
Meta-Analysis of Quantitative Traits Loci (QTL) Identified in Drought Response in Rice (Oryza sativa L.)


Abstract
:1. Introduction
2. Results and Discussion
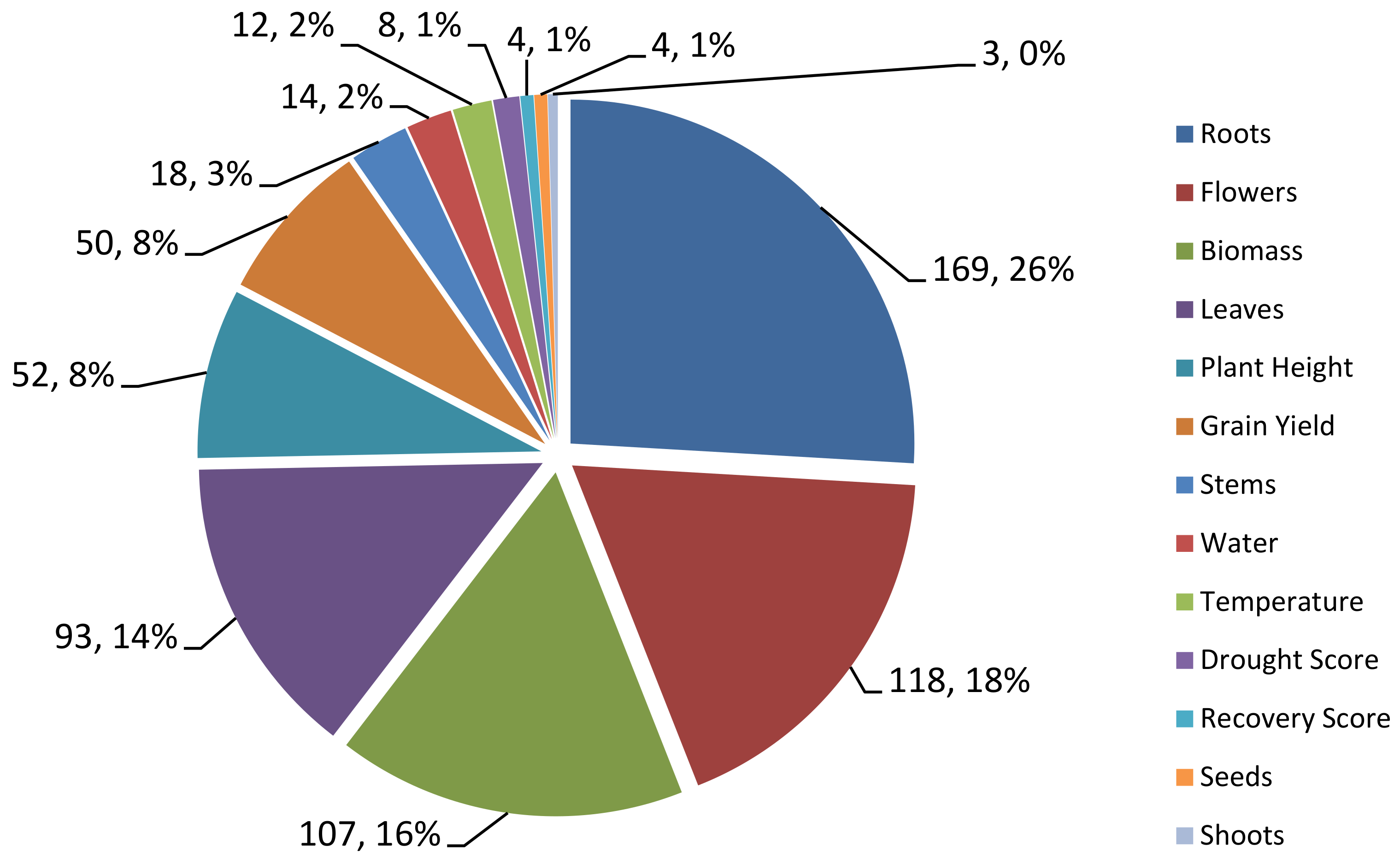
2.1. Collection of QTL Data Related to Drought Tolerance in Rice from Past Studies
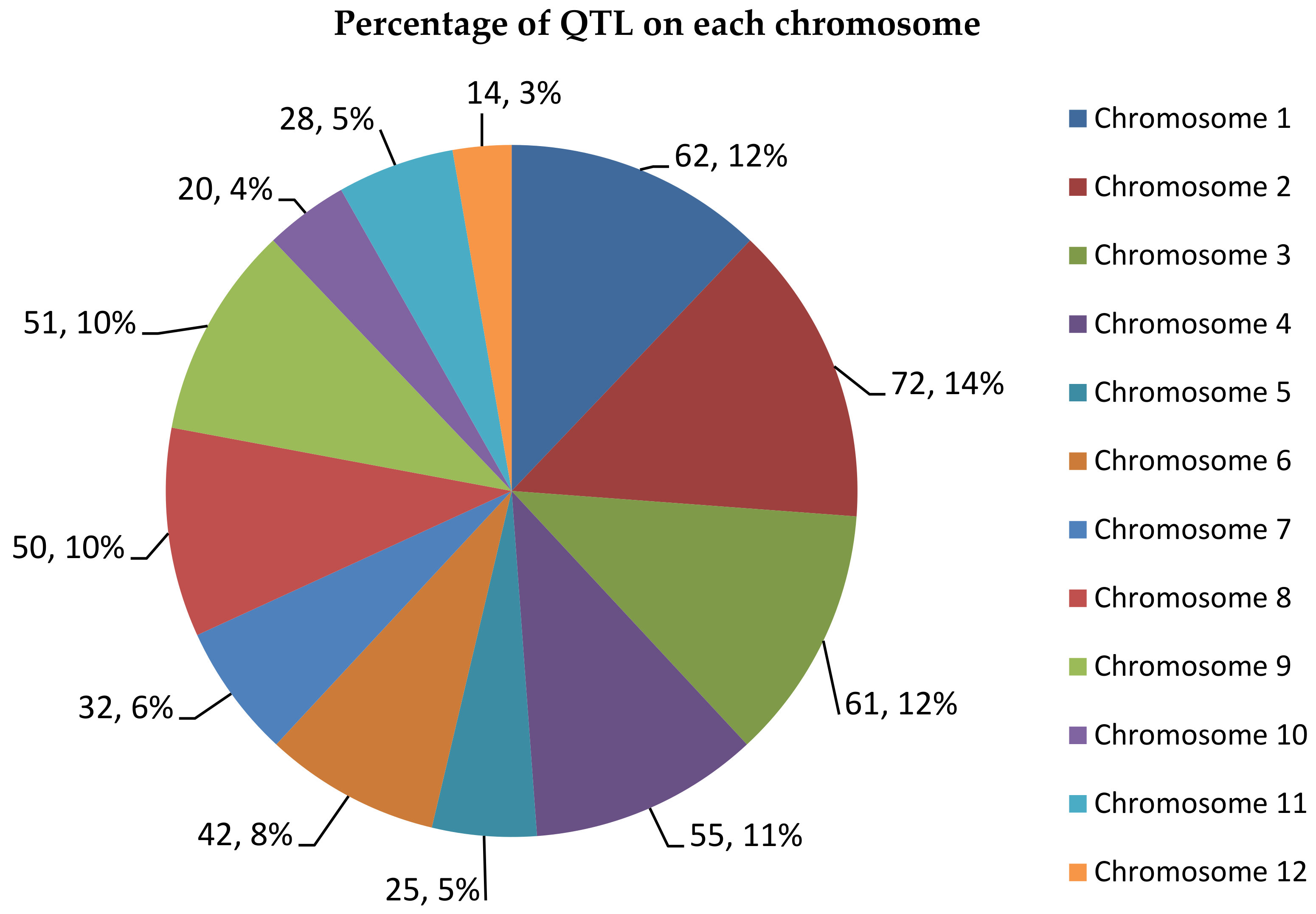
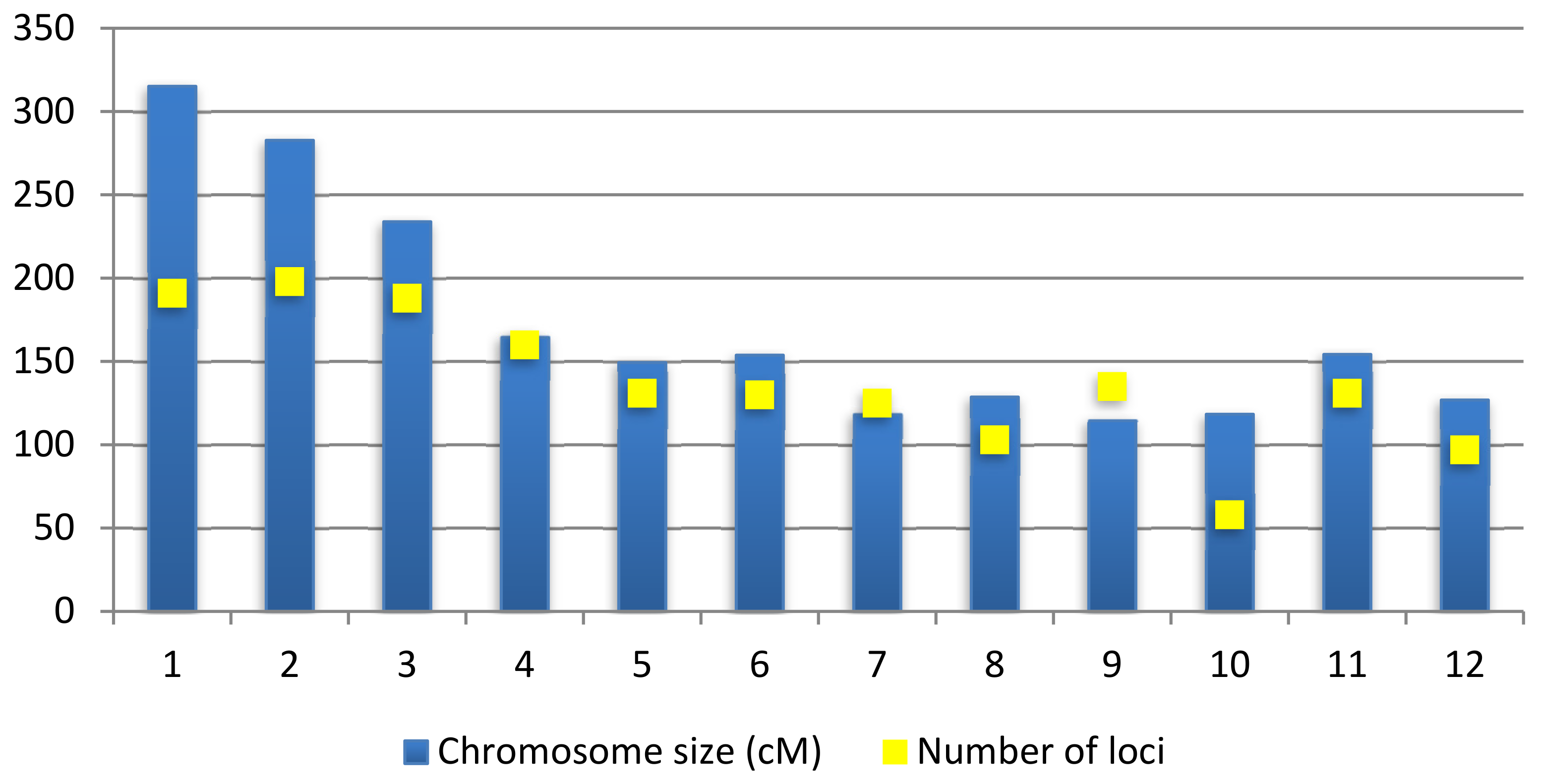
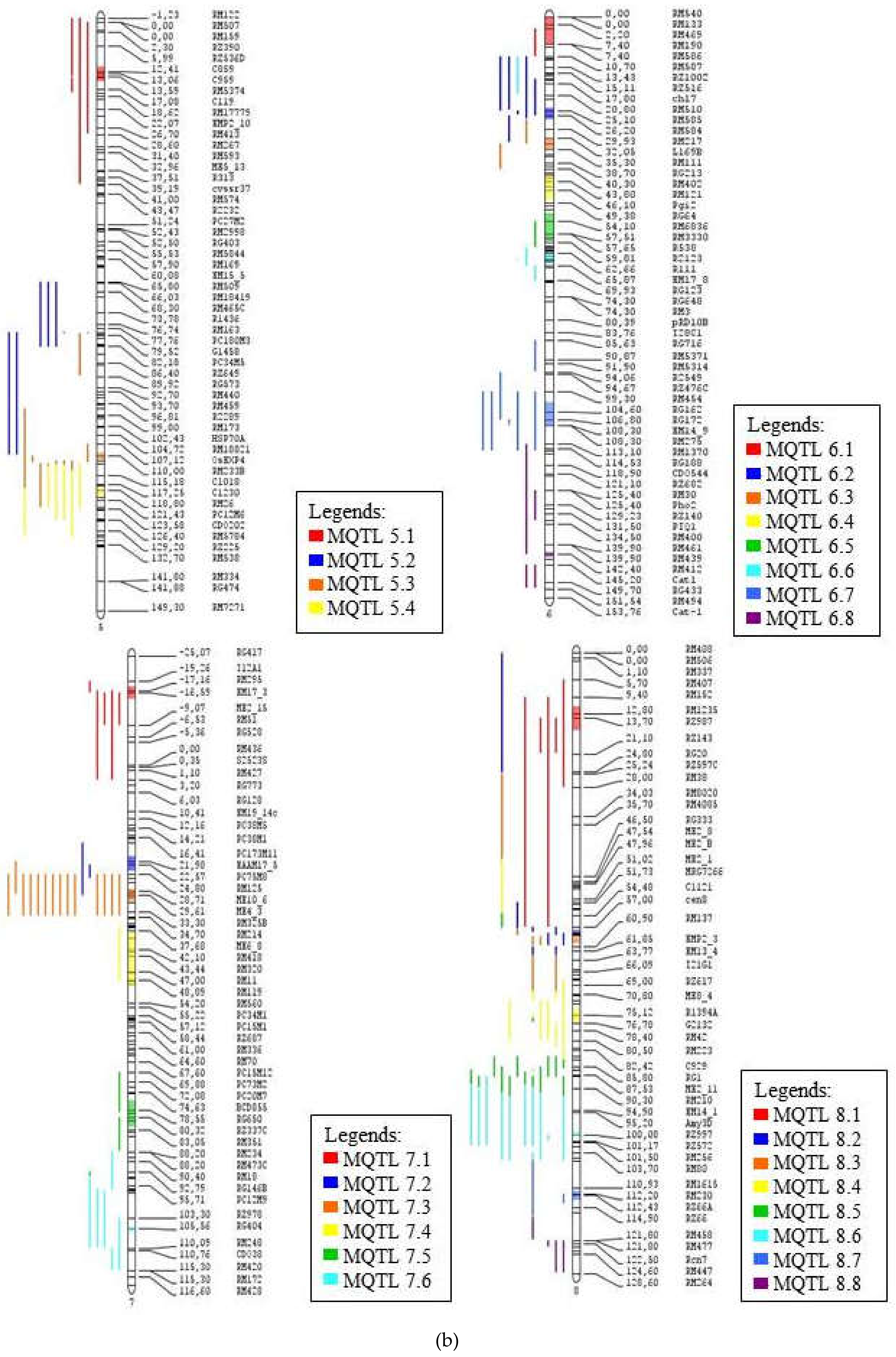
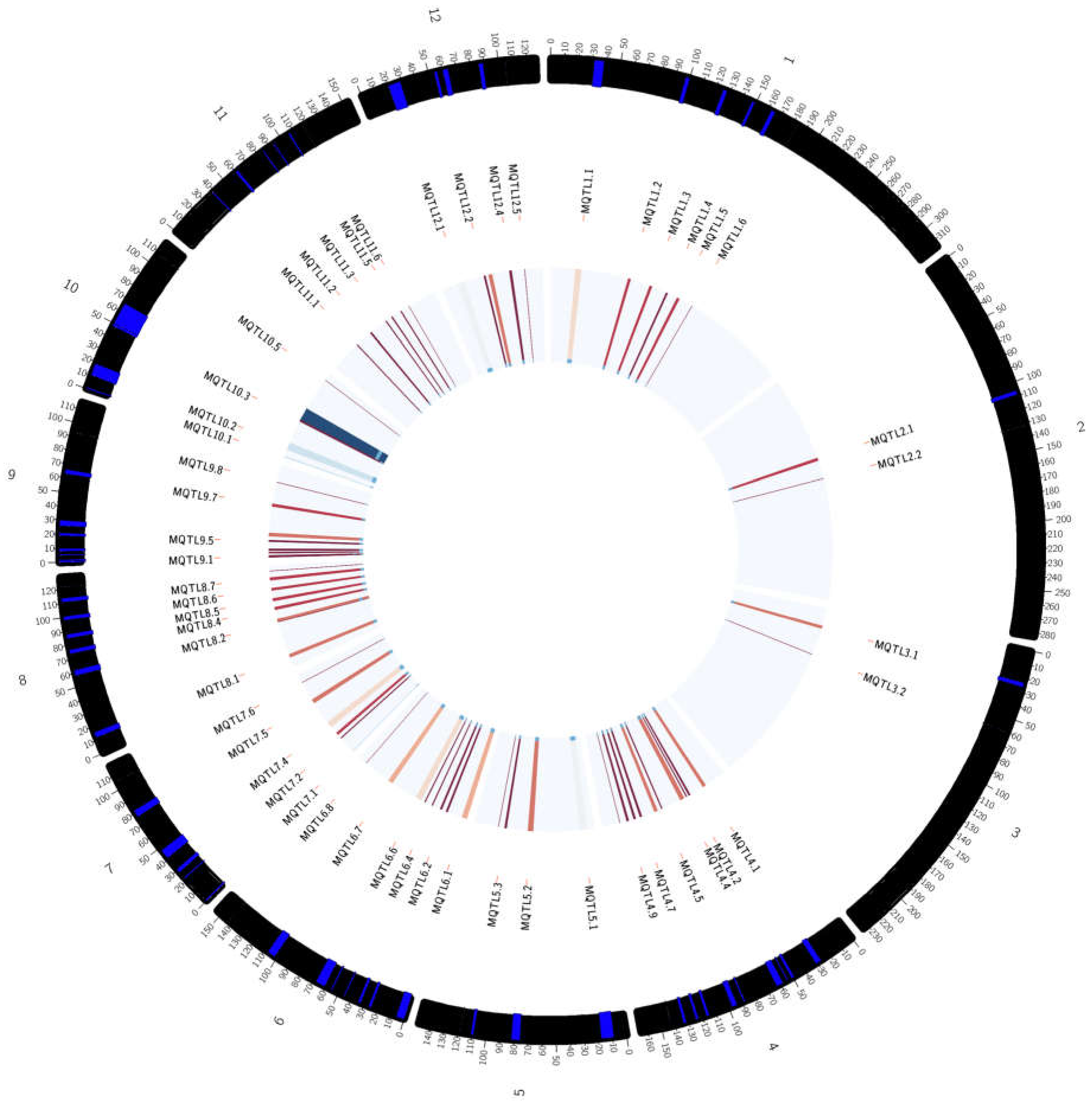
2.2. Generation of Consensus Map and Projection of QTLs
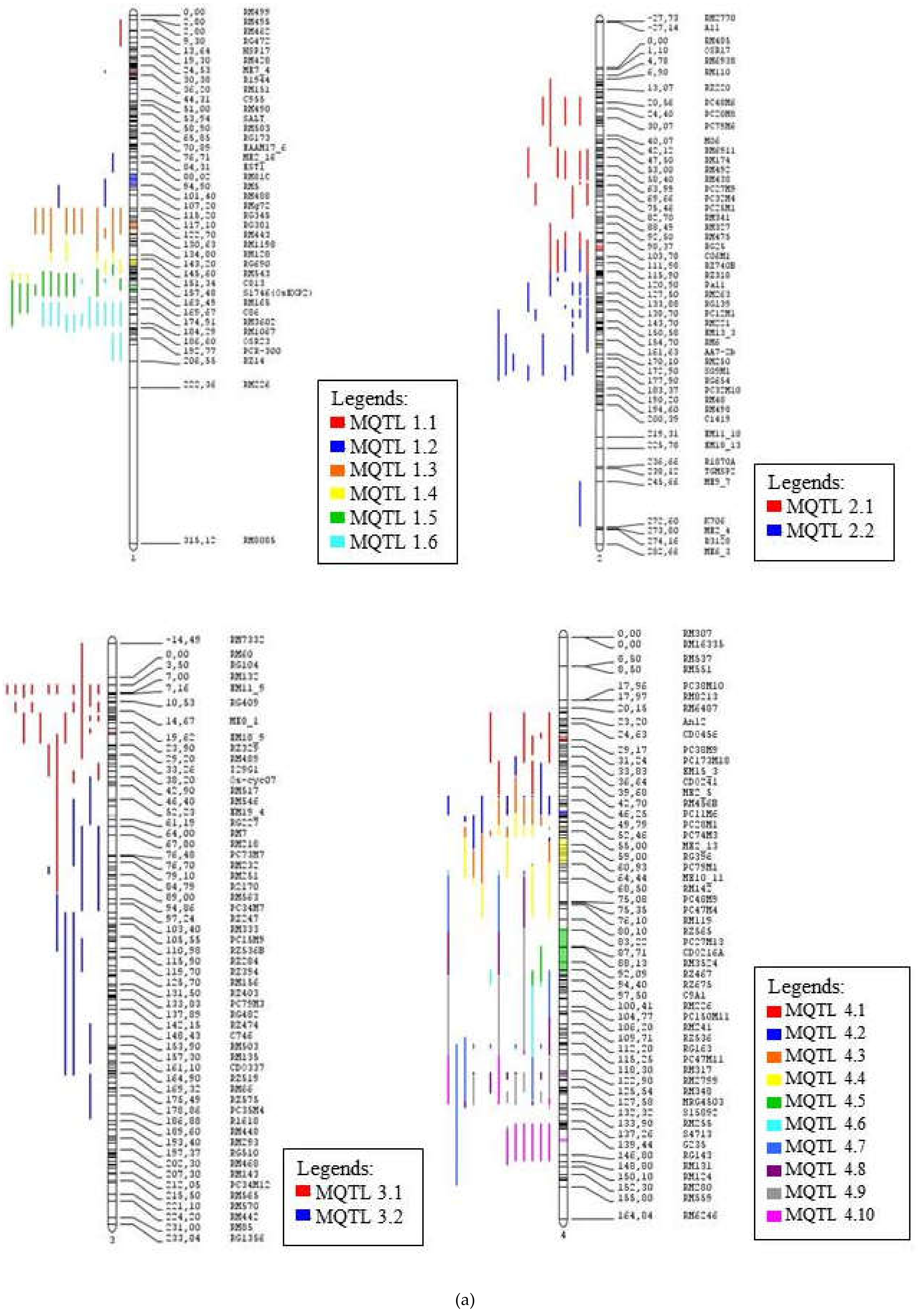
2.3. Meta-QTL Analysis for Frought Tolerance in Rice
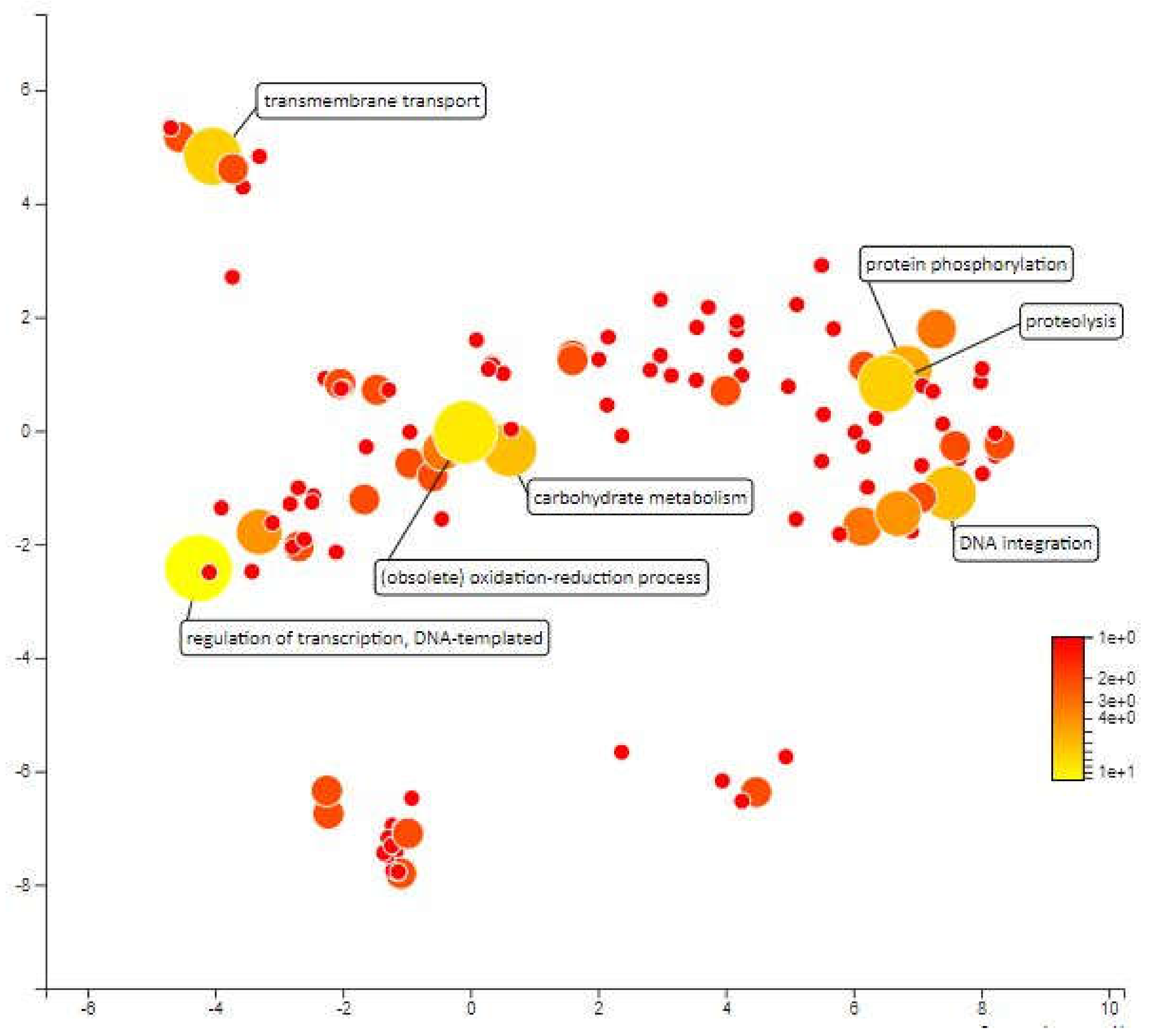
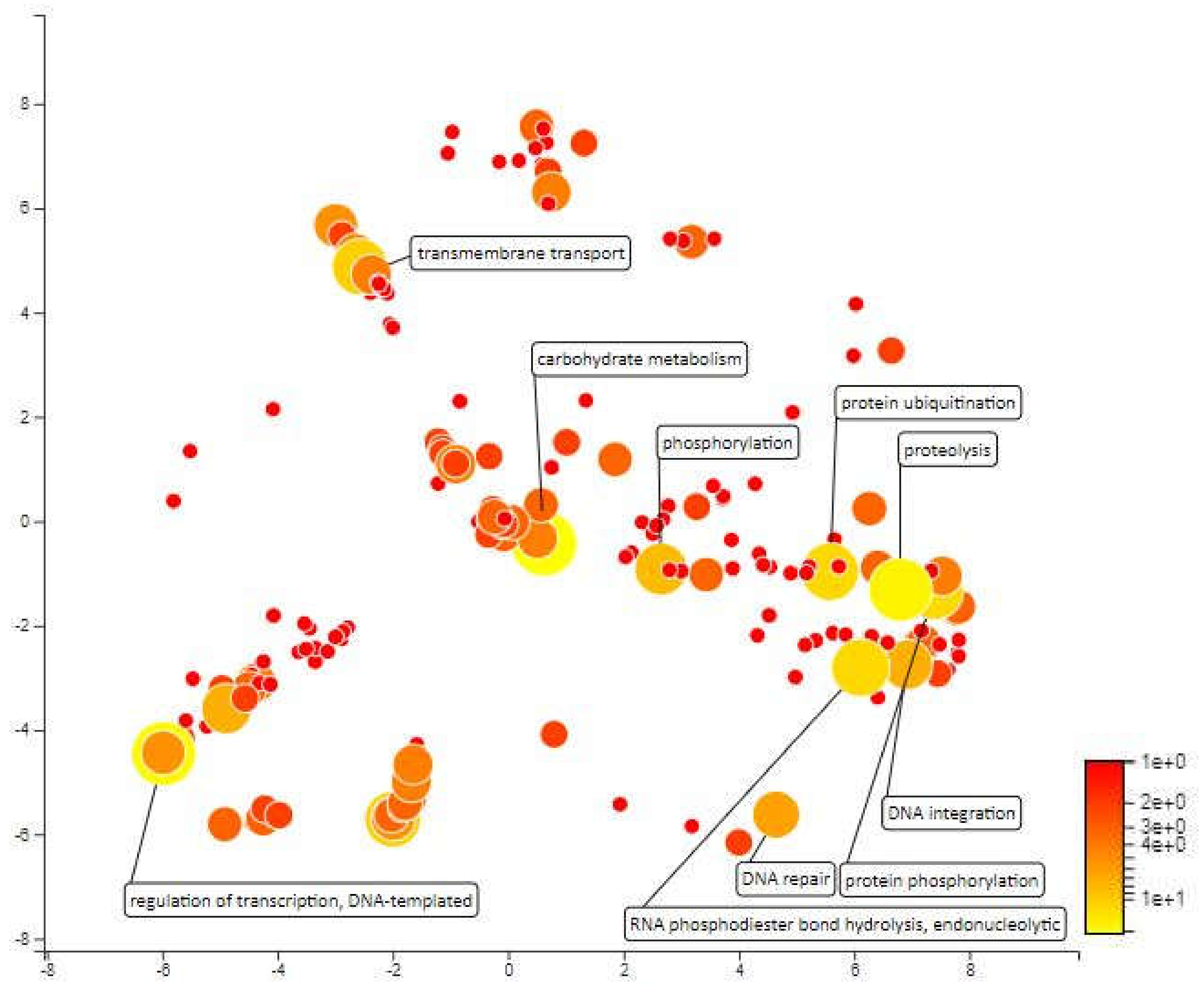
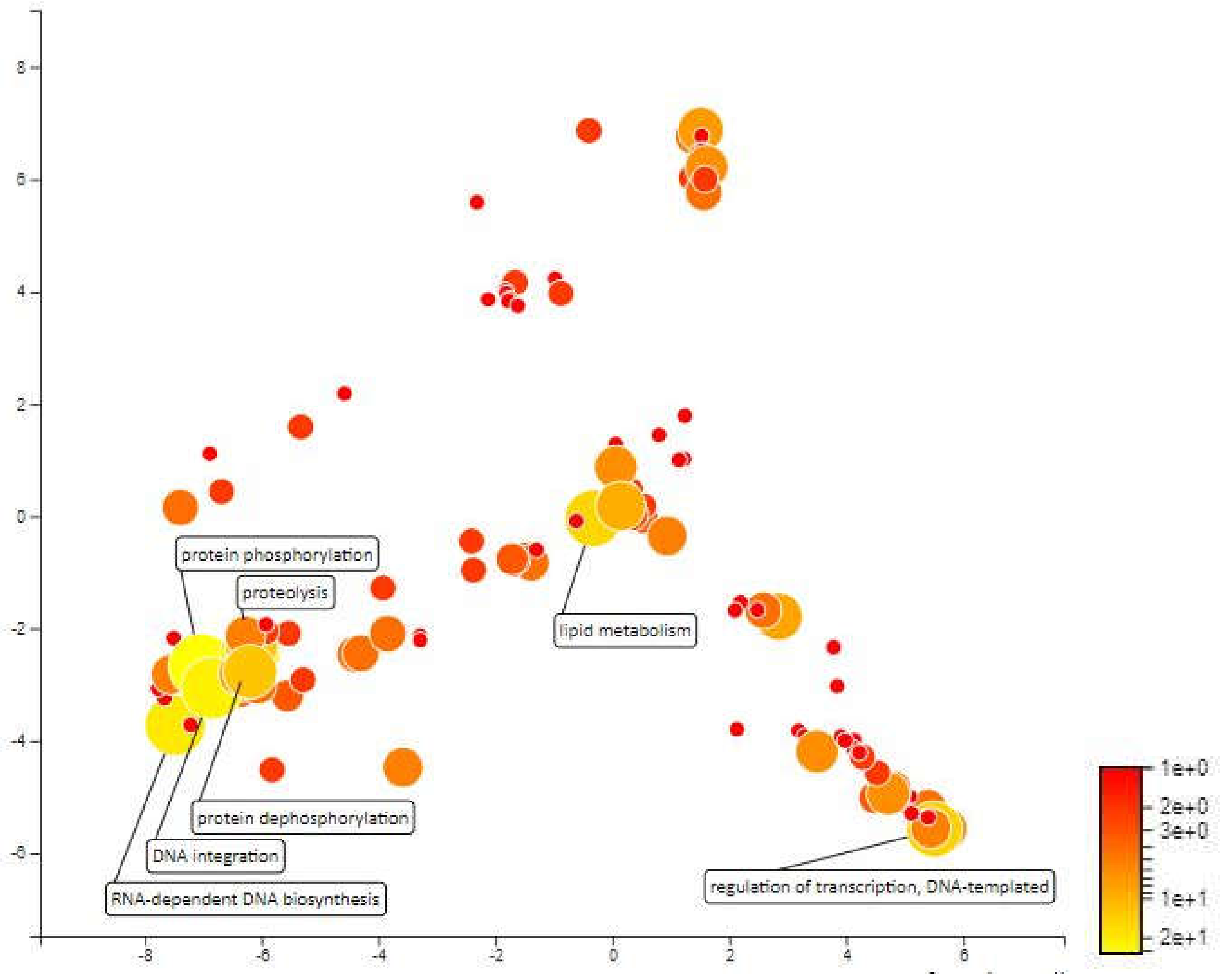
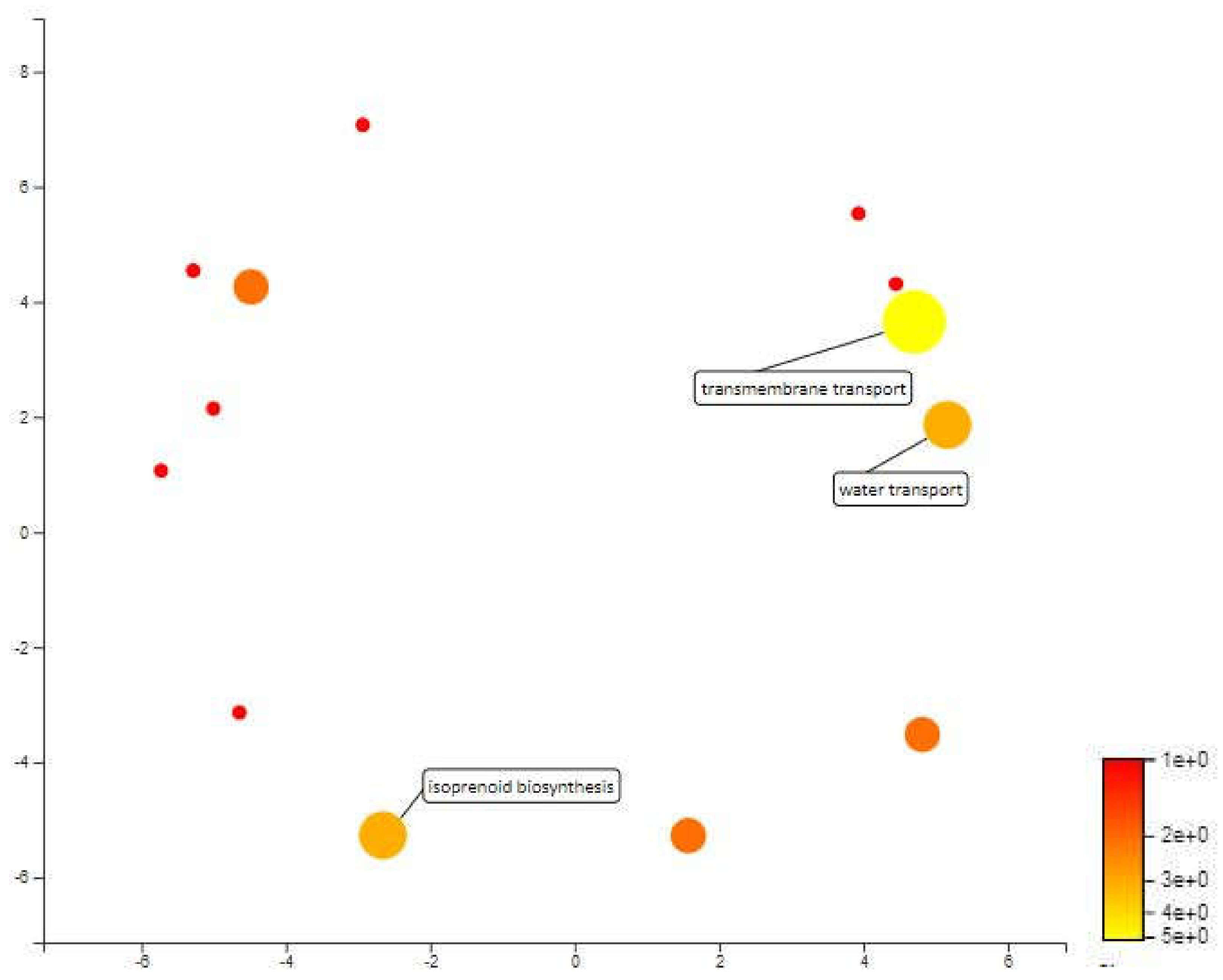
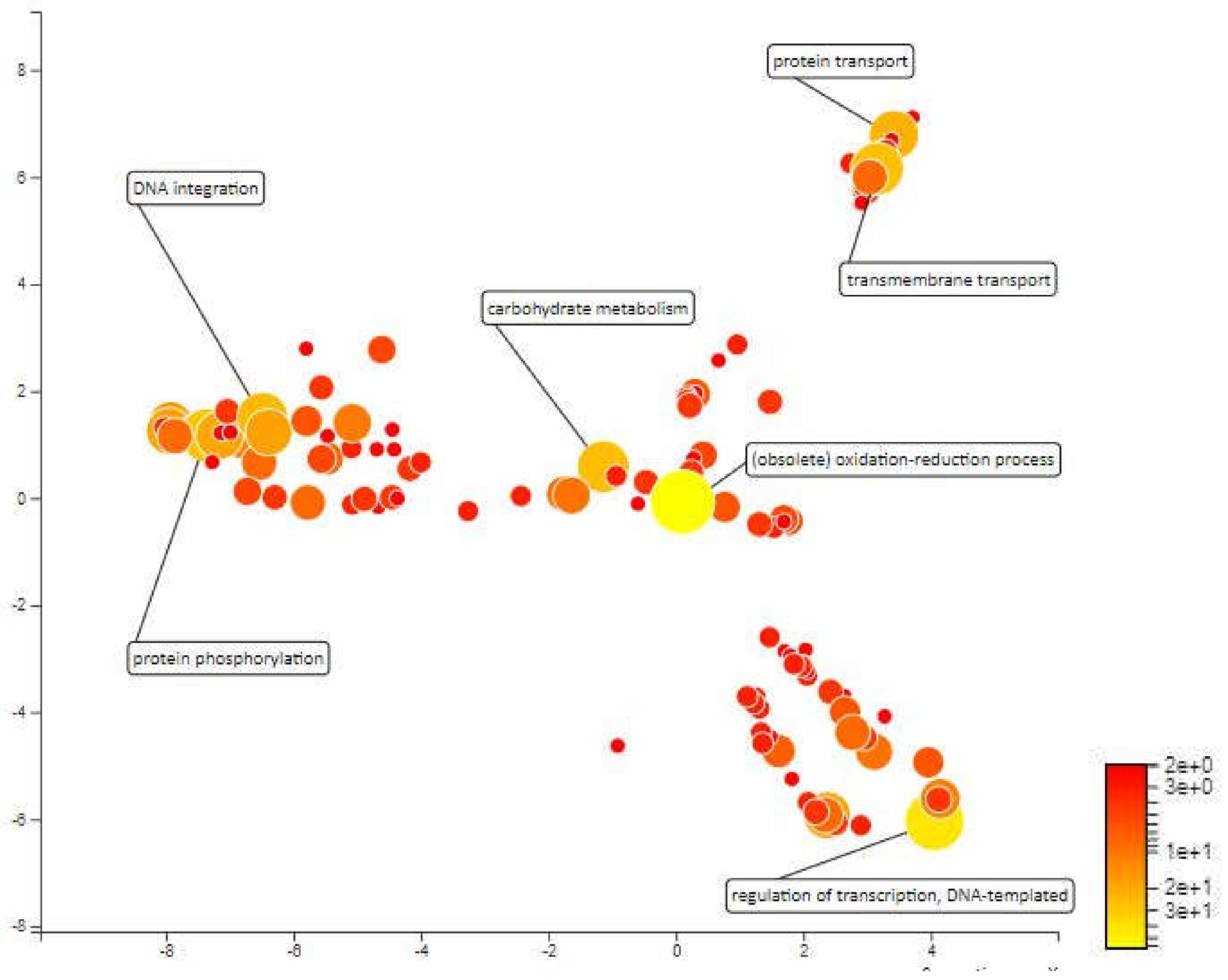
2.4. Functional Annotation for Candidate Meta-QTL
2.4.1. MQTL 1.5
2.4.2. MQTL 1.6
2.4.3. MQTL 2.1
2.4.4. MQTL 2.2
2.4.5. MQTL 3.1
3. Materials and Methods
3.1. Bibliographic Review and Data Synthesis
3.2. Consensus Map and QTL Projection
3.3. Meta-QTL Analysis
3.4. Functional Annotaion
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gosal, S.S.; Wani, S.H.; Kang, M.S. Biotechnology and Drought Tolerance. J. Crop Improv. 2009, 23, 19–54. [Google Scholar] [CrossRef]
- Baldwin, K.; Childs, N.; Dyck, J.; Hansen, J. Southeast Asia’s Rice Surplus. Econ. Res. Serv. 2012, RCS-121-01, 1–38. [Google Scholar]
- Che Omar, S.; Ahmad Shaharudin, A.A.; Tumin, S.A. The Status of the Paddy and Rice Industry in Malaysia; Khazanah Research Institute: Kuala Lumpur, Malaysia, 2019. [Google Scholar]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Asfaliza, R.; Latif, M.A. Blast resistance in rice: A review of conventional breeding to molecular approaches. Mol. Biol. Rep. 2013, 40, 2369–2388. [Google Scholar] [CrossRef]
- Chukwu, S.C.; Rafii, M.Y.; Ramlee, S.I.; Ismail, S.I.; Hasan, M.M.; Oladosu, Y.A.; Magaji, U.G.; Akos, I.; Olalekan, K.K. Bacterial leaf blight resistance in rice: A review of conventional breeding to molecular approach. Mol. Biol. Rep. 2019, 46, 1519–1532. [Google Scholar] [CrossRef] [PubMed]
- Singla, J.; Krattinger, S.G. Biotic Stress Resistance Genes in Wheat; Wrigley, C., Corke, H., Seetharaman, K., Faubion, J.B.T.-E., Eds.; Academic Press: Oxford, UK, 2016; pp. 388–392. ISBN 978-0-12-394786-4. [Google Scholar]
- Kumar, I.S.; Zaharin, N.; Nadarajah, K. In silico Identification of Resistance and Defense Related Genes for Bacterial Leaf Blight (BLB) in Rice. J. Pure Appl. Microbiol. 2018, 12, 1867–1876. [Google Scholar] [CrossRef] [Green Version]
- Kumar, I.S.; Amran, N.A.; Nadarajah, K. In silico Analysis of qBFR4 and qLBL5 in Conferring Quantitative Resistance Against Rice Blast. J. Pure Appl. Microbiol. 2018, 12, 1703–1718. [Google Scholar] [CrossRef]
- Kumar, I.S.; Nadarajah, K. A Meta-Analysis of Quantitative Trait Loci Associated with Multiple Disease Resistance in Rice (Oryza sativa L.). Plants 2020, 9, 1491. [Google Scholar] [CrossRef]
- Ben-Ari, G.; Lavi, U. Marker-assisted selection in plant breeding. Plant Biotechnol. Agric. 2012, 163–184. [Google Scholar]
- Gosal, S.S.; Wani, S.H.; Kang, M.S. Biotechnology and Crop Improvement. J. Crop Improv. 2010, 24, 153–217. [Google Scholar] [CrossRef]
- Singhal, P.; Jan, A.T.; Azam, M.; Haq, Q.M.R. Plant abiotic stress: A prospective strategy of exploiting promoters as alternative to overcome the escalating burden. Front. Life Sci. 2016, 9, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Todaka, D.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Recent advances in the dissection of drought-stress regulatory networks and strategies for development of drought-tolerant transgenic rice plants. Front. Plant Sci. 2015, 6, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nearly 6000 rice farmers affected by drought. The Star, 18 July 2016.
- Benjamin, J.G.; Nielsen, D.C. Water deficit effects on root distribution of soybean, field pea and chickpea. F. Crop. Res. 2006, 97, 248–253. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Samuel, C.; Fatai, A.; Magaji, U. Drought Resistance in Rice from Conventional to Molecular Breeding: A Review. Int. J. Mol. Sci. 2019, 20, 3519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wani, S.H.; Sah, S.K. Biotechnology and Abiotic Stress Tolerance in Rice. J. Rice Res. 2014, 2. [Google Scholar] [CrossRef]
- Solis, J.; Gutierrez, A.; Mangu, V.; Sánchez Timm, E.; Bedre, R.; Linscombe, S.; Baisakh, N. Genetic mapping of quantitative trait loci for grain yield under drought in rice under controlled greenhouse conditions. Front. Chem. 2018, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanceras, J.C.; Pantuwan, G.; Jongdee, B.; Toojinda, T. Quantitative Trait Loci Associated with Drought Tolerance at Reproductive Stage in Rice. Plant Physiol. 2004, 135, 384–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collard, B.C.Y.; Jahufer, M.Z.Z.; Brouwer, J.B.; Pang, E.C.K. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Zhang, X.; Shabala, S.; Koutoulis, A.; Shabala, L.; Zhou, M. Meta-analysis of major QTL for abiotic stress tolerance in barley and implications for barley breeding. Planta 2017, 245, 283–295. [Google Scholar] [CrossRef]
- Khush, G.S. Green revolution: The way forward. Nat. Rev. Genet. 2001, 2, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Dixit, S.; Ram, T.; Yadaw, R.B.; Mishra, K.K.; Mandal, N.P. Breeding high-yielding drought-tolerant rice: Genetic variations and conventional and molecular approaches. J. Exp. Bot. 2014, 65, 6265–6278. [Google Scholar] [CrossRef] [Green Version]
- Sahebi, M.; Hanafi, M.M.; Rafii, M.Y.; Mahmud, T.M.M.; Azizi, P.; Osman, M.; Abiri, R.; Taheri, S.; Kalhori, N.; Shabanimofrad, M.; et al. Improvement of Drought Tolerance in Rice (Oryza sativa L.): Genetics, Genomic Tools, and the WRKY Gene Family. Biomed Res. Int. 2018, 2018, 3158474. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Chung, Y.S.; Lee, E.; Tripathi, P.; Heo, S.; Kim, K.-H. Root Response to Drought Stress in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2020, 21, 1513. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Chen, C.; Ma, X.; Zhang, Y.; Han, J.; Mei, H.; Yu, S. Comparative Analysis of Expression Profiles of Panicle Development among Tolerant and Sensitive Rice in Response to Drought Stress. Front. Plant Sci. 2017, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyaya, H.; Panda, S.K. Chapter 9-Drought Stress Responses and Its Management in Rice. In Advances in Rice Research for Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Nahar, K., Biswas, J.K., Eds.; Woodhead Publishing: Cambridge, UK, 2019; pp. 177–200. ISBN 978-0-12-814332-2. [Google Scholar] [CrossRef]
- Ali, M.L.; Pathan, M.S.; Zhang, J.; Bai, G.; Sarkarung, S.; Nguyen, H.T. Mapping QTLs for root traits in a recombinant inbred population from two indica ecotypes in rice. Theor. Appl. Genet. 2000, 101, 756–766. [Google Scholar] [CrossRef]
- Bernier, J.; Kumar, A.; Ramaiah, V.; Spaner, D.; Atlin, G. A large-effect QTL for grain yield under reproductive-stage drought stress in upland rice. Crop Sci. 2007, 47, 507–518. [Google Scholar] [CrossRef]
- Courtois, B.; Shen, L.; Petalcorin, W.; Carandang, S.; Mauleon, R.; Li, Z. Locating QTLs controlling constitutive root traits in the rice population IAC 165 x Co39. Euphytica 2003, 134, 335–345. [Google Scholar] [CrossRef]
- Gomez, S.M.; Boopathi, N.M.; Kumar, S.S.; Ramasubramanian, T.; Chengsong, Z.; Jeyaprakash, P.; Senthil, A.; Babu, R.C. Molecular mapping and location of QTLs for drought-resistance traits in indica rice (Oryza sativa L.) lines adapted to target environments. Acta Physiol. Plant. 2010, 32, 355–364. [Google Scholar] [CrossRef]
- Gu, J.; Yin, X.; Struik, P.C.; Stomph, T.J.; Wang, H. Using chromosome introgression lines to map quantitative trait loci for photosynthesis parameters in rice (Oryza sativa L.) leaves under drought and well-watered field conditions. J. Exp. Bot. 2012, 63, 455–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.; Hirotsu, S.; Nemoto, K.; Yamagishi, J. Identification of QTLs controlling rice drought tolerance at seedling stage in hydroponic culture. Euphytica 2008, 160, 423–430. [Google Scholar] [CrossRef]
- Kumar, R.; Venuprasad, R.; Atlin, G.N. Genetic analysis of rainfed lowland rice drought tolerance under naturally-occurring stress in eastern India: Heritability and QTL effects. F. Crop. Res. 2007, 103, 42–52. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Klueva, N.; Chamareck, V.; Aarti, A.; Magpantay, G.; Millena, A.C.M.; Pathan, M.S.; Nguyen, H.T. Saturation mapping of QTL regions and identification of putative candidate genes for drought tolerance in rice. Mol. Genet. Genomics 2004, 272, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Palanog, A.D.; Swamy, B.P.M.; Shamsudin, N.A.A.; Dixit, S.; Hernandez, J.E.; Boromeo, T.H.; Cruz, P.C.S.; Kumar, A. Grain yield QTLs with consistent-effect under reproductive-stage drought stress in rice. F. Crop. Res. 2014, 161, 46–54. [Google Scholar] [CrossRef]
- Prince, S.J.; Beena, R.; Gomez, S.M.; Senthivel, S.; Babu, R.C. Mapping Consistent Rice (Oryza sativa L.) Yield QTLs under Drought Stress in Target Rainfed Environments. Rice 2015, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Sabar, M.; Shabir, G.; Shah, S.M.; Aslam, K.; Naveed, S.A.; Arif, M. Identification and mapping of QTLs associated with drought tolerance traits in rice by a cross between super Basmati and IR55419-04. Breed. Sci. 2019, 69, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, N.; Jain, S.; Kumar, A.; Mehla, B.S.; Jain, R. Genetic variation, linkage mapping of QTL and correlation studies for yield, root, and agronomic traits for aerobic adaptation. BMC Genet. 2013, 14, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, J.; Li, Q.; Yue, B.; Xue, W.Y.; Luo, L.J.; Xiong, L.Z. Identification of quantitative trait loci for ABA sensitivity at seed germination and seedling stages in rice. Acta Genet. Sin. 2006, 33, 532–541. [Google Scholar] [CrossRef]
- Yue, B.; Xue, W.; Luo, L.; Xing, Y. Identification of quantitative trait loci for four morphologic traits under water stress in rice (Oryza sativa L.). J. Genet. Genomics 2008, 35, 569–575. [Google Scholar] [CrossRef]
- Yue, B.; Xue, W.; Xiong, L.; Yu, X.; Luo, L.; Cui, K.; Jin, D.; Xing, Y.; Zhang, Q. Genetic basis of drought resistance at reproductive stage in rice: Separation of drought tolerance from drought avoidance. Genetics 2006, 172, 1213–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, B.; Xiong, L.; Xue, W.; Xing, Y.; Luo, L.; Xu, C. Genetic analysis for drought resistance of rice at reproductive stage in field with different types of soil. Theor. Appl. Genet. 2005, 111, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zheng, H.G.; Aarti, A.; Pantuwan, G.; Nguyen, T.T.; Tripathy, J.N.; Sarial, A.K.; Robin, S.; Babu, R.C.; Nguyen, B.D.; et al. Locating genomic regions associated with components of drought resistance in rice: Comparative mapping within and across species. Theor. Appl. Genet. 2001, 103, 19–29. [Google Scholar] [CrossRef]
- Zheng, B.; Yang, L.; Mao, C.; Huang, Y.; Wu, P. Comparison of QTLs for rice seedling morphology under different water supply conditions. J. Genet. Genomics 2008, 35, 473–484. [Google Scholar] [CrossRef]
- Swamy, B.M.; Vikram, P.; Dixit, S.; Ahmed, H.U.; Kumar, A. Meta-analysis of grain yield QTL identified during agricultural drought in grasses showed consensus. BMC Genomics 2011, 12, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khowaja, F.S.; Norton, G.J.; Courtois, B.; Price, A.H. Improved resolution in the position of drought-related QTLs in a single mapping population of rice by meta-analysis. BMC Genomics 2009, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saikumar, S.; Kalmeshwer Gouda, P.; Saiharini, A.; Varma, C.M.K.; Vineesha, O.; Padmavathi, G.; Shenoy, V.V. Major QTL for enhancing rice grain yield under lowland reproductive drought stress identified using an O. sativa/O. glaberrima introgression line. F. Crop. Res. 2014, 163, 119–131. [Google Scholar] [CrossRef]
- Vikram, P.; Swamy, B.P.M.; Dixit, S.; Ahmed, H.U.; Teresa Sta Cruz, M.; Singh, A.K.; Kumar, A. qDTY1.1, a major QTL for rice grain yield under reproductive-stage drought stress with a consistent effect in multiple elite genetic backgrounds. BMC Genet. 2011, 12, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohd Ikmal, A.; Nurasyikin, Z.; Tuan Nur Aqlili, R.T.A.; Puteri Dinie, E.Z.; Wickneswari, R.; Noraziyah, A.A.S. Drought Yield QTL (qDTY) with Consistent Effects on Morphological and Agronomical Traits of Two Populations of New Rice (Oryza sativa) Lines. Plants (Basel, Switzerland) 2019, 8, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamsudin, N.A.A.; Swamy, B.P.M.; Ratnam, W.; Cruz, S.M.T.; Raman, A.; Kumar, A. Marker assisted pyramiding of drought yield QTLs into a popular Malaysian rice cultivar, MR219. BMC Genet. 2016, 17, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grudkowska, M.; Zagdańska, B. Multifunctional role of plant cysteine proteinases. Acta Biochim. Pol. 2004, 51, 609–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simova-Stoilova, L.; Vaseva, I.; Grigorova, B.; Demirevska, K.; Feller, U. Proteolytic activity and cysteine protease expression in wheat leaves under severe soil drought and recovery. Plant Physiol. Biochem. 2010, 48, 200–206. [Google Scholar] [CrossRef]
- Jarzyniak, K.M.; Jasiński, M. Membrane transporters and drought resistance–a complex issue. Front. Plant Sci. 2014, 5, 687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabarty, A.; Aditya, M.; Dey, N.; Banik, N.; Bhattacharjee, S. Antioxidant Signaling and Redox Regulation in Drought- and Salinity-Stressed Plants. In Drought Stress Tolerance in Plants, Vol 1: Physiology and Biochemistry; Hossain, M.A., Wani, S.H., Bhattacharjee, S., Burritt, D.J., Tran, L.-S.P., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 465–498. ISBN 978-3-319-28899-4. [Google Scholar] [CrossRef]
- Skubacz, A.; Daszkowska-Golec, A.; Szarejko, I. The Role and Regulation of ABI5 (ABA-Insensitive 5) in Plant Development, Abiotic Stress Responses and Phytohormone Crosstalk. Front. Plant Sci. 2016, 7, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, A.; Daszkowska-Golec, A.; Kurowska, M.; Szarejko, I. Barley ABI5 (Abscisic Acid INSENSITIVE 5) Is Involved in Abscisic Acid-Dependent Drought Response. Front. Plant Sci. 2020, 11, 1138. [Google Scholar] [CrossRef] [PubMed]
- Menkens, A.E.; Schindler, U.; Cashmore, A.R. The G-box: A ubiquitous regulatory DNA element in plants bound by the GBF family of bZIP proteins. Trends Biochem. Sci. 1995, 20, 506–510. [Google Scholar] [CrossRef]
- Ramegowda, V.; Gill, U.S.; Sivalingam, P.N.; Gupta, A.; Gupta, C.; Govind, G.; Nataraja, K.N.; Pereira, A.; Udayakumar, M.; Mysore, K.S.; et al. GBF3 transcription factor imparts drought tolerance in Arabidopsis thaliana. Sci. Rep. 2017, 7, 9148. [Google Scholar] [CrossRef] [PubMed]
- Dien, D.C.; Thu, T.T.P.; Moe, K.; Yamakawa, T. Proline and carbohydrate metabolism in rice varieties (Oryza sativa L.) under various drought and recovery conditions. Plant Physiol. Reports 2019, 24, 376–387. [Google Scholar] [CrossRef]
- Lyzenga, W.J.; Stone, S.L. Abiotic stress tolerance mediated by protein ubiquitination. J. Exp. Bot. 2012, 63, 599–616. [Google Scholar] [CrossRef] [Green Version]
- Benjamins, R.; Quint, A.; Weijers, D.; Hooykaas, P.; Offringa, R. The PINOID protein kinase regulates organ development in Arabidopsis by enhancing polar auxin transport. Development 2001, 128, 4057–4067. [Google Scholar]
- Nongpiur, R.; Soni, P.; Karan, R.; Singla-Pareek, S.L.; Pareek, A. Histidine kinases in plants: Cross talk between hormone and stress responses. Plant Signal. Behav. 2012, 7, 1230–1237. [Google Scholar] [CrossRef] [Green Version]
- Tran, L.S.P.; Urao, T.; Qin, F.; Maruyama, K.; Kakimoto, T.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid, drought, and salt stress in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 20623–20628. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xiong, Y.; Bassham, D.C. Autophagy is required for tolerance of drought and salt stress in plants. Autophagy 2009, 5, 954–963. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Wang, P.; Jia, X.; Huo, L.; Che, R.; Ma, F. Improvement of drought tolerance by overexpressing MdATG18a is mediated by modified antioxidant system and activated autophagy in transgenic apple. Plant Biotechnol. J. 2018, 16, 545–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, B.; Xie, L.; Yi, T.; Lv, J.; Yang, H.; Cheng, X.; Liu, F.; Zou, X. Genome-Wide Identification and Characterization of the Mitochondrial Transcription Termination Factors (mTERFs) in Capsicum annuum L. Int. J. Mol. Sci. 2019, 21, 269. [Google Scholar] [CrossRef] [Green Version]
- Robles, P.; Micol, J.L.; Quesada, V. Arabidopsis MDA1, a nuclear-encoded protein, functions in chloroplast development and abiotic stress responses. PLoS ONE 2012, 7, e42924. [Google Scholar] [CrossRef] [PubMed]
- Afzal, Z.; Howton, T.C.; Sun, Y.; Mukhtar, M.S. The roles of aquaporins in plant stress responses. J. Dev. Biol. 2016, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.Y.; Kim, D.G.; Kim, Y.O.; Kim, J.S.; Kang, H. An Expression Analysis of a Gene Family Encoding Plasma Membrane Aquaporins in Response to Abiotic Stresses in Arabidopsis Thaliana. Plant Mol. Biol. 2004, 54, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Alexandersson, E.; Danielson, J.Å.H.; Råde, J.; Moparthi, V.K.; Fontes, M.; Kjellbom, P.; Johanson, U. Transcriptional regulation of aquaporins in accessions of Arabidopsis in response to drought stress. Plant J. 2010, 61, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Wang, Z.Y.; Lin, H.; Cui, W.E.; Chen, J.; Liu, M.; Chen, Z.L.; Qu, L.J.; Gu, H. Expression and functional analysis of the rice plasma-membrane intrinsic protein gene family. Cell Res. 2006, 16, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Dixon, R.A. Heterodimeric geranyl(geranyl)diphosphate synthase from hop (Humulus lupulus) and the evolution of monoterpene biosynthesis. Proc. Natl. Acad. Sci. USA 2009, 106, 9914–9919. [Google Scholar] [CrossRef] [Green Version]
- Radwan, A.; Kleinwächter, M.; Selmar, D. Impact of drought stress on specialised metabolism: Biosynthesis and the expression of monoterpene synthases in sage (Salvia officinalis). Phytochemistry 2017, 141, 20–26. [Google Scholar] [CrossRef]
- Savoi, S.; Wong, D.C.J.; Arapitsas, P.; Miculan, M.; Bucchetti, B.; Peterlunger, E.; Fait, A.; Mattivi, F.; Castellarin, S.D. Transcriptome and metabolite profiling reveals that prolonged drought modulates the phenylpropanoid and terpenoid pathway in white grapes (Vitis vinifera L.). BMC Plant Biol. 2016, 16, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriegs, B.; Jansen, M.; Hahn, K.; Peisker, H.; Šamajová, O.; Beck, M.; Braun, S.; Ulbrich, A.; Baluška, F.; Schulz, M. Cyclic monoterpene mediated modulations of Arabidopsis thaliana phenotype. Plant Signal. Behav. 2010, 5, 832–838. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.; Magwanga, R.O.; Kirungu, J.N.; Hu, Y.; Dong, Q.; Cai, X.; Zhou, Z.; Wang, X.; Zhang, Z.; Hou, Y.; et al. Overexpression of Cotton a DTX/MATE Gene Enhances Drought, Salt, and Cold Stress Tolerance in Transgenic Arabidopsis. Front. Plant Sci. 2019, 10, 299. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhu, H.; Pan, Y.; Yu, Y.; Luan, S.; Li, L. A DTX/MATE-Type Transporter Facilitates Abscisic Acid Efflux and Modulates ABA Sensitivity and Drought Tolerance in Arabidopsis. Mol. Plant 2014, 7, 1522–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marathe, A.; Krishnan, V.; Vinutha, T.; Dahuja, A.; Jolly, M.; Sachdev, A. Exploring the role of Inositol 1,3,4-trisphosphate 5/6 kinase-2 (GmITPK2) as a dehydration and salinity stress regulator in Glycine max (L.) Merr. through heterologous expression in E. coli. Plant Physiol. Biochem. PPB 2018, 123, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Liu, L.; You, L.; Yang, M.; He, Y.; Li, X.; Xiong, L. Characterization of an inositol 1,3,4-trisphosphate 5/6-kinase gene that is essential for drought and salt stress responses in rice. Plant Mol. Biol. 2011, 77, 547–563. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Xu, K.; Chen, S.; Li, T.; Xia, H.; Chen, L.; Liu, H.; Luo, L. A stress-responsive bZIP transcription factor OsbZIP62 improves drought and oxidative tolerance in rice. BMC Plant Biol. 2019, 19, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Wahid, A.; Kobayashi, N.S.M.A.; Fujita, D.B. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Temnykh, S.; DeClerck, G.; Lukashova, A.; Lipovich, L.; Cartinhour, S.; McCouch, S. Computational and Experimental Analysis of Microsatellites in Rice (Oryza sativa L.): Frequency, Length Variation, Transposon Associations, and Genetic Marker Potential. Genome Res. 2001, 11, 1441–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darvasi, A.; Soller, M. A Simple Method to Calculate Resolving Power and Confidence Interval of QTL Map Location. Behav. Genet. 1997, 27, 125–132. [Google Scholar] [CrossRef]
- Chardon, F.; Jasinski, S.; Durandet, M.; Lécureuil, A.; Soulay, F.; Bedu, M.; Guerche, P.; Masclaux-Daubresse, C. QTL meta-analysis in Arabidopsis reveals an interaction between leaf senescence and resource allocation to seeds. J. Exp. Bot. 2014, 65, 3949–3962. [Google Scholar] [CrossRef] [PubMed] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Title | Parents | Type of Crossing | Number of Marker Used | Type of Marker | Population Size | Number of QTLs | References |
|---|---|---|---|---|---|---|---|
| Mapping QTLs for root traits in a recombinant inbred population from two indica ecotypes in rice | IR58821–23-B-1–2-1 X IR52561-UBN-1–1-2 | RIL | 367 | AFLP, RFLP | 166 | 39 | [28] |
| A large-effect QTL for grain yield under reproductive-stage drought stress in upland rice | Vandana X Way Rarem | SF3 | 126 | SSR | 436 | 22 | [29] |
| Locating QTLs controlling constitutive root traits in the rice population IAC 165 × Co39. | IAC 165 X CO39 | RIL | 182 | RFLP, SSR | 125 | 42 | [30] |
| Molecular mapping and location of QTLs for drought-resistance traits in indica rice (Oryza sativa L.) lines adapted to target environments | Nootripathu X IR20 | RIL | 79 | SSR | 250 | 21 | [31] |
| Using chromosome introgression lines to map Quantitative Trait Loci for photosynthesis parameters in rice (Oryza sativa L.) leaves under drought and well-watered field conditions. | Haogelao X Shennong265 | BC | 130 | SSR | 94 | 29 | [32] |
| Identification of QTLs controlling rice drought tolerance at seedling stage in hydroponic culture. | IRAT109 X Akihikari | BC | 57 | SSR | 106 | 5 | [33] |
| Genetic analysis of rainfed lowland rice drought tolerance under naturally-occurring stress in eastern India: Heritability and QTL effects. | CT9993 X IR62266 | DH | 315 | AFLP, RFLP, SSR | 154 | 18 | [34] |
| Quantitative Trait Loci Associated with Drought Tolerance at Reproductive Stage in Rice. | CT9993 X IR62266 | DH | 332 | AFLP, RFLP, SSR | 154 | 69 | [19] |
| Saturation mapping of QTL regions and identification of putative candidate genes for drought tolerance in rice. | CT9993 X IR62266 | DH | 400 | AFLP, RFLP, SSR | 154 | 42 | [35] |
| Grains yield QTLs with consistent-effect under reproductive- stage drought stress in rice. | KaliAus X IR64 KaliAus X MTU1010 | BC | 4 | SSR | 300 | 3 | [36] |
| Mapping consistent rice (Oryza sativa L.) yield QTLs under drought stress in target rainfed environments. | Nootripathu X IR20 | RIL | 79 | SSR | 397 | 2 | [37] |
| Identification and mapping of QTLs associated with drought tolerance traits in rice by a cross between Super Basmati and IR55419-04. | IR55419-04 X Super Basmati | SF2 | 73 | SSR | 418 | 21 | [38] |
| Genetic variation, linkage mapping of QTL and correlation studies for yield, root, and agronomic traits for aerobic adaptation. | MASARB25 X Pusa Basmati 1460 MAS26 X HKR47 | SF2 | 50 | SSR | 250 | 35 | [39] |
| Genetic mapping of quantitative trait loci for grain yield under drought in rice under controlled greenhouse conditions. | Vandana X Cocodrie | SF2 | 88 | InDels. SNP, SSR | 187 | 6 | [18] |
| Identification of Quantitative Trait Loci for ABA sensitivity at seed germination and seedling stages in rice. | IRAT109 X Zhenshan 97 | RIL | 220 | SSR | 154 | 17 | [40] |
| Identification of quantitative trait loci for four morphologic traits under water stress in rice (Oryza sativa L.). | IRAT109 X Zhenshan 97 | RIL | 220 | SSR | 180 | 36 | [41] |
| Genetic Basis of Drought Resistance at Reproductive Stage in Rice: Separation of Drought Tolerance From Drought Avoidance. | IRAT109 X Zhenshan 97 | RIL | 220 | SSR | 180 | 12 | [42] |
| Genetic analysis for drought resistance of rice at reproductive stage in field with different types of soil. | IRAT109 X Zhenshan 97 | RIL | 220 | SSR | 180 | 53 | [43] |
| Locating genomic regions associated with components of drought resistance in rice: comparative mapping within and across species. | CT9993 X IR62266 | DH | 315 | AFLP, RFLP, SSR | 154 | 41 | [44] |
| Comparison of QTLs for rice seedling morphology under different water supply conditions. | Azucena X IR64 | DH | 189 | RFLP, SSR | 96 | 12 | [45] |
| Meta-QTL | Position (cM) | Flanking Markers | Distance (cM) | Number of QTL | Number of Traits | Traits Involved |
|---|---|---|---|---|---|---|
| MQTL 1.1 | 34.57 | RM522–RG394 | 6.84 | 3 | 3 | FL, Wat, Lf |
| MQTL 1.2 | 98.65 | RM5–RM34B | 3.2 | 5 | 4 | Lf, Bio, Temp, Rt |
| MQTL 1.3 | 126.31 | RM1216–RM1198 | 2.82 | 12 | 6 | Rt, Lf, GY, DS, FL, Sd |
| MQTL 1.4 | 147.97 | RM543–RM3825 | 2.4 | 8 | 4 | Rt, FL, Bio, PH |
| MQTL 1.5 | 163.7 | RG109–RM315 | 3.17 | 17 | 6 | GY, PH, Bio, SH, Rt, FL |
| MQTL 1.6 | 182.77 | RM431–RM1067 | * | 17 | 8 | GY, Bio, Lf, PH, SH, St, Rt, FL |
| MQTL 2.1 | 107.52 | G45–RM6379 | 3.39 | 21 | 9 | St, FL, Lf, Bio, Rt, DS, Temp, Sd, GY |
| MQTL 2.2 | 132.4 | RM5470–RG139 | * | 32 | 10 | Rt, Lf, FL, RS, GY, St, PH, Temp, Bio, Wat |
| MQTL 3.1 | 24.41 | RZ329–C63 | 3.58 | 25 | 8 | Rt, Lf, St, Bio, FL, Wat, GY, PH |
| MQTL 3.2 | 59.9 | RG396–RG227 | * | 11 | 4 | Lf, Rt, FL, GY |
| MQTL 4.1 | 29.02 | PC33M4–RM5953 | 4.47 | 8 | 3 | Bio, Rt, Lf |
| MQTL 4.2 | 49.97 | PC11M12–RM185 | 1.14 | 6 | 2 | Bio, Rt |
| MQTL 4.3 | 54.59 | PC38M7–ME2_13 | 1.14 | 6 | 2 | Rt, FL |
| MQTL 4.4 | 60.77 | PC36M3–ME10_11 | 4.76 | 4 | 3 | GY, Rt, Lf |
| MQTL 4.5 | 88.6 | PC27M13–EMP3_10 | 1.56 | 1 | 1 | PH |
| MQTL 4.6 | 95.9 | ME9_4–C335 | 4.44 | 3 | 2 | FL, Rt |
| MQTL 4.7 | 116.17 | RM470–RM303 | 1.52 | 6 | 2 | Lf, Rt |
| MQTL 4.8 | 124.22 | RG939–RM348 | 1.65 | 3 | 2 | FL, Rt |
| MQTL 4.9 | 132.98 | R2017–R0874 | 1.87 | 7 | 3 | FL, Rt, Bio |
| MQTL 4.10 | 142.86 | G235–RG143 | * | 6 | 3 | Bio, Rt, Lf |
| MQTL 5.1 | 13.05 | RZ536D–C119 | 9.03 | 4 | 4 | GY, FL, Rt, Bio |
| MQTL 5.2 | 79.08 | RM430–R2217 | 5.43 | 8 | 5 | PH, Bio, Lf, Temp, Rt |
| MQTL 5.3 | 110.1 | RM233B–RM421 | 1.74 | 6 | 5 | FL, St, Rt, Bio, Wat |
| MQTL 5.4 | 119.36 | RM26–RG119 | * | 6 | 4 | Sd, Lf, FL, Bio |
| MQTL 6.1 | 3.4 | RM540–RM190 | 5.87 | 1 | 1 | Bio |
| MQTL 6.2 | 25.86 | Amp5–RM217 | 1.78 | 7 | 2 | Rt, Bio |
| MQTL 6.3 | 33.84 | L169B–RM111 | 2 | 6 | 4 | GY, FL, Lf, Bio |
| MQTL 6.4 | 45.36 | RM557–RG64 | 1.42 | 1 | 1 | GY |
| MQTL 6.5 | 55.71 | RM136–CD096 | 1.37 | 4 | 4 | FL, Lf, GY, Bio |
| MQTL 6.6 | 64.21 | R111–EM17_8 | 6.94 | 4 | 2 | PH, Bio |
| MQTL 6.7 | 105.22 | RM454–RM275 | 5.95 | 9 | 5 | Lf, Rt, Bio, FL, GY |
| MQTL 6.8 | 142.5 | RM439–RM141 | * | 5 | 5 | Bio, Wat, temp, SH, DS |
| MQTL 7.1 | −16.55 | T12A1–ME2_15 | 9.7 | 5 | 4 | Lf, Rt, Bio, PH |
| MQTL 7.2 | 22.33 | RZ448–RM125 | 1.66 | 2 | 2 | Rt, Bio |
| MQTL 7.3 | 29.67 | ME10_6–RM8010 | 2.67 | 16 | 6 | FL, Lf, PH, GY, Rt, Bio |
| MQTL 7.4 | 44.34 | ME6_8–RM2878 | 7.23 | 1 | 1 | Rt |
| MQTL 7.5 | 79.42 | PC34M16–RM351 | 5.31 | 2 | 2 | Bio, Rt |
| MQTL 7.6 | 105.9 | RG404–RM248 | 0.66 | 6 | 5 | Bio, FL, 2 Rt, Lf, St |
| MQTL 8.1 | 13.8 | RM152–RZ143 | 4.43 | 8 | 4 | Lf, FL, St, Bio |
| MQTL 8.2 | 57.89 | RM126–ME5_3 | 0.59 | 5 | 4 | Lf, Rt, St, FL |
| MQTL 8.3 | 59.77 | MRG2181–RM137 | 3.92 | 4 | 3 | FL, GY, Rt |
| MQTL 8.4 | 75.37 | RM6032–G2132 | 2.74 | 6 | 6 | Temp, Wat, DS, GY, Rt, FL |
| MQTL 8.5 | 86.89 | G187–ME2_11 | 2.81 | 7 | 3 | FL, GY, Bio |
| MQTL 8.6 | 99.96 | RZ997–RZ572 | 2.93 | 11 | 3 | PH, FL, Rt |
| MQTL 8.7 | 112.54 | RM1615–RZ66 | 3.16 | 2 | 1 | FL |
| MQTL 8.8 | 122.49 | RM477–Rcn7 | * | 6 | 5 | FL, Lf, St, Bio, Sd |
| MQTL 9.1 | −0.61 | PC75M6–PC75M4 | 0.91 | 2 | 1 | FL |
| MQTL 9.2 | 1.71 | ME2_17–C711 | 1.47 | 6 | 2 | FL, Rt |
| MQTL 9.3 | 5.35 | PC32M13–S55 | 1.17 | 1 | 1 | Bio |
| MQTL 9.4 | 9.03 | S55–PC12M5 | 2.39 | 8 | 5 | FL, GY, Lf, Bio, PH |
| MQTL 9.5 | 20.2 | EM14_6–RG553 | 1.41 | 5 | 4 | Rt, Lf, Bio, Wat |
| MQTL 9.6 | 28.23 | RM296–RM105 | 3.66 | 1 | 1 | Lf |
| MQTL 9.7 | 64.11 | ME9_3–RM257 | 2.87 | 18 | 5 | PH, FL, Bio, Rt, Lf |
| MQTL 9.8 | 93.31 | RZ596–HSP84B | * | 8 | 5 | Bio, GY, DS, Rt, FL |
| MQTL 10.1 | −39.67 | G333–EM18_17 | 10.36 | 3 | 3 | Bio, GY, FL |
| MQTL 10.2 | 14.52 | RM222–RM216 | 9.56 | 1 | 1 | FL |
| MQTL 10.3 | 49.1 | RM467–RM1937 | 2.21 | 5 | 4 | PH, FL, Bio, Lf |
| MQTL 10.4 | 57.73 | RM596–RM271 | 16.02 | 5 | 4 | Bio, DS, Rt, St |
| MQTL 10.5 | 107.67 | RM34–RM333 | * | 6 | 4 | FL, GY, Lf, PH |
| MQTL 11.1 | 38.41 | RM6894–RM552 | 1.15 | 11 | 4 | Bio, Rt, Lf, FL |
| MQTL 11.2 | 60.79 | PC4M2–PC33M2 | 1.62 | 4 | 3 | Bio, Rt, St |
| MQTL 11.3 | 84.53 | PC35M2–RM21 | 0.87 | 6 | 3 | Lf, Bio, Rt |
| MQTL 11.4 | 93.64 | Xa23–ME7_2 | 1.14 | 1 | 1 | Rt |
| MQTL 11.5 | 106.12 | RM206–RM254 | 1.14 | 3 | 3 | PH, Temp, FL |
| MQTL 11.6 | 118.17 | RM5926–C950 | * | 2 | 2 | FL, Rt |
| MQTL 12.1 | 24.55 | RM19–RM453 | 9.2 | 1 | 1 | FL |
| MQTL 12.2 | 55.15 | AF6 –RM28130 | 1.61 | 1 | 1 | GY |
| MQTL 12.3 | 62.19 | RM260–RM519 | 4.55 | 4 | 2 | Bio, Rt |
| MQTL 12.4 | 87.5 | CD0344–RG901 | 2.48 | 3 | 2 | Rt, Bio |
| MQTL 12.5 | 105.4 | RG958–RG323 | * | 5 | 5 | Bio, PH, Lf, FL, Rt |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selamat, N.; Nadarajah, K.K. Meta-Analysis of Quantitative Traits Loci (QTL) Identified in Drought Response in Rice (Oryza sativa L.). Plants 2021, 10, 716. https://doi.org/10.3390/plants10040716
Selamat N, Nadarajah KK. Meta-Analysis of Quantitative Traits Loci (QTL) Identified in Drought Response in Rice (Oryza sativa L.). Plants. 2021; 10(4):716. https://doi.org/10.3390/plants10040716
Chicago/Turabian StyleSelamat, Nurhanis, and Kalaivani K. Nadarajah. 2021. "Meta-Analysis of Quantitative Traits Loci (QTL) Identified in Drought Response in Rice (Oryza sativa L.)" Plants 10, no. 4: 716. https://doi.org/10.3390/plants10040716
APA StyleSelamat, N., & Nadarajah, K. K. (2021). Meta-Analysis of Quantitative Traits Loci (QTL) Identified in Drought Response in Rice (Oryza sativa L.). Plants, 10(4), 716. https://doi.org/10.3390/plants10040716