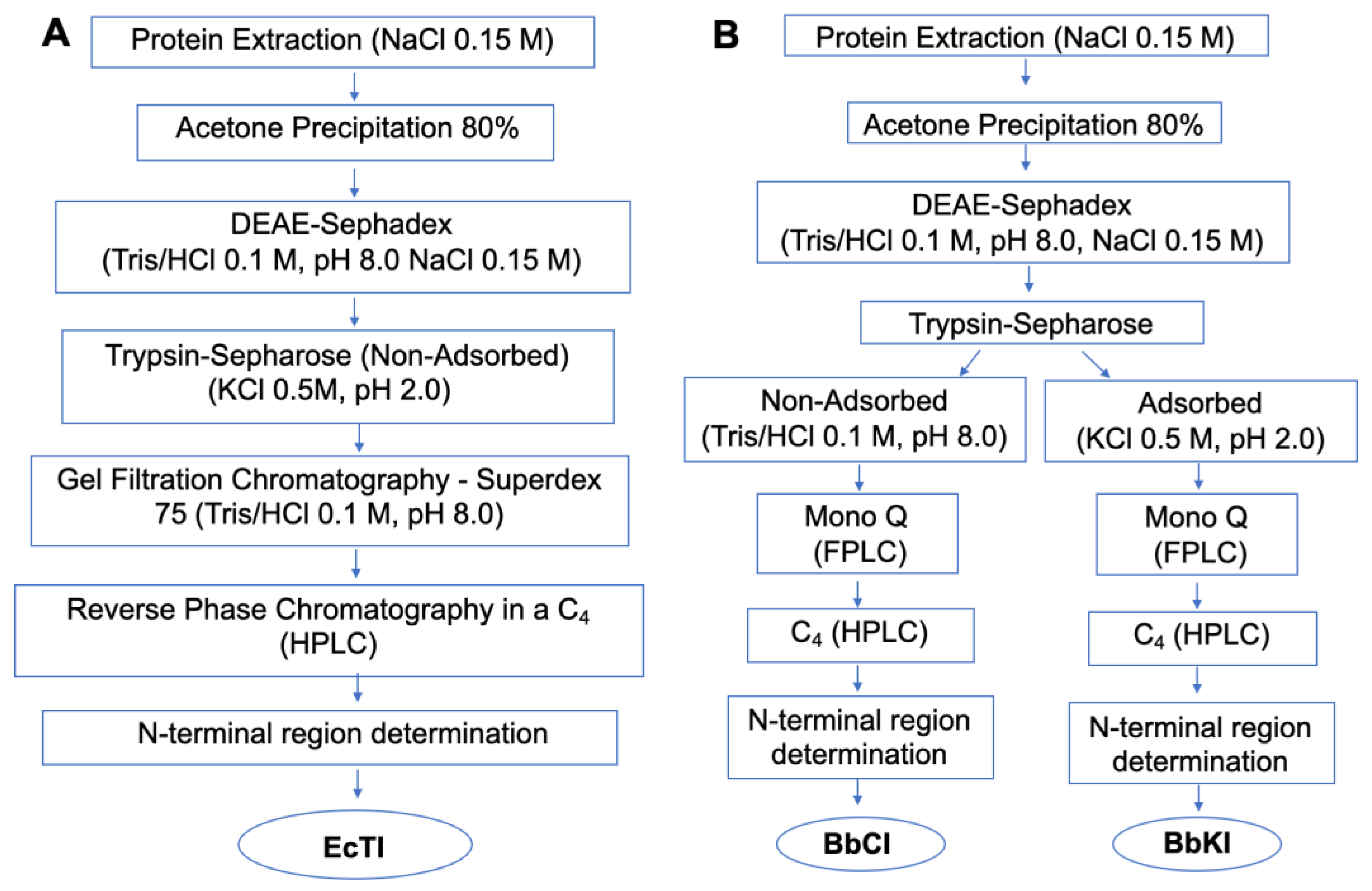
2.1. Inhibitor Purification and Characterization
2.2. Effects of rBbCI, rBbKI, and EcTI on the L929 Fibrosarcoma Lineage
2.2.1. Effects of Inhibitors on Cell Viability
2.2.2. Effects of Inhibitors on Cell Adhesion
2.2.3. Effects of Inhibitors on the Cell Cycle
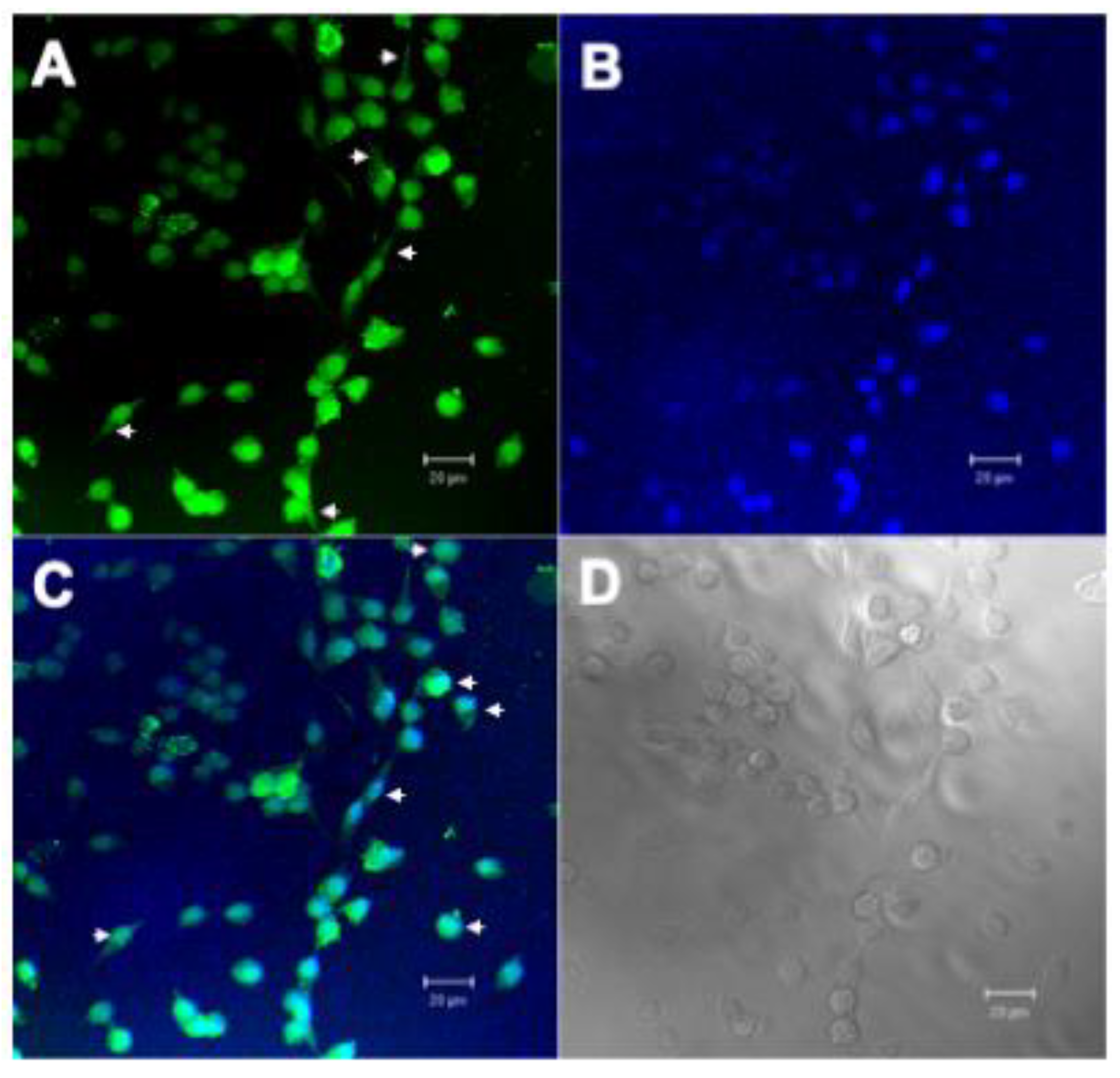
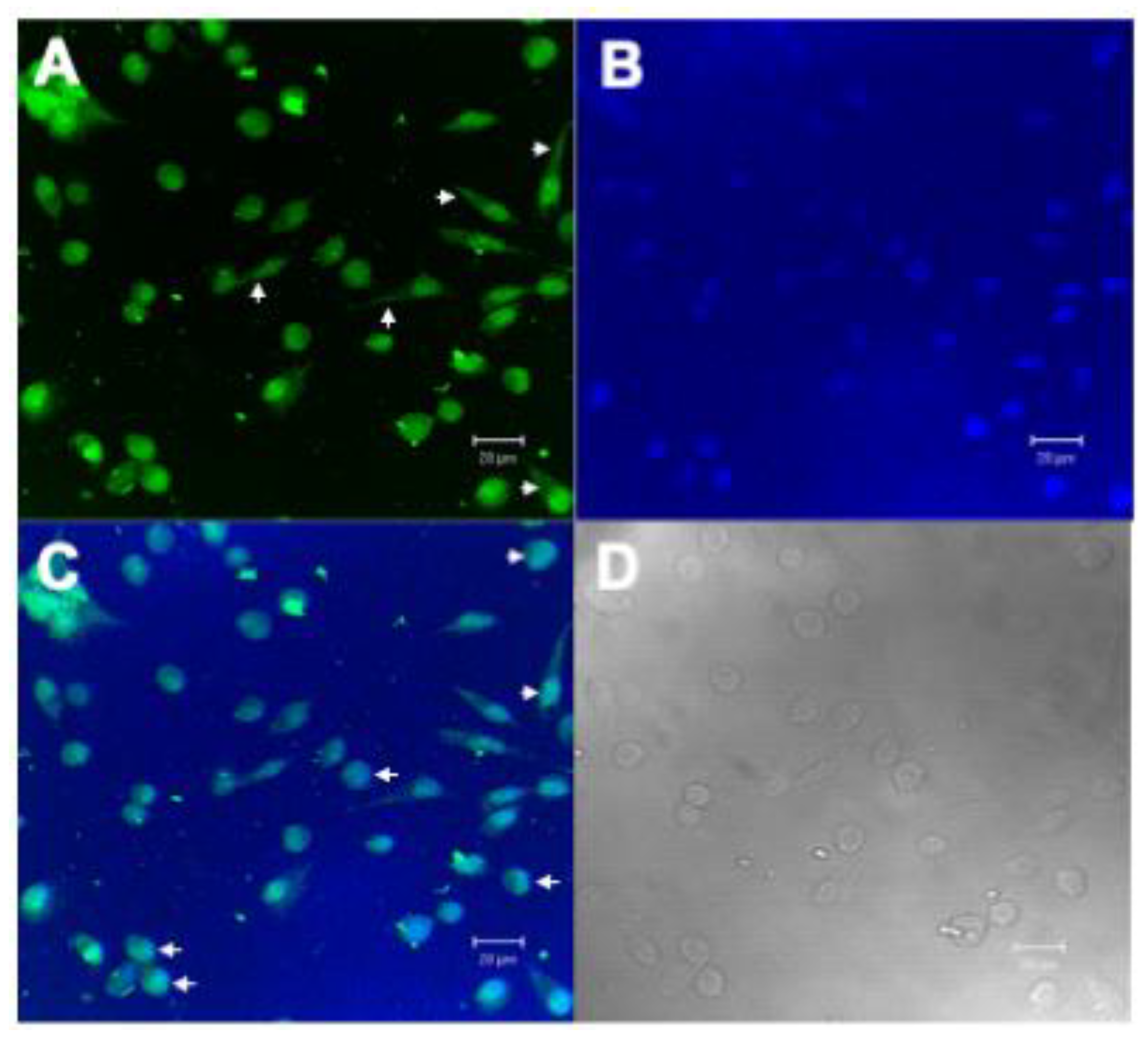
2.2.4. Confocal Microscopy
2.2.5. Effect of the BbKI-Derived Peptide on L929 Cell Viability
3. Discussion
4. Materials and Methods
4.1. Seeds
4.2. Purification of Native Inhibitors
4.3. Extraction of B. bauhinioides and E. contortisiliquum Inhibitors
4.4. Ion Exchange Chromatography on DEAE-Sephadex
4.5. Affinity Chromatography on Trypsin-Sepharose
4.6. Fast Protein Liquid Chromatography on Mono Q
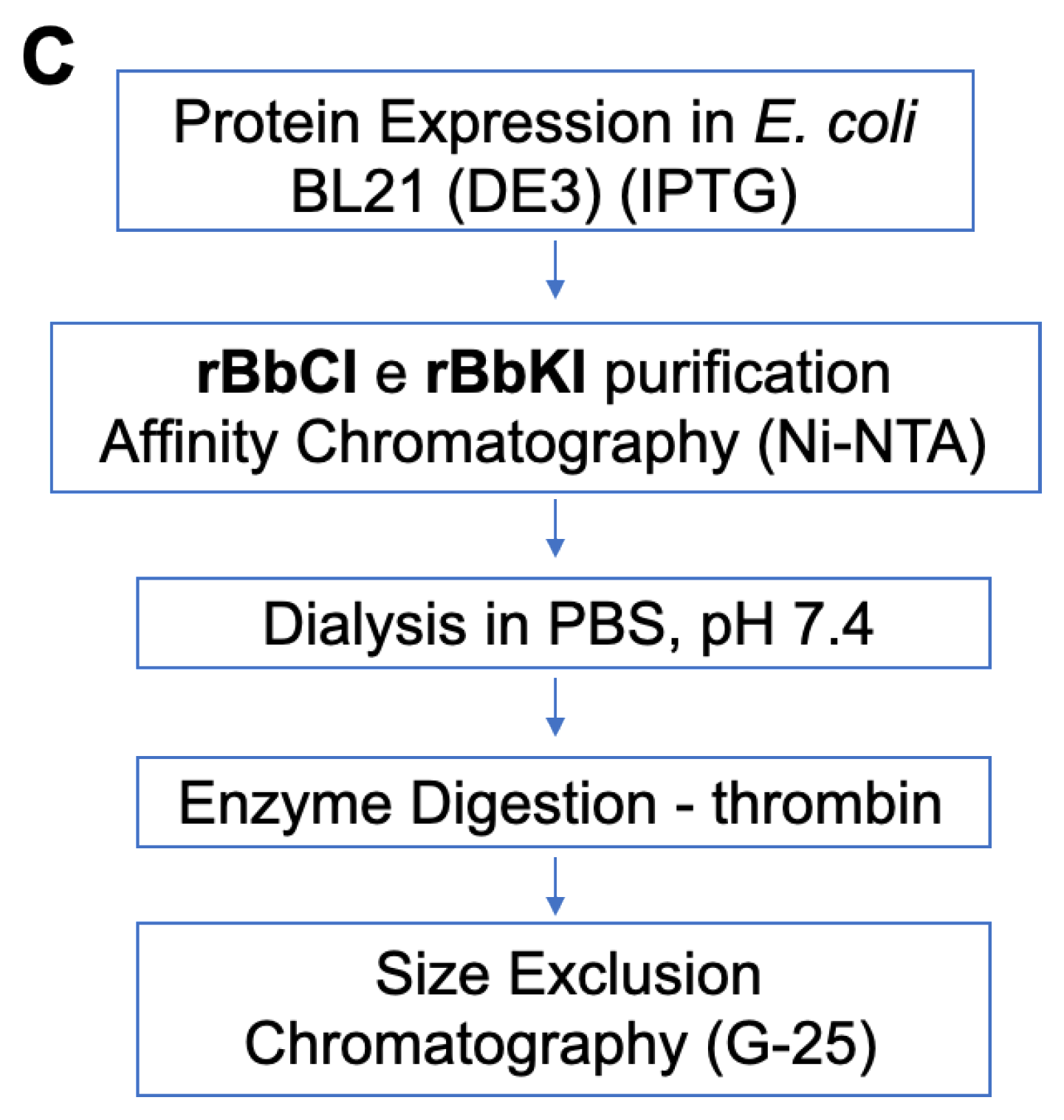
4.7. Recombinant rBbCI and rBbKI Inhibitors
4.7.1. Cloning
4.7.2. Expression of rBbCI and rBbKI Inhibitors
4.8. Purification of rBbKI and rBbCI Inhibitors
4.8.1. Affinity Chromatography and Cleavage of the Fusion Peptide
4.8.2. Size Exclusion Chromatography
4.8.3. Reverse-Phase Chromatography (HPLC System)
4.8.4. Enzymatic Assays
4.8.5. Hydrolysis of Substrates by Serine Proteases and Determination of Inhibitory Activity
4.9. Hydrolysis of Z-Phe-Arg-AMC by Cysteine Proteases and Determination of Inhibitory Activity
4.9.1. Determination of rBbKI and EcTI Concentrations
4.9.2. Determination of rBbCI Concentration
4.9.3. Synthetic Peptide RPGLPVRFESPL-NH2
4.9.4. Cultivation Conditions for the L929 Cell Line
4.9.5. Cell Viability
4.9.6. Cell Adhesion Assay
4.9.7. Cell Cycle Assay
4.10. Analysis of Cell Interaction with Inhibitors by Confocal Microscopy
4.10.1. Covalent Conjugation of Inhibitors to Fluorescent Dye
4.10.2. Cell Labeling for Confocal Microscopy
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Morrison, B.A. Soft Tissue Sarcomas of the Extremities. Bayl. Univ. Med Cent. Proc. 2003, 16, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Gore, M.R. Treatment, outcomes, and demographics in sinonasal sarcoma: A systematic review of the literature. BMC Ear Nose Throat Disord. 2018, 18, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folpe, A.L. Fibrosarcoma: A review and update. Histopathology 2013, 64, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, W.S. Advances in the adjuvant treatment of infantile fibrosarcoma. Expert Rev. Anticancer. Ther. 2003, 3, 185–191. [Google Scholar] [CrossRef]
- Hoefkens, F.; Dehandschutter, C.; Somville, J.; Meijnders, P.; Van Gestel, D. Soft tissue sarcoma of the extremities: Pending questions on surgery and radiotherapy. Radiat. Oncol. 2016, 11, 136. [Google Scholar] [CrossRef] [Green Version]
- Soofiyani, S.R.; Kazemi, T.; Lotfipour, F.; Hosseini, A.M.; Shanehbandi, D.; Hallaj-Nezhadi, S.; Baradaran, B. Gene therapy with IL-12 induced enhanced anti-tumor activity in fibrosarcoma mouse model. Artif. Cells Nanomed. Biotechnol. 2016, 44, 1988–1993. [Google Scholar] [CrossRef]
- FDA: U.S. Food and Drug Administration. Center for Drug Evaluation and Research. Available online: https://www.cancer.gov/about-cancer/treatment/drugs/soft-tissue-sarcoma (accessed on 2 March 2021).
- Liyanage, C.; Fernando, A.; Batra, J. Differential roles of protease isoforms in the tumor microenvironment. Cancer Metastasis Rev. 2019, 38, 389–415. [Google Scholar] [CrossRef]
- Turk, B.; Turk, D.; Turk, V. Protease signalling: The cutting edge. EMBO J. 2012, 31, 1630–1643. [Google Scholar] [CrossRef] [Green Version]
- Oliva, M.L.V.; Santomauro-Vaz, E.M.; Andrade, S.A.; Juliano, M.A.; Pott, V.J.; Sampaio, M.U.; Sampaio, C.A.M. Synthetic Peptides and Fluorogenic Substrates Related to the Reactive Site Sequence of Kunitz-Type Inhibitors Isolated from Bauhinia: Interaction with Human Plasma Kallikrein. Biol. Chem. 2001, 382, 109–113. [Google Scholar] [CrossRef]
- de Paula, C.A.A.; Coulson-Thomas, V.J.; Ferreira, J.G.; Maza, P.K.; Suzuki, E.; Nakahata, A.M.; Nader, H.B.; Sampaio, M.U.; Oliva, M.L.V. Enterolobium contortisiliquum Trypsin Inhibitor (EcTI), a Plant Proteinase Inhibitor, Decreases in Vitro Cell Adhesion and Invasion by Inhibition of Src Protein-Focal Adhesion Kinase (FAK) Signaling Pathways. J. Biol. Chem. 2012, 287, 170–182. [Google Scholar] [CrossRef] [Green Version]
- Nakahata, A.M.; Mayer, B.; Neth, P.; Hansen, D.; Sampaio, M.U.; Oliva, M.L.V. Blocking the Proliferation of Human Tumor Cell Lines by Peptidase Inhibitors from Bauhinia Seeds. Planta Medica 2013, 79, 227–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, M.V.; De Oliveira, C.; Salu, B.R.; Andrade, S.A.; Malloy, P.M.; Sato, A.C.; Vicente, C.P.; Sampaio, M.U.; Maffei, F.H.; Oliva, M.L.V. The Kallikrein Inhibitor from Bauhinia bauhinioides (BbKI) shows antithrombotic properties in venous and arterial thrombosis models. Thromb. Res. 2014, 133, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Martins-Olivera, B.T.; Almeida-Reis, R.; Theodoro-Júnior, O.A.; Oliva, L.V.; Nunes, N.N.D.S.; Olivo, C.R.; de Brito, M.V.; Prado, C.M.; Leick, E.A.; Martins, M.D.A.; et al. The Plant-DerivedBauhinia bauhinioidesKallikrein Proteinase Inhibitor (rBbKI) Attenuates Elastase-Induced Emphysema in Mice. Mediat. Inflamm. 2016, 2016, 5346574. [Google Scholar] [CrossRef] [Green Version]
- Lubkowski, J.; Durbin, S.V.; Silva, M.C.C.; Farnsworth, D.; Gildersleeve, J.C.; Oliva, M.L.V.; Wlodawer, A. Structural analysis and unique molecular recognition properties of a Bauhinia forficata lectin that inhibits cancer cell growth. FEBS J. 2016, 284, 429–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolozzo, A.S.S.; Rodrigues, A.P.D.; Arantes-Costa, F.M.; Saraiva-Romanholo, B.M.; De Souza, F.C.R.; Brüggemann, T.R.; De Brito, M.V.; Ferreira, R.D.S.; Correia, M.T.D.S.; Paiva, P.M.G.; et al. The Plant Proteinase Inhibitor CrataBL Plays a Role in Controlling Asthma Response in Mice. BioMed Res. Int. 2018, 2018, 9274817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, C.; Santana, L.; Carmona, A.; Cezari, M.; Sampaio, M.; Sampaio, C.; Oliva, M. Structure of Cruzipain/Cruzain Inhibitors Isolated from Bauhinia bauhinioides Seeds. Biol. Chem. 2001, 382, 847–852. [Google Scholar] [CrossRef]
- Ferreira, J.G.; Diniz, P.M.M.; de Paula, C.A.A.; Lobo, Y.A.; Paredes-Gamero, E.J.; Paschoalin, T.; Nogueira-Pedro, A.; Maza, P.K.; Toledo, M.S.; Suzuki, E.; et al. The Impaired Viability of Prostate Cancer Cell Lines by the Recombinant Plant Kallikrein Inhibitor. J. Biol. Chem. 2013, 288, 13641–13654. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, R.; Brito, M.; Napoleão, T.; Silva, M.; Paiva, P.; Oliva, M. Effects of two protease inhibitors from Bauhinia bauhinoides with different specificity towards gut enzymes of Nasutitermes corniger and its survival. Chemosphere 2019, 222, 364–370. [Google Scholar] [CrossRef]
- Lobo, Y.A.; Bonazza, C.; Batista, F.P.; Castro, R.A.; Bonturi, C.R.; Salu, B.R.; Sinigaglia, R.D.C.; Toma, L.; Vicente, C.M.; Pidde, G.; et al. EcTI impairs survival and proliferation pathways in triple-negative breast cancer by modulating cell-glycosaminoglycans and inflammatory cytokines. Cancer Lett. 2020, 491, 108–120. [Google Scholar] [CrossRef]
- Bonturi, C.R.; Motaln, H.; Silva, M.C.C.; Salu, B.R.; De Brito, M.V.; Costa, L.D.A.L.; Torquato, H.F.V.; Nunes, N.N.D.S.; Paredes-Gamero, E.J.; Turnšek, T.L.; et al. Could a plant derived protein potentiate the anticancer effects of a stem cell in brain cancer? Oncotarget 2018, 9, 21296–21312. [Google Scholar] [CrossRef] [Green Version]
- Bonturi, C.R.; Silva, M.C.C.; Motaln, H.; Salu, B.R.; Ferreira, R.D.S.; Batista, F.P.; Correia, M.T.D.S.; Paiva, P.M.G.; Turnšek, T.L.; Oliva, M.L.V. A Bifunctional Molecule with Lectin and Protease Inhibitor Activities Isolated from Crataeva tapia Bark Significantly Affects Cocultures of Mesenchymal Stem Cells and Glioblastoma Cells. Molecules 2019, 24, 2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theodoro-Júnior, O.A.; Righetti, R.F.; Almeida-Reis, R.; Martins-Oliveira, B.T.; Oliva, L.V.; Prado, C.M.; Saraiva-Romanholo, B.M.; Leick, E.A.; Pinheiro, N.M.; Lobo, Y.A.; et al. A Plant Proteinase Inhibitor from Enterolobium contortisiliquum Attenuates Pulmonary Mechanics, Inflammation and Remodeling Induced by Elastase in Mice. Int. J. Mol. Sci. 2017, 18, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, A.P.D.; Bortolozzo, A.S.S.; Arantes-Costa, F.M.; Saraiva-Romanholo, B.M.; De Souza, F.C.R.; Brüggemann, T.R.; Santana, F.P.R.; De Brito, M.V.; Bonturi, C.R.; Nunes, N.N.D.S.; et al. A plant proteinase inhibitor from Enterolobium contortisiliquum attenuates airway hyperresponsiveness, inflammation and remodeling in a mouse model of asthma. Histol. Histopathol. 2018, 34, 537–552. [Google Scholar] [PubMed]
- Nakahata, A.M.; Mayer, B.; Ries, C.; De Paula, C.A.A.; Karow, M.; Neth, P.; Sampaio, M.U.; Jochum, M.; Oliva, M.L.V. The effects of a plant proteinase inhibitor from Enterolobium contortisiliquum on human tumor cell lines. Biol. Chem. 2011, 392, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, I.F.; Oliva, M.L.V.; Araujo, M.S.; Sampaio, M.U.; Richardson, M.; Fritz, H.; Sampaio, C.A. Primary structure of a Kunitz-type trypsin inhibitor from Enterolobium contortisiliquum seeds. Phytochemistry 1996, 41, 1017–1022. [Google Scholar] [CrossRef]
- Almeida-Reis, R.; Theodoro-Junior, O.A.; Oliveira, B.T.M.; Oliva, L.V.; Toledo-Arruda, A.C.; Bonturi, C.R.; Brito, M.V.; Lopes, F.D.T.Q.S.; Prado, C.M.; Florencio, A.C.; et al. Plant Proteinase Inhibitor BbCI Modulates Lung Inflammatory Responses and Mechanic and Remodeling Alterations Induced by Elastase in Mice. BioMed Res. Int. 2017, 2017, 8287125. [Google Scholar] [CrossRef] [Green Version]
- Lerman, I.; Hammes, S.R. Neutrophil elastase in the tumor microenvironment. Steroids 2018, 133, 96–101. [Google Scholar] [CrossRef]
- Koumandou, V.L.; Scorilas, A. Evolution of the Plasma and Tissue Kallikreins, and Their Alternative Splicing Isoforms. PLoS ONE 2013, 8, e68074. [Google Scholar] [CrossRef]
- Shi, K.; Wang, S.-L.; Shen, B.; Yu, F.-Q.; Weng, D.-F.; Lin, J.-H. Clinicopathological and prognostic values of fibronectin and integrin αvβ3 expression in primary osteosarcoma. World J. Surg. Oncol. 2019, 17, 1–12. [Google Scholar] [CrossRef]
- Cagliari, C.I.; De Caroli, F.P.; Nakahata, A.M.; Araújo, M.S.; Nakaie, C.R.; Sampaio, M.U.; Sampaio, C.A.M.; Oliva, M.L.V. Action of Bauhinia bauhinioides synthetic peptides on serine proteinases. Biochem. Biophys. Res. Commun. 2003, 311, 241–245. [Google Scholar] [CrossRef]
- Figueroa, C.D.; Molina, L.; Bhoola, K.D.; Ehrenfeld, P. Overview of tissue kallikrein and kallikrein-related peptidases in breast cancer. Biol. Chem. 2018, 399, 937–957. [Google Scholar] [CrossRef] [PubMed]
- Oikonomopoulou, K.; Hansen, K.K.; Saifeddine, M.; Vergnolle, N.; Tea, I.; Diamandis, E.P.; Hollenberg, M.D. Proteinase-mediated cell signalling: Targeting proteinase-activated receptors (PARs) by kallikreins and more. Biol. Chem. 2006, 387, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, J.S.; Talmage, K.E.; Liu, H.; La Jeunesse, C.M.; Witte, D.P.; Degen, J.L. Plasminogen supports tumor growth through a fibrinogen-dependent mechanism linked to vascular patency. Blood 2003, 102, 2819–2827. [Google Scholar] [CrossRef] [PubMed]
- Nyberg, P.; Ylipalosaari, M.; Sorsa, T.; Salo, T. Trypsins and their role in carcinoma growth. Exp. Cell Res. 2006, 312, 1219–1228. [Google Scholar] [CrossRef]
- Zhou, D.; Hansen, D.; Shabalin, I.G.; Gustchina, A.; Vieira, D.F.; De Brito, M.V.; Araújo, A.P.U.; Oliva, M.L.V.; Wlodawer, A. Structure of BbKI, a disulfide-free plasma kallikrein inhibitor. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2015, 71, 1055–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.-L.; Zhao, Y.-F.; Tan, C.-H.; Xiong, Y.-J.; Wang, W.-J.; Wu, F.; Fei, Y.; Wang, L.; Liang, Z.-Q. Cathepsin L upregulation-induced EMT phenotype is associated with the acquisition of cisplatin or paclitaxel resistance in A549 cells. Acta Pharmacol. Sin. 2016, 37, 1606–1622. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Shen, X.; Zhu, Y.; Wang, A.; Xiong, Y.; Wang, L.; Fei, Y.; Wang, Y.; Wang, W.; Lin, F.; et al. Cathepsin L-mediated resistance of paclitaxel and cisplatin is mediated by distinct regulatory mechanisms. J. Exp. Clin. Cancer Res. 2019, 38, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Araújo, A.P.U.; Hansen, D.; Vieira, D.F.; De Oliveira, C.; Santana, L.A.; Beltramini, L.M.; Sampaio, C.A.; Sampaio, M.U.; Oliva, M.L.V. Kunitz-type Bauhinia bauhinioides inhibitors devoid of disulfide bridges: Isolation of the cDNAs, heterologous expression and structural studies. Biol. Chem. 2005, 386, 561–568. [Google Scholar] [CrossRef]
- Morrison, J.F. The slow-binding and slow, tight-binding inhibition of enzyme-catalysed reactions. Trends Biochem. Sci. 1982, 7, 102–105. [Google Scholar] [CrossRef]
- Anastasi, A.; Brown, M.A.; Kembhavi, A.A.; Nicklin, M.J.H.; Sayers, C.A.; Sunter, D.C.; Barrett, A.J. Cystatin, a protein inhibitor of cysteine proteinases. Improved purification from egg white, characterization, and detection in chicken serum. Biochem. J. 1983, 211, 129–138. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | BbCI | rBbCI | BbKI | rBbKI | EcTI |
|---|---|---|---|---|---|
| Trypsin | φ | φ | 20.00 | 28.00 | 0.88 |
| Chymotrypsin | φ | φ | 26.00 | ND | 1.11 |
| Plasmin | φ | φ | 330.00 | ND | 9.36 |
| HNE | 5.30 | 1.70 | φ | φ | 55.0 |
| Factor Xa | φ | ND | φ | ND | φ |
| Thrombin | φ | ND | φ | ND | ND |
| PKa | φ | φ | 2.40 | 2.00 | 6.15 |
| PoPK | φ | φ | 200.00 | 900.00 | φ |
| Cathepsin G | 160.00 | ND | φ | φ | ND |
| Cathepsin L | 0.22 | ND | φ | ND | ND |
| Cruzain | 0.30 | 0.30 | φ | φ | ND |
| Cruzipain | 1.30 | 1.20 | φ | φ | ND |
| Enzyme | Substrate | Buffer |
|---|---|---|
| trypsin (20 μL, 0.41 μM) * NPGB | Bz-Arg-pNan (25 μL, 10 mM) | Tris/HCl 0.1 M, pH 8.0, CaCl2 0.02% (v/v) |
| HNE (20 μL, 0.21 μM) (* α1-anti-trypsin) | MeO-Suc-Ala-Ala-Pro-Val-pNan (25 μL, 1.1 mM) | Tris/HCl 0.1 M, pH 7.0, NaCl 0.5 M |
| PKa (20 μL, 0.84 μM) (* EcTI) | H-D-Pro-Phe-Arg-pNan (25 μL, 5 mM) | Tris/HCl 0.05 M, pH 8.0 |
| PoPK (30 μL, 0.16 nM) (* aprotinin) | H-D-Pro-Phe-Arg-AMC (30 μL, 5 mM) | Tris/HCl 0.1 M, pH 9.0, plus albumin 0.1% (v/v) |
| chymotrypsin (40 μL, 0.88 µM) (* EcTI) | Suc-Phe-pNan (20 μL, 20 mM) | Tris/HCl 0.1 M, pH 8.0, CaCl2 0.02% (v/v) |
| plasmin (25 μL, 0.028 μM) (* BvTI) | H-D-Val-Leu-Lys-pNan (20 μL, 9 mM) | Tris/HCl 0.1 M, pH 7.4, NaCl 0.2 M |
| thrombin (10 μL, 0.267 μM) (* rhodinin) | H-D-Phe-L-Pip-L-Arg-pNan (20 μL, 2 mM) | Tris/HCl 0.05 M, pH 8.0 |
| factor Xa (30 μL, 0.467 μM) (* BuXI) | Boc-Ile-Glu-Gly-Arg-AMC (60 μL, 6 mM) | Tris/HCl 0.05 M, pH 8.0 |
| cathepsin G (30 μL, 0.25 μM) (* α1-anti-trypsin) | N-Suc-Ala-Ala-Pro-Phe-pNan (25 μL, 1 mM) | Tris/HCl 0.05 M, pH 7.0, NaCl 0.5 M |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoo Im, S.; Ramalho Bonturi, C.; Miti Nakahata, A.; Ryuichi Nakaie, C.; Pott, A.; Pott, V.J.; Vilela Oliva, M.L. Differences in the Inhibitory Specificity Distinguish the Efficacy of Plant Protease Inhibitors on Mouse Fibrosarcoma. Plants 2021, 10, 602. https://doi.org/10.3390/plants10030602
Yoo Im S, Ramalho Bonturi C, Miti Nakahata A, Ryuichi Nakaie C, Pott A, Pott VJ, Vilela Oliva ML. Differences in the Inhibitory Specificity Distinguish the Efficacy of Plant Protease Inhibitors on Mouse Fibrosarcoma. Plants. 2021; 10(3):602. https://doi.org/10.3390/plants10030602
Chicago/Turabian StyleYoo Im, Sonia, Camila Ramalho Bonturi, Adriana Miti Nakahata, Clóvis Ryuichi Nakaie, Arnildo Pott, Vali Joana Pott, and Maria Luiza Vilela Oliva. 2021. "Differences in the Inhibitory Specificity Distinguish the Efficacy of Plant Protease Inhibitors on Mouse Fibrosarcoma" Plants 10, no. 3: 602. https://doi.org/10.3390/plants10030602
APA StyleYoo Im, S., Ramalho Bonturi, C., Miti Nakahata, A., Ryuichi Nakaie, C., Pott, A., Pott, V. J., & Vilela Oliva, M. L. (2021). Differences in the Inhibitory Specificity Distinguish the Efficacy of Plant Protease Inhibitors on Mouse Fibrosarcoma. Plants, 10(3), 602. https://doi.org/10.3390/plants10030602