
Distribution of Orchids with Different Rooting Systems in the Czech Republic



Abstract
:1. Introduction
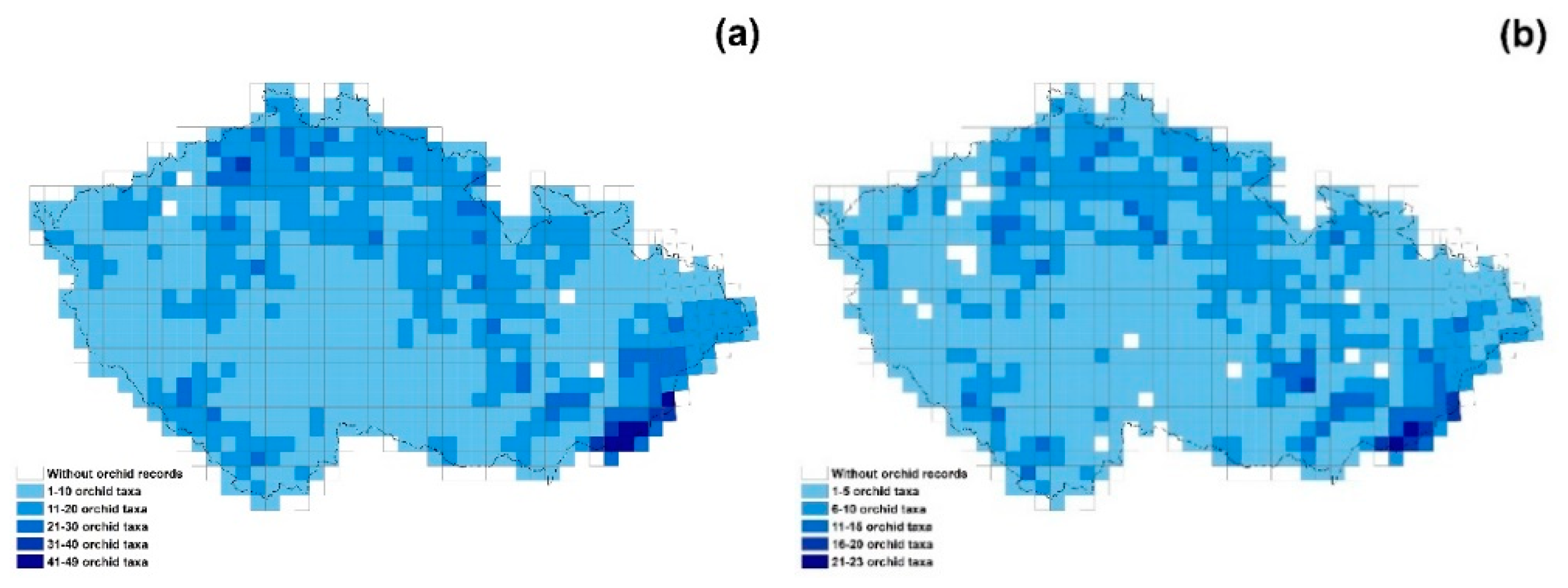
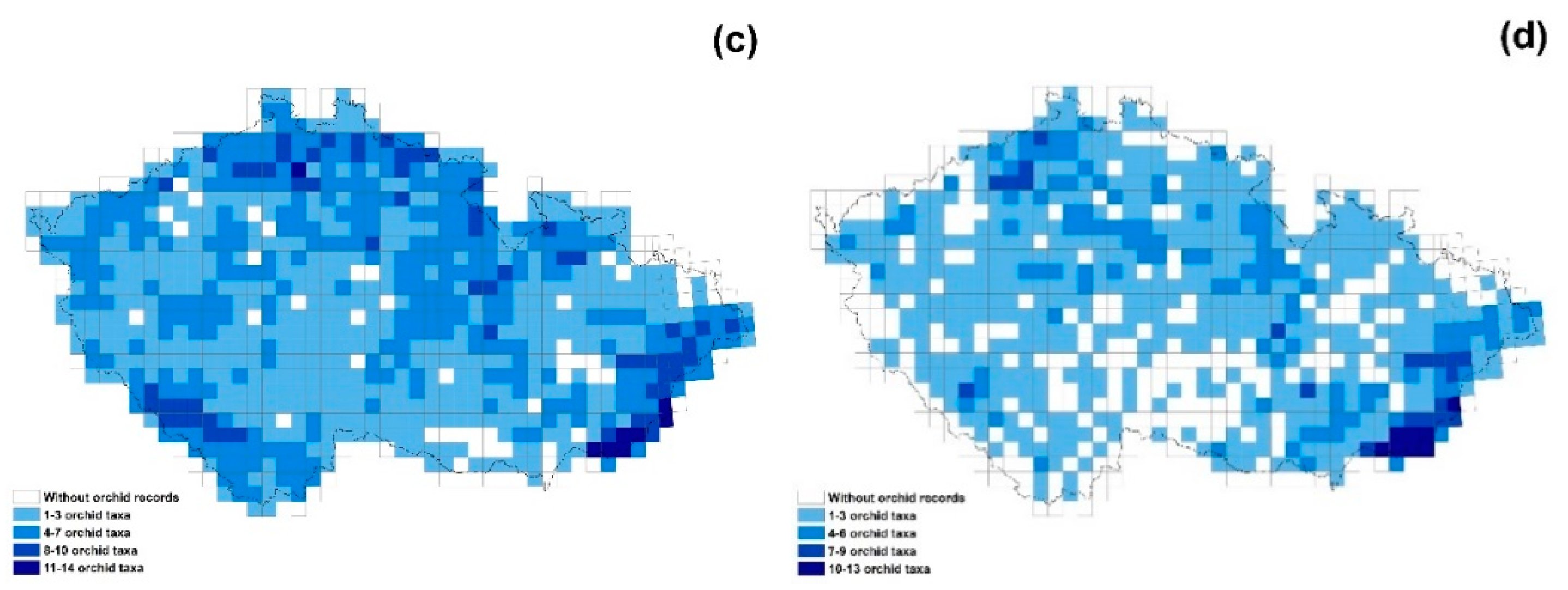
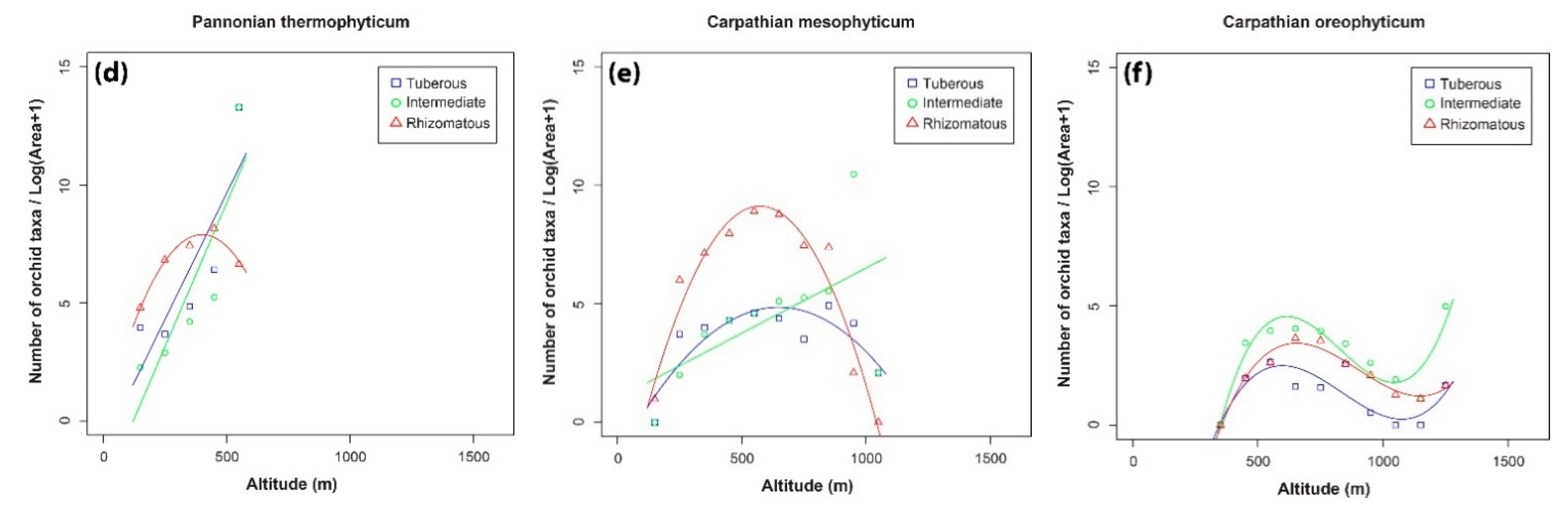
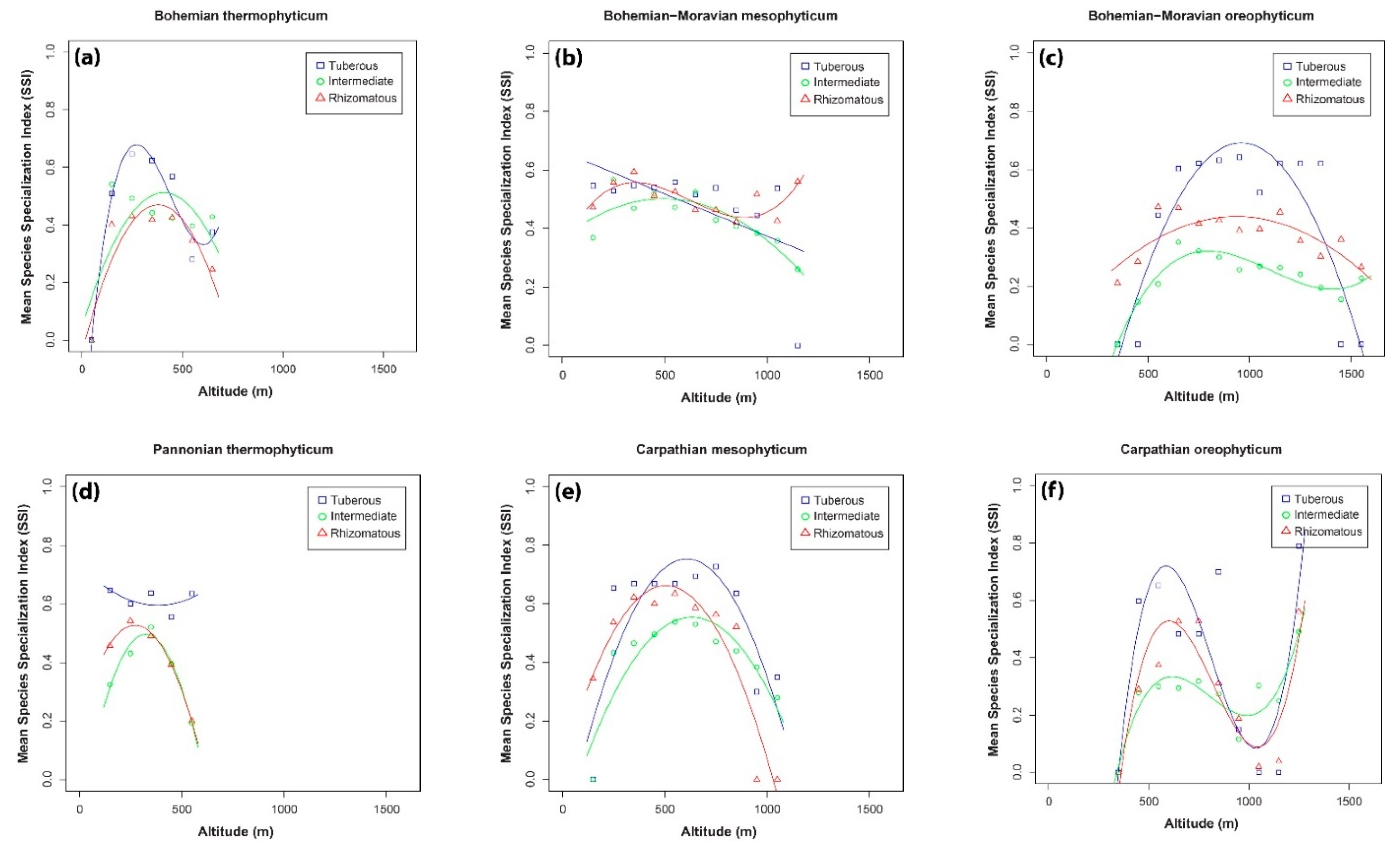
2. Results
3. Discussion
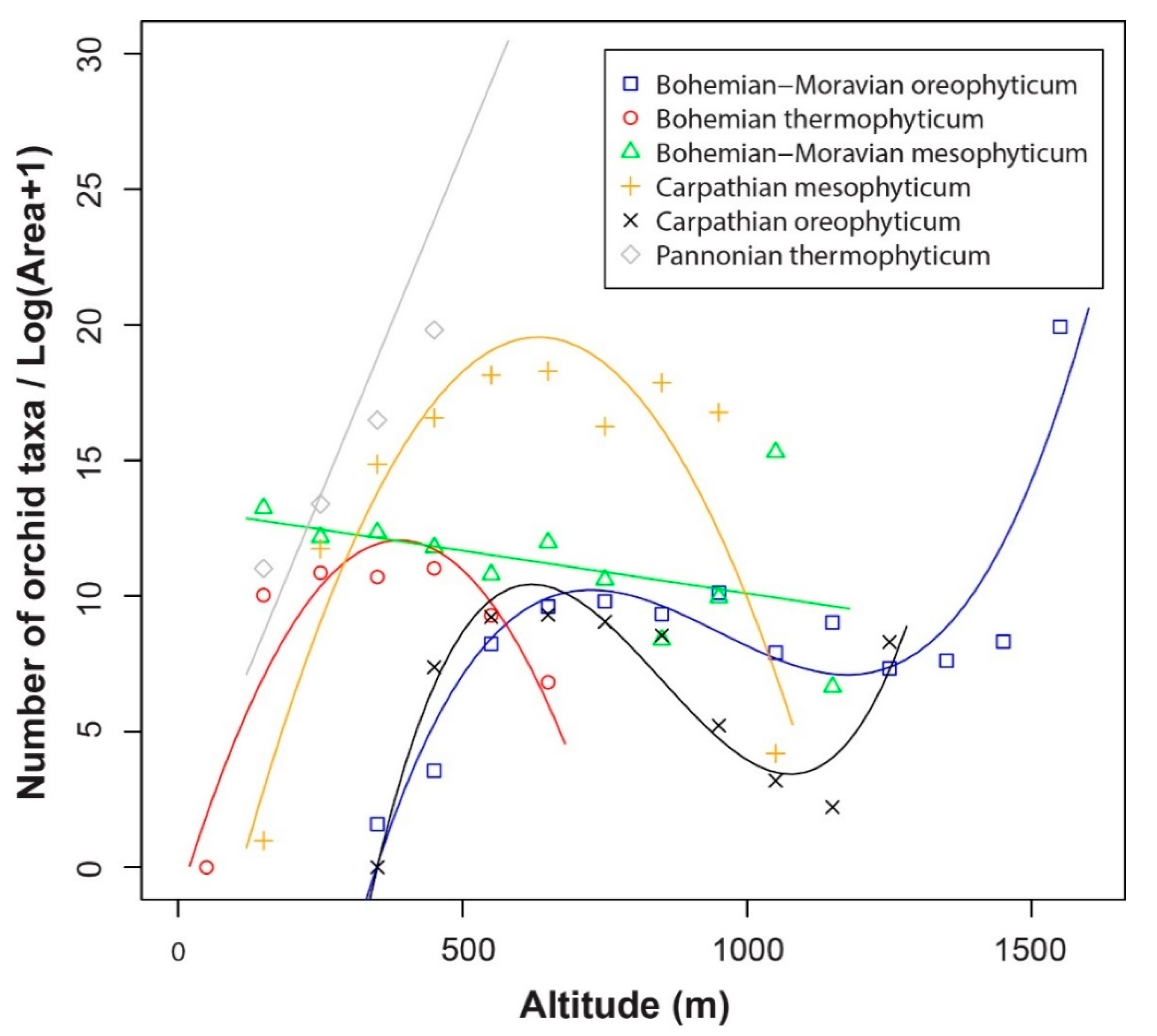
3.1. Orchid Species Density along Altitudinal Gradients
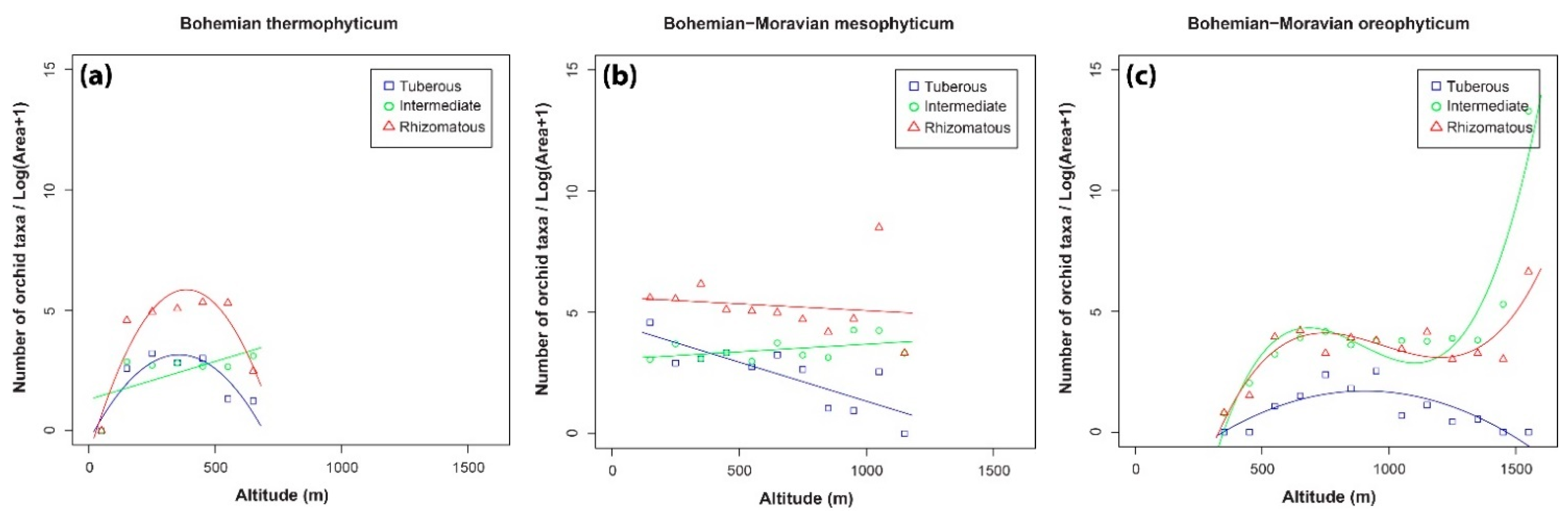
3.2. Patterns in the Distributions of the Three Orchid Groups
3.3. Orchid Species Richness in the Different Phytogeographical Areas
3.4. Relationship between Mean Species Specialization Index and Altitude
4. Materials and Methods
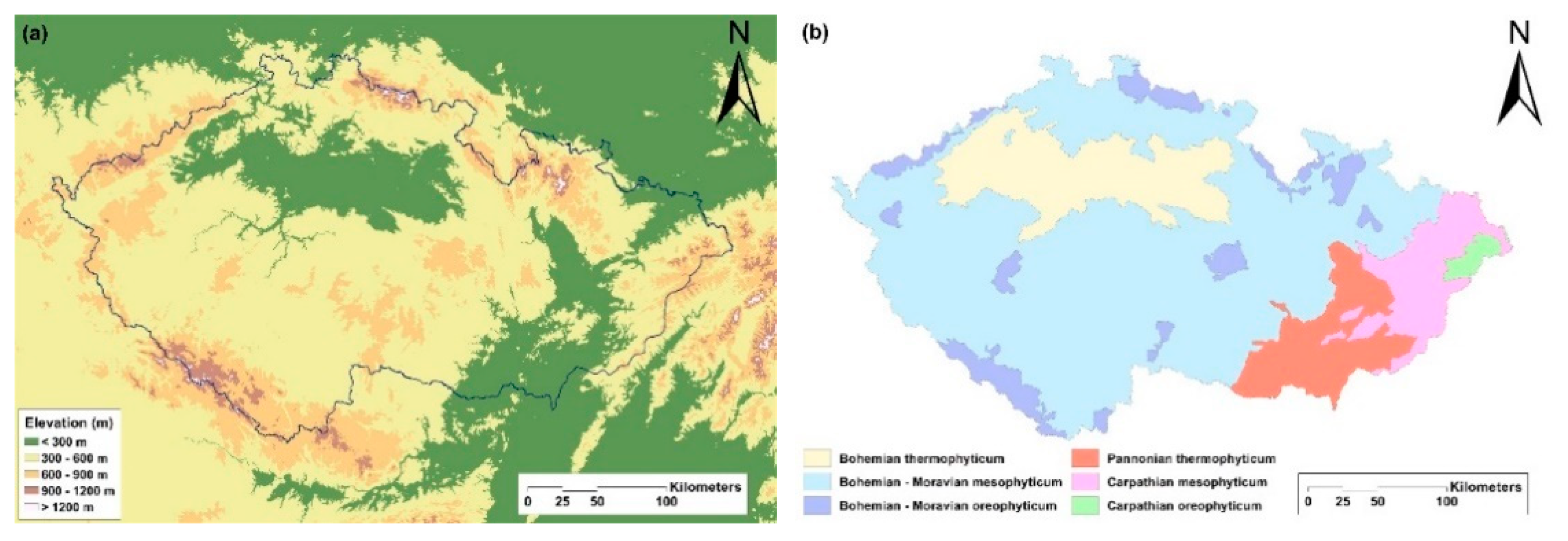
4.1. Areas Studied
4.2. Dataset
4.3. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wotavová, K.; Balounová, Z.; Kindlmann, P. Factors affecting persistence of terrestrial orchids in wet meadows and implications for their conservation in a changing agricultural landscape. Biol. Conserv. 2004, 118, 271–279. [Google Scholar] [CrossRef]
- Pfeifer, M.; Wiegand, K.; Heinrich, W.; Jetschke, G. Long-term demographic fluctuations in an orchid species driven by weather: Implications for conservation planning. J. Appl Ecol. 2006, 43, 313–324. [Google Scholar] [CrossRef]
- Swarts, N.D.; Dixon, K.W. Conservation Methods for Terrestrial Orchids; J. Ross Publishing: Plantation, FL, USA, 2017. [Google Scholar]
- Štípková, Z.; Romportl, D.; Černocká, V.; Kindlmann, P. Factors associated with the distributions of orchids in the Jeseníky Mountains, Czech Republic. Eur. J. Environ. Sci. 2017, 7, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Štípková, Z.; Kosánová, K.; Romportl, D.; Kindlmann, P. Determinants of Orchid Occurrence: A Czech Example. In Selected Studies in Biodiversity; Şen, B., Grillo, O., Eds.; InTech Open: London, UK, 2018; pp. 1–24. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.H. Macroecology; University of Chicago Press: Chicago, IL, USA, 1995. [Google Scholar]
- Gaston, K.J.; Blackburn, T.M. Pattern and Process in Macroecology; Blackwell Science Ltd.: Malden, MA, USA, 2000. [Google Scholar]
- Tsiftsis, S.; Štípková, Z.; Kindlmann, P. Role of way of life, latitude, elevation and climate in the richness and distribution of orchid species. Biodivers Conserv. 2019, 28, 75–96. [Google Scholar] [CrossRef]
- Swarts, N.D.; Dixon, W.D. Terrestrial orchid conservation in the age of extinction. Ann. Bot-Lond. 2009, 104, 543–556. [Google Scholar] [CrossRef] [Green Version]
- Tsiftsis, S.; Tsiripidis, I.; Karagiannakidou, V.; Alifragis, D. Niche analysis and conservation of orchids of east Macedonia (NE Greece). Acta Oecol. 2008, 33, 27–35. [Google Scholar] [CrossRef]
- Štípková, Z.; Tsiftsis, S.; Kindlmann, P. How did the agricultural policy during the communist period affect the decline in orchid biodiversity in Central and Eastern Europe? Glob. Ecol. Conserv. 2021, 26, e01498. [Google Scholar] [CrossRef]
- Štípková, Z.; Kindlmann, P. Orchid extinction over the last 150 years in the Czech Republic. Diversity 2021, 13, 78. [Google Scholar] [CrossRef]
- Dykyjová, D. Ekologie Středoevropských Orchidejí; KOPP: České Budějovice, Česká republika, 2003. [Google Scholar]
- Jersáková, J.; Kindlmann, P. Zásady Péče o Orchidejová Stanoviště; KOPP: České Budějovice, Česká republika, 2004. [Google Scholar]
- Průša, D. Orchideje České Republiky; Computer press: Brno, Česká republika, 2005. [Google Scholar]
- Slavík, S.; Hejný, B. Květena České Republiky 8; Academia: Praha, Česká republika, 2010. [Google Scholar]
- Danihelka, J.; Chrtek, J., Jr.; Kaplan, Z. Checklist of vascular plants of the Czech Republic. Preslia 2012, 84, 647–811. [Google Scholar]
- Lepší, P.; Lepší, M.; Boublík, K.; Stech, M.; Hans, V. Červená Kniha Květeny Jižní Části Čech; Jihočeské muzeum v Českých Budějovicích: České Budějovice, Česká republika, 2013. [Google Scholar]
- Chytrý, M.; Danihelka, J.; Kaplan, Z.; Pyšek, P. Flora and Vegetation of the Czech. Republic; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar]
- Štípková, Z.; Tsiftsis, S.; Kindlmann, P. Pollination mechanisms are driving orchid distribution in space. Sci. Rep. 2020, 10, 850. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, H.N. Terrestrial Orchids from Seed to Mycotrophic Plant; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Dressler, R.L. The Orchids: Natural History and Classification; Harvard University Press: Cambridge, MA, USA, 1981. [Google Scholar]
- Averyanov, L. A review of the genus Dactylorhiza. In Orchid Biology—Reviews and Perspectives; Arditti, J., Ed.; V. Timber Press Inc.: Portland, OR, USA, 1990; pp. 159–206. [Google Scholar]
- Tatarenko, I. Growth habits of temperate terrestrial orchids. In Orchid Biology—Reviews and Perspectives, IX; Cameron, K.M., Arditti, J., Kull, T., Eds.; The New York Botanical Garden Press: Bronx, NY, USA, 2007; pp. 91–161. [Google Scholar]
- Kaplan, Z. Flora and phytogeography of the Czech Republic. Preslia 2012, 84, 505–573. [Google Scholar]
- Stevens, G.C. The latitudinal gradient in geographical range: How so many species coexist in the tropics. Am. Nat. 1989, 133, 240–256. [Google Scholar] [CrossRef]
- Stevens, G.C. The elevational gradient in altitudinal range: An extension of Rapoport’s latitudinal rule to altitude. Am. Nat. 1992, 140, 893–911. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, K.R.; Vetaas, O.R. Variation in plant species richness of different life forms along a subtropical elevation gradient in the Himalayas, east Nepal. Glob. Ecol. Biogeogr. 2003, 12, 327–340. [Google Scholar] [CrossRef]
- Grytnes, J.A. Species-richness patterns of vascular plants along seven altitudinal transects in Norway. Ecography 2003, 26, 291–300. [Google Scholar] [CrossRef]
- Hrivnák, R.; Gömöry, D.; Slezák, M.; Ujházy, K.; Hédl, R.; Jarčuška, B.; Ujházyová, M. Species richness pattern along altitudinal gradient in central European beech forests. Folia. Geobot. 2014, 49, 425–441. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.D.; Ochola, A.C.; Njogu, A.W.; Boru, B.H.; Mwachala, G.; Hu, G.W.; Xin, H.P.; Wang, Q.F. The species richness pattern of vascular plants along a tropical elevational gradient and the test of elevational Rapoport’s rule depend on different life-forms and phytogeographic affinities. Ecol. Evol. 2019, 9, 4495–4503. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Micheneau, C.; Roberts, D.L.; Pailler, T. Elevational gradients of species diversity, breeding system and floral traits of orchid species on Réunion Island. J. Biogeogr. 2005, 32, 1751–1761. [Google Scholar] [CrossRef]
- Del Prete, C.; Mazzola, P. Endemism and speciation in the orchids of Mediterranean Islands. Ecol. Mediterr. 1995, 21, 119–134. [Google Scholar] [CrossRef]
- Prach, K.; Jongepierová, I.; Řehounková, K. Large-scale restoration of dry grasslands on ex-arable land using a regional seed mixture: Establishment of target species. Restor. Ecol. 2004, 21, 33–39. [Google Scholar] [CrossRef]
- Johanidesová, E.; Fajmon, K.; Jongepierová, I.; Prach, K. Spontaneous colonization of restored dry grasslands by target species: Restoration proceeds beyond sowing regional seed mixtures. Grass Forage Sci. 2003, 70, 631–638. [Google Scholar] [CrossRef]
- Kull, T.; Hutchings, M.J. A comparative analysis of decline in the distribution ranges of orchid species in Estonia and the United Kingdom. Biol. Conserv. 2006, 129, 31–39. [Google Scholar] [CrossRef]
- Swenson, J.J.; Young, B.E.; Beck, S.; Comer, P.; Cordova, J.H.; Dyson, J.; Embert, D.; Encarnacion, F.; Ferreira, W.; Franke, I.; et al. Plant and animal endemism in the eastern Andean slope: Challenges to conservation. BMC Ecol. 2012, 12, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsiftsis, S.; Tsiripidis, I.; Karagiannakidou, V. Identifying areas of high importance for orchid conservation in east Macedonia (NE Greece). Biodivers Conserv. 2009, 18, 1765–1780. [Google Scholar] [CrossRef]
- Djordjević, V.; Tsiftsis, S.; Lakušić, D.; Jovanović, S.; Stevanović, V. Factors affecting the distribution and abundance of orchids in grasslands and herbaceous wetlands. Syst. Biodivers 2016, 14, 355–370. [Google Scholar] [CrossRef]
- Djordjević, V.; Tsiftsis, S. The role of ecological factors in distribution and abundance of terrestrial orchids. In Orchids Phytochemistry, Biology and Horticulture; Mérillon, J.M., Kodja, H., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 1–71. [Google Scholar]
- Pecoraro, K.; Caruso, T.; Cai, L.; Gupta, V.K.; Liu, Z.-J. Fungal networks and orchid distribution: New insight from above- and below-ground analyses of fungal communities. IMA Fungus 2018, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Davis, B.J.; Phillips, R.D.; Wright, M.; Linde, C.C.; Dixon, K.W. Continent-wide distribution in mycorrhizal fungi: Implications for the biogeography of specialized species. Ann. Bot-Lond. 2015, 116, 413–421. [Google Scholar] [CrossRef]
- Carvalho, L.M.; Correia, P.M.; Ryel, R.J.; Martins-Loucao, M.A. Spatial variability of arbuscular mycorrhizal fungal spores in two natural plant communities. Plant. Soil 2003, 251, 227–236. [Google Scholar] [CrossRef]
- McCormick, M.K.; Jacquemyn, H. What constraints the distribution of orchid populations? New Phytol. 2014, 202, 392–400. [Google Scholar] [CrossRef]
- McCormick, M.K.; Whigham, D.F.; Canchani-Viruet, A. Mycorrhizal fungi affect orchid distribution and population dynamics. New Phytol. 2018, 219, 1207–1215. [Google Scholar] [CrossRef] [Green Version]
- Geml, J. Altitudinal gradients in mycorrhizal symbioses. In Biogeography of Mycorrhizal Symbiosis; Tedersoo, L., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 107–123. [Google Scholar]
- Arroyo, M.T.K.; Primack, R.; Armesto, J. Community studies in pollination ecology in the high temperate Andes of central Chile. I. Pollination mechanisms and altitudinal variation. Am. J. Bot. 1982, 69, 82. [Google Scholar] [CrossRef]
- Gilián, L.D.; Enrédi, A.; Zsinka, B.; Neményi, A.; Nagy, J.G. Morphological and reproductive trait-variability of a food deceptive orchid, Cephalanthera rubra along different altitudes. Appl. Ecol. Environ. Res. 2019, 17, 5619–5639. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Brys, R.; Hermy, M.; Willems, J.H. Does nectar reward affect rarity and extinction probabilities of orchid species? An assessment using historical records from Belgium and the Netherlands. Biol. Conserv. 2005, 121, 257–263. [Google Scholar]
- Devoto, M.; Medan, D.; Montaldo, N.H. Patterns of interaction between plants and pollinators along an environmental gradient. Oikos 2005, 109, 461–472. [Google Scholar] [CrossRef]
- Rasmann, S.; Alvarez, N.; Pellissier, L. The altitudinal niche-breadth hypothesis in insect-plant interactions. Annu. Plant. Rev. 2014, 47, 339–360. [Google Scholar]
- Chytrý, M.; Kučera, T.; Kočí, M. Katalog Biotopů České Republiky; AOPK ČR: Praha, Česká republika, 2001. [Google Scholar]
- Grulich, V. The Red List of vascular plants of the Czech Republic. Příroda 2017, 35, 75–132. [Google Scholar]
- Tremblay, R.L. Trends in the pollination ecology of the Orchidaceae: Evolution and systematics. Can. J. Bot. 1992, 70, 642–650. [Google Scholar] [CrossRef]
- Tylianakis, J.M. The global plight of pollinators. Science 2013, 339, 1532–1533. [Google Scholar] [CrossRef]
- Darwin, C. The Various Contrivances by Which Orchids Are Fertilized by Orchids; John Murray: London, UK, 1862. [Google Scholar]
- Neiland, M.R.M.; Wilcock, C.C. Fruit set, nectar reward, and rarity in the Orchidaceae. Am. J. Bot 1998, 85, 1657–1671. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.D.; Brown, A.P.; Dixon, K.W.; Hopper, S.D. Orchid biogeography and factors associated with rarity in a biodiversity hotspot, the Southwest Australian Floristic Region. J. Biogeogr. 2010, 28, 487–501. [Google Scholar] [CrossRef]
- Crain, B.J.; Tremblay, R.L. Do richness and rarity hotspots really matter for orchid conservation inlight of anticipated habitat loss? Divers. Distrib. 2014, 20, 652–662. [Google Scholar] [CrossRef]
- WCSP. World Checklist of Selected Plant. Families. Facilitated by the Royal Botanic Gardens, Kew. 2017. Available online: http://apps.kew.org/wcsp/ (accessed on 5 February 2017).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Grytnes, J.A.; Vetaas, O.R. Species richness and altitude: A comparison between null models and interpolated plant species richness along the Himalayan altitudinal gradient, Nepal. Am. Nat. 2002, 159, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Dolédec, D.; Chessel, D.; Gimaret-Carpentier, C. Niche separation in community analysis: A new method. Ecology 2000, 81, 2914–2927. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; 2013; Available online: http://www.R-project.org/ (accessed on 3 February 2016).
- ESRI. ArcGIS—ArcMap: ArcInfo (version 10.1); Environmental Science Research Institute: Redlands, CA, USA, 2012. [Google Scholar]
- Dray, S.; Dufour, A.; Chessel, D. The ade4 Package—II: Two-Table and K-Table Methods. R News 2007, 7, 47–52. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biogeographical Region | R2 | p Value |
|---|---|---|
| Bohemian thermophyticum | 0.75 (b) | p < 0.05 |
| Bohemian-Moravian mesophyticum | 0.11 (a) | ns |
| Bohemian-Moravian oreophyticum | 0.80 (c) | p < 0.001 |
| Pannonian thermophyticum | 0.79 (a) | p < 0.05 |
| Carpathian mesophyticum | 0.81 (b) | p < 0.001 |
| Carpathian oreophyticum | 0.86 (c) | p < 0.001 |
| Root System | Biogeographical Region | R2 | p Value |
|---|---|---|---|
| Tuberous | Bohemian thermophyticum | 0.69(b) | p < 0.05 |
| Pannonian thermophyticum | 0.63(a) | p < 0.05 | |
| Bohemian-Moravian mesophyticum | 0.64(a) | p < 0.01 | |
| Carpathian mesophyticum | 0.62(b) | p < 0.05 | |
| Bohemian-Moravian oreophyticum | 0.59(b) | p < 0.01 | |
| Carpathian oreophyticum | 0.55(c) | p < 0.05 | |
| Intermediate | Bohemian thermophyticum | 0.31(a) | p < 0.05 |
| Pannonian thermophyticum | 0.66(a) | p < 0.05 | |
| Bohemian-Moravian mesophyticum | 0.12(a) | ns | |
| Carpathian mesophyticum | 0.27(a) | ns | |
| Bohemian-Moravian oreophyticum | 0.83(c) | p < 0.001 | |
| Carpathian oreophyticum | 0.78(c) | p < 0.01 | |
| Rhizomatous | Bohemian thermophyticum | 0.80(b) | p < 0.05 |
| Pannonian thermophyticum | 0.95(b) | p < 0.05 | |
| Bohemian-Moravian mesophyticum | 0.09(a) | ns | |
| Carpathian mesophyticum | 0.92(b) | p < 0.001 | |
| Bohemian-Moravian oreophyticum | 0.69(c) | p < 0.01 | |
| Carpathian oreophyticum | 0.93(b) | p < 0.001 |
| Biogeographical Region | Root System | R2 | p Value |
|---|---|---|---|
| Bohemian thermophyticum | Tuberous | 0.91(c) | p < 0.01 |
| Intermediate | 0.27(b) | ns | |
| Rhizomatous | 0.73(b) | p < 0.05 | |
| Bohemian-Moravian mesophyticum | Tuberous | 0.28(a) | p < 0.05 |
| Intermediate | 0.69(b) | p < 0.01 | |
| Rhizomatous | 0.45(c) | p < 0.05 | |
| Bohemian-Moravian oreophyticum | Tuberous | 0.77(b) | p < 0.001 |
| Intermediate | 0.87(c) | p < 0.001 | |
| Rhizomatous | 0.51(b) | p < 0.01 | |
| Pannonian thermophyticum | Tuberous | 0.38(b) | ns |
| Intermediate | 0.93(b) | p < 0.05 | |
| Rhizomatous | 0.98(b) | p < 0.01 | |
| Carpathian mesophyticum | Tuberous | 0.63(b) | p < 0.01 |
| Intermediate | 0.73(b) | p < 0.01 | |
| Rhizomatous | 0.84(b) | p < 0.001 | |
| Carpathian oreophyticum | Tuberous | 0.54(c) | p < 0.05 |
| Intermediate | 0.75(c) | p < 0.01 | |
| Rhizomatous | 0.74(c) | p < 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Štípková, Z.; Tsiftsis, S.; Kindlmann, P. Distribution of Orchids with Different Rooting Systems in the Czech Republic. Plants 2021, 10, 632. https://doi.org/10.3390/plants10040632
Štípková Z, Tsiftsis S, Kindlmann P. Distribution of Orchids with Different Rooting Systems in the Czech Republic. Plants. 2021; 10(4):632. https://doi.org/10.3390/plants10040632
Chicago/Turabian StyleŠtípková, Zuzana, Spyros Tsiftsis, and Pavel Kindlmann. 2021. "Distribution of Orchids with Different Rooting Systems in the Czech Republic" Plants 10, no. 4: 632. https://doi.org/10.3390/plants10040632
APA StyleŠtípková, Z., Tsiftsis, S., & Kindlmann, P. (2021). Distribution of Orchids with Different Rooting Systems in the Czech Republic. Plants, 10(4), 632. https://doi.org/10.3390/plants10040632