Transcriptomic Analyses Shed Light on Critical Genes Associated with Bibenzyl Biosynthesis in Dendrobium officinale

 ,
, 
Abstract
:1. Introduction
2. Results
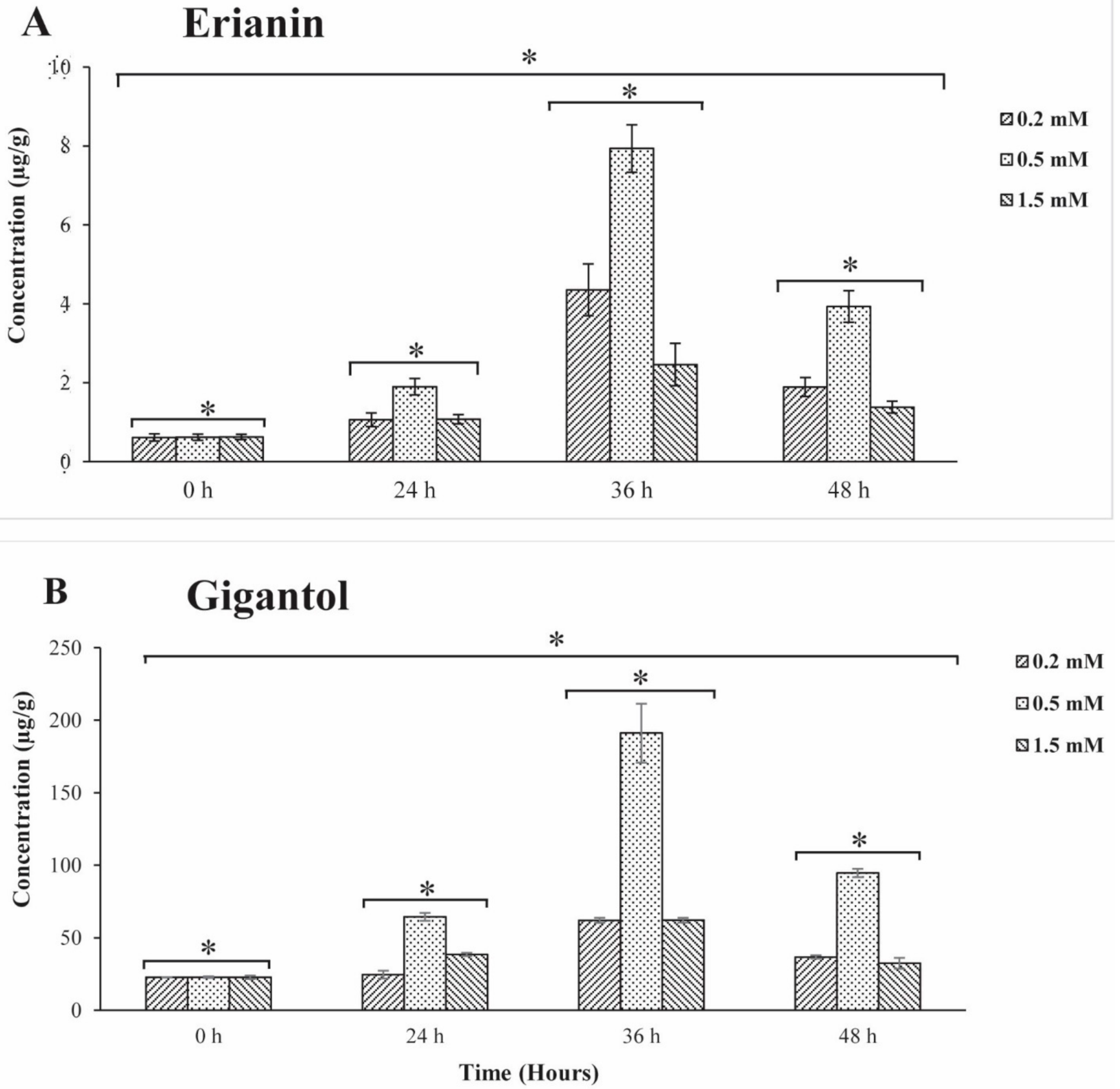
2.1. Investigation of Bibenzyl Accumulation among Tissues
2.2. Transcriptome Sequencing Datasets
2.3. Identification of Differentially Expressed Genes
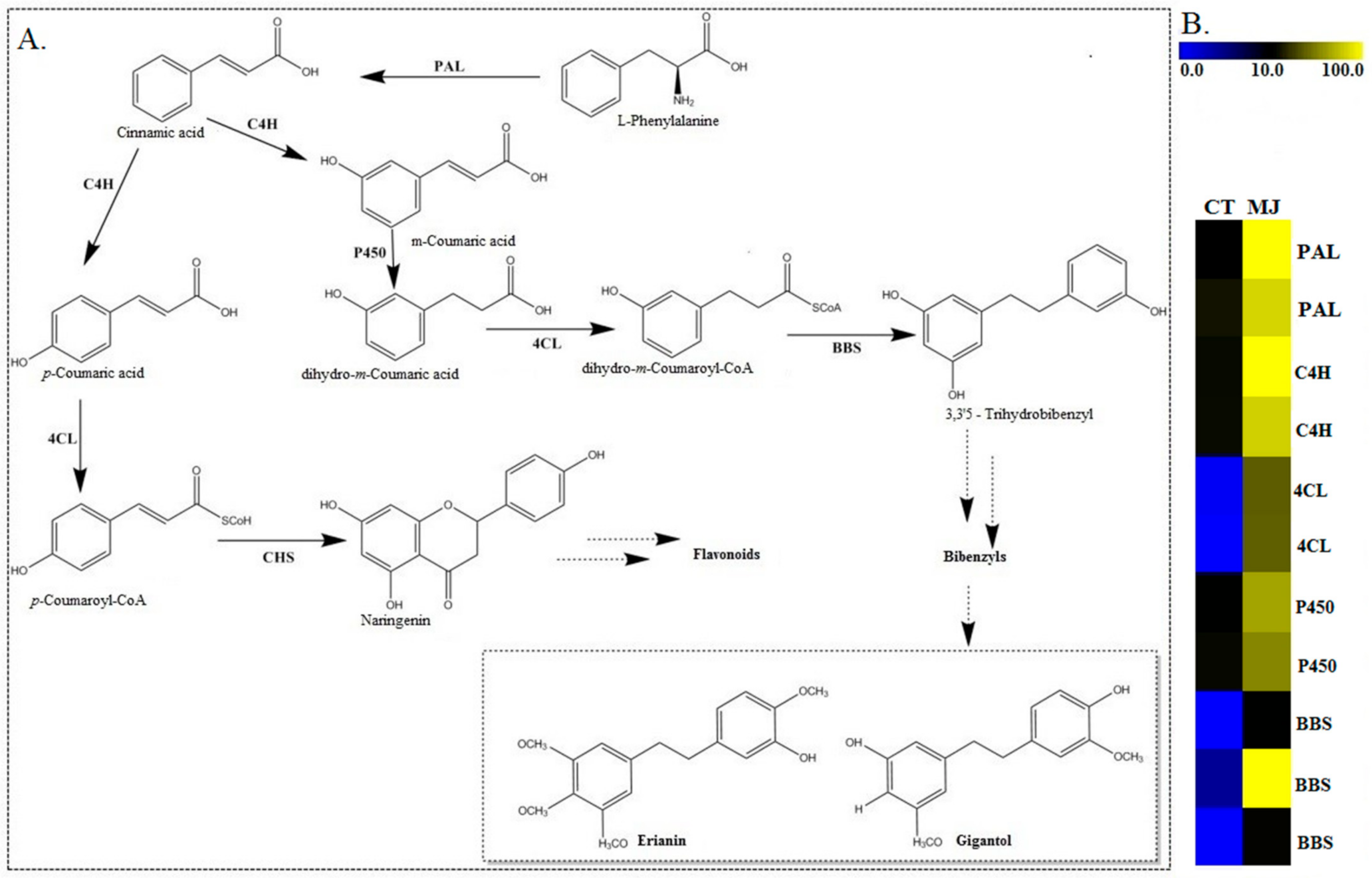
2.4. Identification of Candidate Genes Involved in Bibenzyl Biosynthesis
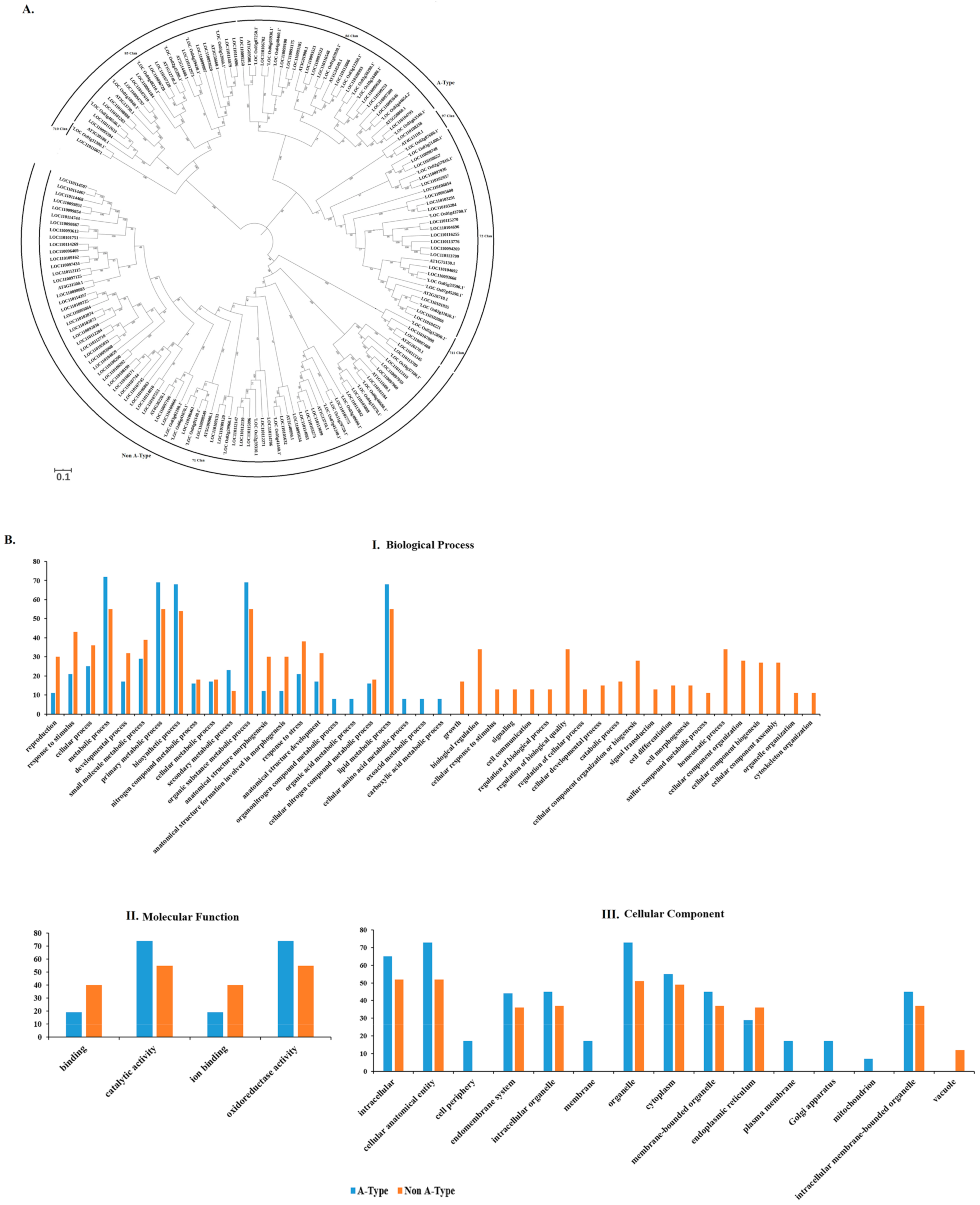
2.5. Identification, Phylogenetic Analysis, and Classification of CYP450 Gene Family in Dendrobium
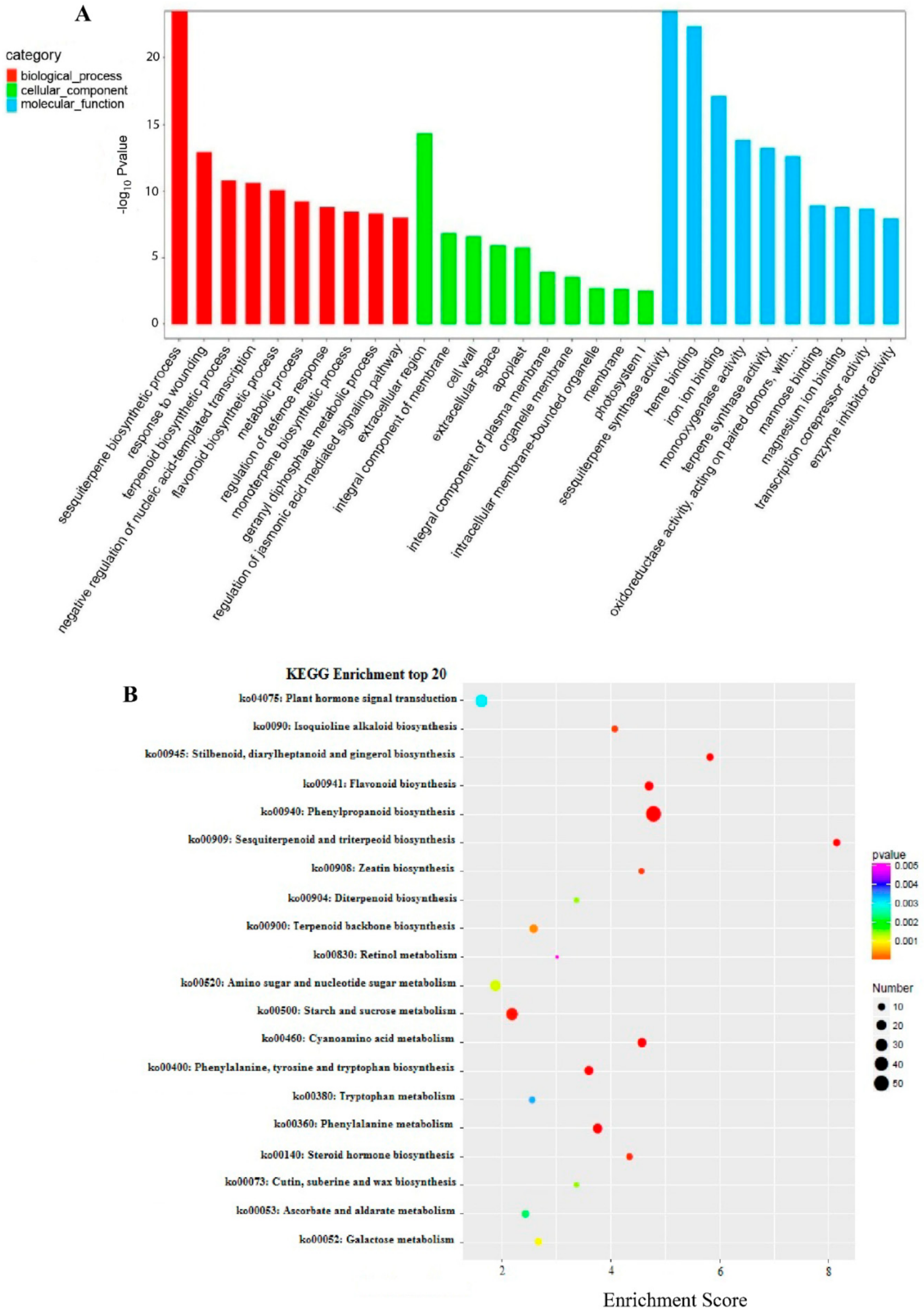
2.6. Functional Annotation of Dendrobium officinale CYP450 Genes
2.7. KEGG Pathway Analysis of D. officinale CYP450 Genes
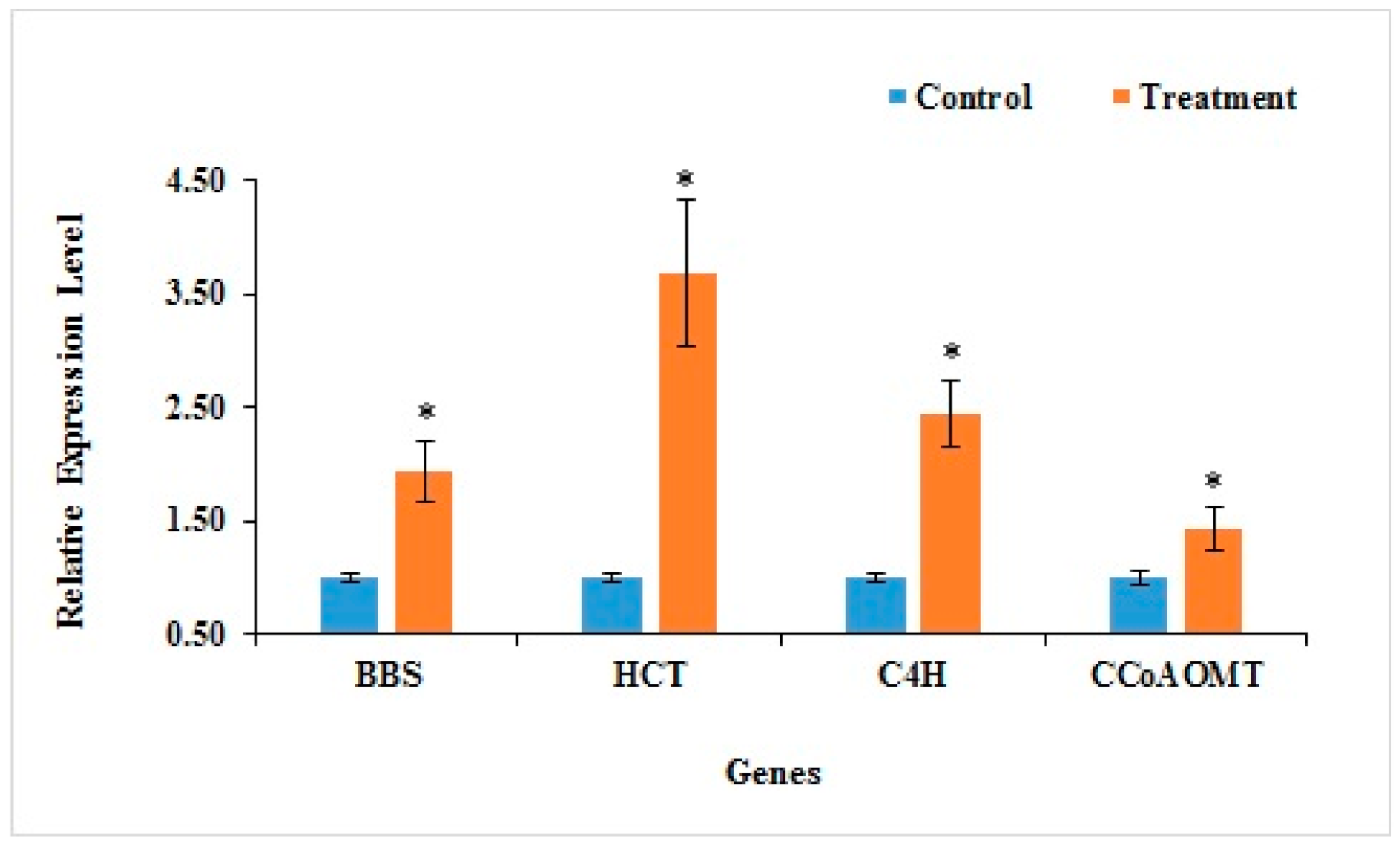
2.8. qRT-PCR Verification
3. Discussion
4. Materials and Methods
4.1. Plant Material and Sample Collection
4.2. Extraction of Plant Materials and Analysis of Bibenzyl Content
4.3. MeJA Treatment Conditions
4.4. RNA Extraction, cDNA Library Preparation, and Transcriptome Sequencing
4.5. Illumina Sequencing Data Analysis
4.6. Analysis of Differentially Expressed Genes (DEGs)
4.7. Functional Annotation
4.8. Alignment and Phylogenetic Study of the Gene Family Dendrobium CYP450
4.9. Classification and Characterization of CYP450 Genes in Dendrobium
Validation of Genes Related to Bibenzyl Biosynthesis by qRT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dixon, R.A. Natural products and plant disease resistance. Nature 2001, 411, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.O.; Wightman, E.L. Herbal extracts and phytochemicals: Plant secondary metabolites and the enhancement of human brain function. Adv. Nutr. 2011, 2, 32–50. [Google Scholar] [CrossRef] [PubMed]
- Kliebenstein, D.J.; Osbourn, A. Making new molecules—Evolution of pathways for novel metabolites in plants. Curr. Opin. Plant Biol. 2012, 15, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Goossens, A.H.; Laakso, S.T.; Seppanen-Laakso, I.; Biondi, T.; De Sutter, S.; Lammertyn, V.; Nuutila, F.; Soderlund, A.M.; Zabeau, H.; Inze, M.; et al. A functional genomics approach toward the understanding of secondary metabolism in plant cells. Proc. Natl. Acad. Sci. USA 2003, 100, 8595–8600. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.S.F.; Ansari, S.; Rahman, S.; Ahmad, M.A.; Saeed, I.Z. Current approaches toward production of secondary plant metabolites. J. Pharm. Bioallied Sci. 2012, 4, 10–20. [Google Scholar] [CrossRef]
- Yang, D.L.; Yang, J.; Mei, C.S.; Tong, X.H.; Zeng, L.J.; Li, Q.; Xiao, L.T.; Sun, T.P.; Li, J.; Deng, X.W.; et al. Plant hormone jasmonate prioritizes defense over growth by interfering with gibberellin signaling cascade. Proc. Natl. Acad. Sci. USA 2012, 109, E1192–E1200. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.Y.; Raven, P.H.; Hong, D.Y. Flora of China. Harv. Pap. Bot. 2010, 25, 347. [Google Scholar]
- Yang, S. Comprehensive Utilization of Citrus By-Products. Methods for Determining the Functional Components of Citrus Peel; Elsevier: Amsterdam, The Netherlands, 2016; pp. 15–30. [Google Scholar]
- Zhang, J.; He, C.; Wu, K.; Teixeira da Silva, J.A.; Zeng, S.; Zhang, X.; Yu, Z.; Xia, H.; Duan, J. Transcriptome Analysis of Dendrobium officinale and its Application to the Identification of Genes Associated with Polysaccharide Synthesis. Front. Plant Sci. 2016, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Zhao, T.; Sheng, Y.; Zheng, T.; Fu, L.; Zhang, Y. Dendrobium officinale Kimura et Migo: A Review on Its Ethnopharmacology, Phytochemistry, Pharmacology, and Industrialization. Evid. Based Complementary Altern. Med. eCAM 2017, 2017, 7436259. [Google Scholar] [CrossRef] [Green Version]
- Kreis, L.M.; Carreira, E.M. Total synthesis of (-)-dendrobine. Angew. Chem. Int. Ed. Engl. 2012, 51, 3436–3439. [Google Scholar] [CrossRef]
- Li, Q.; Ding, G.; Li, B.; Guo, S.X. Transcriptome Analysis of Genes Involved in Dendrobine Biosynthesis in Dendrobium nobile Lindl. Infected with Mycorrhizal Fungus MF23 (Mycena sp.). Sci. Rep. 2017, 7, 316. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhao, W.M.; Qian, Z.M.; Guan, J.; Li, S.P. Fast determination of five components of coumarin, alkaloids and bibenzyls in Dendrobium spp. using pressurized liquid extraction and ultra-performance liquid chromatography. J. Sep. Sci. 2010, 33, 1580–1586. [Google Scholar] [CrossRef]
- Gong, Y.Q. Mechanisms of Erianin Anti-Tumor Angiogenesis; China Pharmaceutical University: Nanjing, China, 2003. [Google Scholar]
- Barbosa, E.G.; Bega, L.A.; Beatriz, A.; Sarkar, T.; Hamel, E.; do Amaral, M.S.; de Lima, D.P. A diaryl sulfide, sulfoxide, and sulfone bearing structural similarities to combretastatin A-4. Eur. J. Med. Chem. 2009, 44, 2685–2688. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, C.; Wang, F. Chemical constituents of Dendrobium candidum. China J. Chin. Mater. Med. 2010, 35, 1715–1719. [Google Scholar]
- Su, P. Research on the Molecular Mechanism of Erianin AntiHepatoma Effect; University of Chinese Academy of Sciences: Beijing, China, 2011. [Google Scholar]
- Cai, H.L.; Huang, X.J.; Nie, S.P.; Xie, M.Y.; Phillips, G.O.; Cui, S.W. Study on Dendrobium officinale O-acetylglucomannan (Dendronan): Part III-Immunomodulatory activity in vitro. Bioact. Carbohydr. Diet. Fibre 2015, 5, 99–105. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, J.K.; Wang, J.; Wang, N.L.; Kurihara, H.; Kitanaka, S.; Yao, X.S. Bioactive bibenzyl derivatives and fluorenones from Dendrobium nobile. J. Nat. Prod. 2007, 70, 24–28. [Google Scholar] [CrossRef]
- Hossain, M.M. Therapeutic orchids: Traditional uses and recent advances—An overview. Fitoterapia 2011, 82, 102–140. [Google Scholar] [CrossRef]
- Majumder, P.L.; Sen, S.; Majumder, S. Phenanthrene derivatives from the orchid Coelogyne cristata. Phytochemistry 2001, 58, 581–586. [Google Scholar] [CrossRef]
- Yahyaa, M.; Ali, S.; Davidovich-Rikanati, R.; Ibdah, M.; Shachtier, A.; Eyal, Y.; Lewinsohn, E.; Ibdah, M. Characterization of three chalcone synthase-like genes from apple (Malus x domestica Borkh.). Phytochemistry 2017, 140, 125–133. [Google Scholar] [CrossRef]
- Ibdah, M.; Martens, S.; Gang, D.R. Biosynthetic Pathway and Metabolic Engineering of Plant Dihydrochalcones. J. Agric. Food Chem. 2018, 66, 2273–2280. [Google Scholar] [CrossRef]
- Jeong, Y.J.; An, C.H.; Woo, S.G.; Park, J.H.; Lee, K.W.; Lee, S.H.; Rim, Y.; Jeong, H.J.; Ryu, Y.B.; Kim, C.Y. Enhanced production of resveratrol derivatives in tobacco plants by improving the metabolic flux of intermediates in the phenylpropanoid pathway. Plant. Mol. Biol. 2016, 92, 117–129. [Google Scholar] [CrossRef]
- Peled-Zehavi, H.; Oliva, M.; Xie, Q.; Tzin, V.; Oren-Shamir, M.; Aharoni, A.; Galili, G. Metabolic Engineering of the Phenylpropanoid and Its Primary, Precursor Pathway to Enhance the Flavor of Fruits and the Aroma of Flowers. Bioengineering 2015, 2, 204–212. [Google Scholar] [CrossRef]
- Ferreyra, M.L.F.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant. Sci. 2012, 3, 222. [Google Scholar]
- Singh, B.; Kumar, A.; Malik, A.K. Flavonoids biosynthesis in plants and its further analysis by capillary electrophoresis. Electrophoresis 2017, 38, 820–832. [Google Scholar] [CrossRef]
- Camila Gomez, G.C.; Laurent, T.; Véronique, C.; Nancy, T.; Agnès, A. In vivo grapevine anthocyanin transport involves vesicle-mediated trafficking and the contribution of anthoMATE transporters and GST. Plant J. 2011, 67, 960–970. [Google Scholar] [CrossRef]
- Jimenez-Garcia, S.N.V.; Moises, A.; Guevara-Gonzalez, R.G.; Torres-Pacheco, I.; Cruz-Hernandez, A.; Feregrino-Perez, A.A. Current Approaches for Enhanced Expression of Secondary Metabolites as Bioactive Compounds in Plants for Agronomic and Human Health Purposes—A Review. Polish J. Food Nutr. Sci. 2013, 63, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.L.; Wang, G.L.; Xiong, F.; Yu, X.R.; Xu, Z.S.; Wang, F.; Xiong, A.S. De novo assembly, transcriptome characterization, lignin accumulation, and anatomic characteristics: Novel insights into lignin biosynthesis during celery leaf development. Sci. Rep. 2015, 5, 8259. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.H.; Cao, T.J.; Dai, H.F.; Li, H.L.; Guo, D.; Mei, W.L.; Peng, S.Q. De Novo transcriptome characterization of Dracaena cambodiana and analysis of genes involved in flavonoid accumulation during formation of dragon’s blood. Sci. Rep. 2016, 6, 38315. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Kumar, S.; Bains, S.; Vaidya, V.; Singh, B.; Kaur, R.; Kaur, J.; Singh, K. De novo Transcriptome Analysis Revealed Genes Involved in Flavonoid and Vitamin C Biosynthesis in Phyllanthus emblica (L.). Front. Plant. Sci. 2016, 7, 1610. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S.; Goel, R.; Bhardwaj, A.; Asif, M.H.; Sawant, S.V.; Misra, P. Transcriptome analysis provides insight into prickle development and its link to defense and secondary metabolism in Solanum viarum Dunal. Sci. Rep. 2018, 8, 17092. [Google Scholar] [CrossRef]
- Lei, Z.; Zhou, C.; Ji, X.; Wei, G.; Huang, Y.; Yu, W.; Luo, Y.; Qiu, Y. Transcriptome Analysis Reveals genes involved in flavonoid biosynthesis and accumulation in Dendrobium catenatum From Different Locations. Sci. Rep. 2018, 8, 6373. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Yu, D.; Xue, J.; Lu, J.; Feng, S.; Shen, C.; Wang, H. A transcriptome-wide, organ-specific regulatory map of Dendrobium officinale, an important traditional Chinese orchid herb. Sci. Rep. 2016, 6, 18864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Yu, M.; Jia, Z.; Song, X.; Liang, Y.; Zhang, J. Analysis of Dendrobium huoshanense transcriptome unveils putative genes associated with active ingredients synthesis. BMC Genom. 2018, 19, 978. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.M.; Wang, R.; Li, X.D.; Jiang, Y.M.; Wang, C.Y.; Quan, J.P. Effect of abiotic and biotic elicitors on growth and alkaloid accumulation of Lycoris chinensis seedlings. Z. Naturforsch. C 2009, 64, 541–550. [Google Scholar] [CrossRef]
- Wang, H.; Yu, M.; Paek, K.Y.; Piao, X.C.; Lian, M.L. An efficient strategy for enhancement of bioactive compounds by protocorm-like body culture of Dendrobium candidum. Ind. Crop. Prod. 2016, 84, 112–130. [Google Scholar] [CrossRef]
- Zhang, X.N.; Liu, J.; Liu, Y.; Wang, Y.; Abozeid, A.; Yu, Z.G.; Tang, Z.H. Metabolomics Analysis Reveals that Ethylene and Methyl Jasmonate Regulate Different Branch Pathways to Promote the Accumulation of Terpenoid Indole Alkaloids in Catharanthus roseus. J. Nat. Prod. 2018, 81, 335–342. [Google Scholar] [CrossRef]
- Shen, C.; Guo, H.; Chen, H.; Shi, Y.; Meng, Y.; Lu, J.; Feng, S.; Wang, H. Identification and analysis of genes associated with the synthesis of bioactive constituents in Dendrobium officinale using RNA-Seq. Sci. Rep. 2017, 7, 187. [Google Scholar] [CrossRef] [Green Version]
- Schwab, W.; Wust, M. Understanding the Constitutive and Induced Biosynthesis of Mono- and Sesquiterpenes in Grapes (Vitis vinifera): A Key to Unlocking the Biochemical Secrets of Unique Grape Aroma Profiles. J. Agric. Food Chem. 2015, 63, 10591–10603. [Google Scholar] [CrossRef]
- Conesa, A.; Gotz, S. Blast2GO: A comprehensive suite for functional analysis in plant genomics. Int. J. Plant. Genom. 2008, 2008, 619832. [Google Scholar] [CrossRef]
- Ng, T.B.; Liu, J.; Wong, J.H.; Ye, X.; Wing Sze, S.C.; Tong, Y.; Zhang, K.Y. Review of research on Dendrobium, a prized folk medicine. Appl. Microbiol. Biotechnol. 2012, 93, 1795–1803. [Google Scholar] [CrossRef]
- Li, Y.; Li, F.; Gong, Q.; Wu, Q.; Shi, J. Inhibitory effects of Dendrobium alkaloids on memory impairment induced by lipopolysaccharide in rats. Planta Med. 2011, 77, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Ge, Y.H.; Wang, J.; Yang, F.; Dai, G.H.; Tong, Y.L. Effects of fresh Dendrobium officinale polysaccharides on immune function in mice with Lewis lung cancer. Zhejiang J. Tradit. Chin. Med. 2014, 49, 277–279. [Google Scholar]
- Li, M.; He, Y.; Peng, C.; Xie, X.; Hu, G. Erianin inhibits human cervical cancer cell through regulation of tumor protein p53 via the extracellular signal-regulated kinase signaling pathway. Oncol. Lett. 2018, 16, 5006–5012. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.Q.; Fan, Y.; Wu, D.Z.; Yang, H.; Hu, Z.B.; Wang, Z.T. In vivo and in vitro evaluation of erianin, a novel anti-angiogenic agent. Eur. J. Cancer 2004, 40, 1554–1565. [Google Scholar] [CrossRef]
- He, C.; Zhang, J.; Liu, X.; Zeng, S.; Wu, K.; Yu, Z.; Wang, X.; Teixeira da Silva, J.A.; Lin, Z.; Duan, J. Identification of genes involved in biosynthesis of mannan polysaccharides in Dendrobium officinale by RNA-seq analysis. Plant. Mol. Biol. 2015, 88, 219–231. [Google Scholar] [CrossRef]
- Charoenrungruang, S.; Chanvorachote, P.; Sritularak, B.; Pongrakhananon, V. Gigantol, a bibenzyl from Dendrobium draconis, inhibits the migratory behavior of non-small cell lung cancer cells. J. Nat. Prod. 2014, 77, 1359–1366. [Google Scholar] [CrossRef]
- Unahabhokha, T.; Chanvorachote, P.; Sritularak, B.; Kitsongsermthon, J.; Pongrakhananon, V. Gigantol Inhibits Epithelial to Mesenchymal Process in Human Lung Cancer Cells. Evid. Based Complementary Altern. Med. eCAM 2016, 2016, 4561674. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, Y.; Lyu, P.; Chen, L.; Shen, C.; Sun, C. Comparative transcriptomic analysis reveal the regulation mechanism underlying MeJA-induced accumulation of alkaloids in Dendrobium officinale. J. Plant. Res. 2019, 132, 419–429. [Google Scholar] [CrossRef]
- Ge, Q.; Zhang, Y.; Hua, W.P.; Wu, Y.C.; Jin, X.X.; Song, S.H.; Wang, Z.Z. Combination of transcriptomic and metabolomic analyses reveals a JAZ repressor in the jasmonate signaling pathway of Salvia miltiorrhiza. Sci. Rep. 2015, 5, 14048. [Google Scholar] [CrossRef] [Green Version]
- Cheong, J.J.; Choi, Y.D. Methyl jasmonate as a vital substance in plants. Trends Genet. 2003, 19, 409–413. [Google Scholar] [CrossRef]
- Guo, X.; Li, Y.; Li, C.; Luo, H.; Wang, L.; Qian, J.; Luo, X.; Xiang, L.; Song, J.; Sun, C.; et al. Analysis of the Dendrobium officinale transcriptome reveals putative alkaloid biosynthetic genes and genetic markers. Gene 2013, 527, 131–138. [Google Scholar] [CrossRef]
- Chen, Y.; Li, F.; Tian, L.; Huang, M.; Deng, R.; Li, X.; Chen, W.; Wu, P.; Li, M.; Jiang, H.; et al. The Phenylalanine Ammonia Lyase Gene LjPAL1 Is Involved in Plant Defense Responses to Pathogens and Plays Diverse Roles in Lotus japonicus-Rhizobium Symbioses. Mol. Plant. Microbe Interact. 2017, 30, 739–753. [Google Scholar] [CrossRef] [Green Version]
- Yanfang, Y.; Kaikai, Z.; Liying, Y.; Xing, L.; Ying, W.; Hongwei, L.; Qiang, L.; Duanfen, C.; Deyou, Q. Identification and characterization of MYC transcription factors in Taxus sp. Gene 2018, 675, 1–8. [Google Scholar] [CrossRef]
- De Boer, K.; Tilleman, S.; Pauwels, L.; Vanden Bossche, R.; De Sutter, V.; Vanderhaeghen, R.; Hilson, P.; Hamill, J.D.; Goossens, A. APETALA2/ETHYLENE RESPONSE FACTOR and basic helix-loop-helix tobacco transcription factors cooperatively mediate jasmonate-elicited nicotine biosynthesis. Plant J. 2011, 66, 1053–1065. [Google Scholar] [CrossRef]
- Toledo-Ortiz, G.; Huq, E.; Quail, P.H. The Arabidopsis basic/helix-loop-helix transcription factor family. Plant Cell 2003, 15, 1749–1770. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Fan, H.J.; Ling, H.Q. Genome-wide identification and characterization of the bHLH gene family in tomato. BMC Genom. 2015, 16, 9. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.J.; Wang, J.R. Global identification, structural analysis and expression characterization of bHLH transcription factors in wheat. BMC Plant Biol. 2017, 17, 90. [Google Scholar] [CrossRef] [Green Version]
- Niu, X.; Guan, Y.; Chen, S.; Li, H. Genome-wide analysis of basic helix-loop-helix ( bHLH) transcription factors in Brachypodium distachyon. BMC Genom. 2017, 18, 619. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Ran, F.; Dong, H.L.; Wen, J.; Li, J.N.; Liang, Z. Genome-Wide Analysis, Classification, Evolution, and Expression Analysis of the Cytochrome P450 93 Family in Land Plants. PLoS ONE 2016, 11, e0165020. [Google Scholar] [CrossRef] [Green Version]
- Anderson, N.A.; Bonawitz, N.D.; Nyffeler, K.; Chapple, C. Loss of FERULATE 5-HYDROXYLASE Leads to Mediator-Dependent Inhibition of Soluble Phenylpropanoid Biosynthesis in Arabidopsis. Plant Physiol. 2015, 169, 1557–1567. [Google Scholar] [CrossRef] [Green Version]
- Humphreys, J.M.; Hemm, M.R.; Chapple, C. New routes for lignin biosynthesis defined by biochemical characterization of recombinant ferulate 5-hydroxylase, a multifunctional cytochrome P450-dependent monooxygenase. Proc. Natl. Acad. Sci. USA 1999, 96, 10045–10050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, Y.; Shimada, T.; Motomura, Y.; Sato, F. Modulation of benzylisoquinoline alkaloid biosynthesis by heterologous expression of CjWRKY1 in Eschscholzia californica cells. PLoS ONE 2017, 12, e0186953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, A.; Trapnell, C.; Donaghey, J.; Rinn, J.L.; Pachter, L. Improving RNA-Seq expression estimates by correcting for fragment bias. Gen. Biol. 2011, 12, R22. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Huber, W. Differential Expression of RNA-Seq Data at the Gene Level—The DESeq Package; European Molecular Biology Laboratory (EMBL): Heidelberg, Germany, 2012; Volume 23. [Google Scholar]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Gen. Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Nelson, D.R.; Koymans, L.; Kamataki, T.; Stegeman, J.J.; Feyereisen, R.; Waxman, D.J.; Waterman, M.R.; Gotoh, O.; Coon, M.J.; Estabrook, R.W.; et al. P450 superfamily: Update on new sequences, gene mapping, accession numbers and nomenclature. Pharmacogenetics 1996, 6, 1–42. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Erianin (µg/g) | Gigantol (µg/g) | Total Content (µg/g) |
|---|---|---|---|
| Roots | 2.63 ± 0.69 | 37.01 ± 2.16 | 39.64 ± 4.14 |
| basal stems | 0.61 ± 0.01 | 22.67 ± 0.15 | 23.28 ± 0.25 |
| upper stems | ND | 12.15 ± 1.87 | 12.15 ± 1.87 |
| Leaves | ND | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adejobi, O.I.; Guan, J.; Yang, L.; Hu, J.-M.; Yu, A.; Muraguri, S.; Liu, A. Transcriptomic Analyses Shed Light on Critical Genes Associated with Bibenzyl Biosynthesis in Dendrobium officinale. Plants 2021, 10, 633. https://doi.org/10.3390/plants10040633
Adejobi OI, Guan J, Yang L, Hu J-M, Yu A, Muraguri S, Liu A. Transcriptomic Analyses Shed Light on Critical Genes Associated with Bibenzyl Biosynthesis in Dendrobium officinale. Plants. 2021; 10(4):633. https://doi.org/10.3390/plants10040633
Chicago/Turabian StyleAdejobi, Oluwaniyi Isaiah, Ju Guan, Liu Yang, Jiang-Miao Hu, Anmin Yu, Sammy Muraguri, and Aizhong Liu. 2021. "Transcriptomic Analyses Shed Light on Critical Genes Associated with Bibenzyl Biosynthesis in Dendrobium officinale" Plants 10, no. 4: 633. https://doi.org/10.3390/plants10040633
APA StyleAdejobi, O. I., Guan, J., Yang, L., Hu, J. -M., Yu, A., Muraguri, S., & Liu, A. (2021). Transcriptomic Analyses Shed Light on Critical Genes Associated with Bibenzyl Biosynthesis in Dendrobium officinale. Plants, 10(4), 633. https://doi.org/10.3390/plants10040633