
From Genetic Maps to QTL Cloning: An Overview for Durum Wheat




Abstract
:1. Introduction
2. Genotyping Tools
3. Linkage and Consensus Maps
4. Traits and QTL Analysis
4.1. Quality
4.2. Biotic Stress
4.3. Abiotic Stress
4.4. QTL Mapping
4.5. Genome-Wide Association Studies
4.6. QTL Meta-Analysis
5. Innovative Experimental Designs for Enhanced Gene Discovery
5.1. Bulked Segregant Analysis by Sequencing (BSAseq)
5.2. Development of New Populations
5.3. Candidate Gene: A New Approach for Studying Quantitative Trait Loci
6. Gene Editing in Durum Wheat
7. From Classical Breeding to Genomic Selection
8. Speed Breeding
9. Future Prospects
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 78–202. [Google Scholar] [CrossRef]
- Royo, C.; Álvaro, F.; Martos, V.; Ramdani, A.; Isidro, J.; Villegas, D.; García del Moral, L.F. Genetic changes in durum wheat yield components and associated traits in Italian and Spanish varieties during the 20th century. Euphytica 2007, 155, 259–270. [Google Scholar] [CrossRef]
- Subira, J.; Peña, R.J.; Álvaro, F.; Ammar, K.; Ramdani, A.; Royo, C. Breeding progress in the pasta-making quality of durum wheat cultivars released in Italy and Spain during the 20th Century. Crop Pasture Sci. 2014, 65, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Maccaferri, M.; Cané, M.A.; Sanguineti, M.C.; Salvi, S.; Colalongo, M.C.; Massi, A.; Clarke, F.; Knox, R.; Pozniak, C.J.; Clarke, J.M.; et al. A consensus framework map of durum wheat (Triticum durum Desf.) suitable for linkage disequilibrium analysis and genome-wide association mapping. BMC Genom. 2014, 15, 873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colasuonno, P.; Gadaleta, A.; Giancaspro, A.; Nigro, D.; Give, S.; Incerti, O.; Mangini, G.; Signorile, A.; Simeone, R.; Blanco, A. Development of a high-density SNP-based linkage map and detection of yellow pigment content QTLs in durum wheat. Mol. Breed. 2014, 34, 1563–1578. [Google Scholar] [CrossRef]
- Kumar, S.; Goyal, A.; Mohan, A.; Balyan, H.S.; Gupta, P.K. An integrated physical map of simple sequence repeats in bread wheat. Aust. J. Crop Sci. 2013, 7, 460–468. [Google Scholar]
- Blanco, A.; Mangini, G.; Giancaspro, A.; Giove, S.; Colasuonno, P.; Simeone, R.; Signorile, A.; de Vita, P.; Mastrangelo, A.M.; Cattivelli, L.; et al. Relationships between grain protein content and grain yield components through quantitative trait locus analyses in a recombinant inbred line population derived from two elite durum wheat cultivars. Mol. Breed. 2012, 30, 79–92. [Google Scholar] [CrossRef]
- Maccaferri, M.; Mantovani, P.; Tuberosa, R.; DeAmbrogio, E.; Giuliani, S.; Demontis, A.; Massi, A.; Sanguineti, M.C. A major QTL for durable leaf rust resistance widely exploited in durum wheat breeding programs maps on the distal region of chromosome arm 7BL. Theor. Appl. Genet. 2008, 117, 1225–1240. [Google Scholar] [CrossRef]
- Blanco, A.; Gadaleta, A.; Cenci, A.; Carluccio, A.V.; Abdelbacki, A.M.M.; Simeone, R. Molecular mapping of the novel powdery mildew resistance gene Pm36 introgressed from Triticum turgidum var dicoccoides in durum wheat. Theor. Appl. Genet. 2008, 117, 135–142. [Google Scholar] [CrossRef]
- Blanco, A.; Colasuonno, P.; Gadaleta, A.; Mangini, G.; Schiavulli, A.; Simeone, R.; Digesù, A.M.; de Vita, P.; Mastrangelo, A.M.; Cattivelli, L. Quantitative trait loci for yellow pigment concentration and individual carotenoid compounds in durum wheat. J. Cereal Sci. 2011, 54, 255–264. [Google Scholar] [CrossRef]
- Gadaleta, A.; Giancaspro, A.; Give, S.L.; Zacheo, S.; Mangini, G.; Simeone, R.; Signorile, A.; Blanco, A. Genetic and physical mapping of new EST-derived SSRs on the A and B genome chromosomes of wheat. Theor. Appl. Genet. 2009, 118, 1015–1025. [Google Scholar] [CrossRef]
- Maccaferri, M.; Harris, N.S.; Twardziok, S.O.; Pasam, R.K.; Gundlach, H.; Spannagl, M.; Ormanbekova, D.; Lux, T.; Prade, V.M.; Milner, S.G.; et al. Durum wheat genome reveals past domestication signatures and future improvement targets. Nat. Genet. 2019, 51, 885–895. [Google Scholar] [CrossRef] [Green Version]
- Nam, J.H.; Song, H.S.; Suh, S.J.; Heo, H.Y.; Park, M.W.; Baek, S.B.; Park, H.H. Achievement and prospect of good quality breeding in winter cereals. Korean J. Crop Sci. 1998, 43, 11–15. [Google Scholar]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- Mullis, K.B.; Faloona, F. Specific synthesis of DNA in vitro via a polymerase catalysed chain reaction. Meth. Enzymol. 1987, 155, 335–350. [Google Scholar]
- Williams, J.G.K.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphism amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef] [Green Version]
- Tautz, D.; Trick, M.; Dover, G. Cryptic simplicity in DNA is a major source of genetic variation. Nature 1986, 322, 652–656. [Google Scholar] [CrossRef]
- Tautz, D. Hypervariability of simple sequence as a general source for polymorphic DNA markers. Nucleic Acids Res. 1989, 17, 6463–6471. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [Green Version]
- Jaccoud, D.; Peng, K.; Feinstein, D.; Kilian, A. Diversity arrays: A solid state technology for sequence information independent genotyping. Nucleic Acid Res. 2001, 29, E25. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Wang, Y.; Chen, M.; Edae, E.; Poland, J.; Akhunov, E.; Chao, S.; Bai, G.; Carver, B.F.; Yan, L. Precisely mapping a major gene conferring resistance to Hessian fly in bread wheat using genotyping-by-sequencing. BMC Genom. 2015, 16, 108. [Google Scholar] [CrossRef] [Green Version]
- Cabral, A.L.; Jordan, M.C.; McCartney, C.A.; You, F.M.; Humphreys, D.G.; MacLachlan, R.; Pozniac, C.J. Identification of candidate genes.; regions and markers for pre-harvest sprouting resistance in wheat (Triticum aestivum L.). BMC Plant Biol. 2014, 14, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Priya, P.; Misra, G.; Yadav, G. Structural and biochemical perspectives in plant isoprenoid biosynthesis. Phytochem. Rev. 2013, 12, 255–291. [Google Scholar] [CrossRef]
- Cavanagh, C.R.; Chao, S.; Wang, S.; Huang, B.E.; Stephen, S.; Kiani, S.; Forrest, K.; Saintenac, C.; Brown-Guedira, G.L.; Akhunova, A. Genome-wide comparative diversity uncovers multiple targets of selection for improvement in hexaploid wheat landraces and cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 8057–8062. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using the high-density 90,000 SNP array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Boeven, P.H.; Longin, C.F.; Leiser, W.L.; Kollers, S.; Ebmeyer, E.; Würschum, T. Genetic architecture of male floral traits required for hybrid wheat breeding. Theor. Appl. Genet. 2016, 129, 2343–2357. [Google Scholar] [CrossRef]
- Winfield, M.O.; Allen, A.M.; Burridge, A.J.; Barker, G.L.A.; Benbow, H.R.; Wilkinson, P.A.; Coghill, J.; Waterfall, C.; Davassi, A.; Scopes, G. High-density SNP genotyping array for hexaploid wheat and its secondary and tertiary gene pool. Plant Biotechnol. J. 2016, 14, 1195–1206. [Google Scholar] [CrossRef]
- Allen, A.M.; Winfield, M.O.; Burridge, A.J.; Downie, R.C.; Benbow, H.R.; Barker, G.L.; Wilkinson, P.A.; Coghill, J.; Waterfall, C.; Davassi, A. Characterization of a Wheat Breeders Array suitable for high-throughput SNP genotyping of global accessions of hexaploid bread wheat (Triticum aestivum). Plant Biotechnol. J. 2017, 15, 390–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasheed, A.; Xia, X. From markers to genome-based breeding in wheat. Theor. Appl. Genet. 2019, 132, 767–784. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Dong, Z.; Zhao, L.; Ren, Y.; Zhang, N.; Chen, F. The Wheat 660K SNP array demonstrates great potential for marker-assisted selection in polyploid wheat. Plant Biotechnol. J. 2020, 18, 1354–1360. [Google Scholar] [CrossRef] [PubMed]
- Maccaferri, M.; Ricci, A.; Salvi, S.; Milner, S.G.; Noli, E.; Martelli, P.L.; Casadio, R.; Akhunov, E.; Scalabrin, S.; Vendramin, V.; et al. A high-density SNP-based consensus map of tetraploid wheat as a bridge to integrate durum and bread wheat genomics and breeding. Plant Biotechnol. J. 2015, 13, 5. [Google Scholar] [CrossRef]
- Pont, C.; Murat, F.; Guizard, S.; Flores, R.; Foucrier, S.; Bidet, Y.; Quraishi, U.M.; Alaux, M.; Doležel, J.; Fahima, T.; et al. Wheat syntenome unveils new evidences of contrasted evolutionary plasticity between paleo- and neoduplicated subgenomes. Plant J. 2013, 76, 1030–1044. [Google Scholar] [CrossRef] [Green Version]
- Allen, A.M.; Barker, G.L.A.; Berry, S.T.; Coghill, J.A.; Gwilliam, R.; Kirby, S.; Robinson, P.; Brenchley, R.C.; D’Amore, R.; McKenzie, N.; et al. Transcript-specific.; single-nucleotide polymorphism discovery and linkage analysis in hexaploid bread wheat (Triticum aestivum L.). Plant Biotechnol. J. 2011, 9, 1086–1099. [Google Scholar] [CrossRef] [Green Version]
- Giancaspro, A.; Give, S.L.; Zito, D.; Blanco, A.; Gadaleta, A. Mapping QTL for Fusarium head blight resistance in an interspecific wheat population. Front. Plant Sci. 2016, 7, 1381. [Google Scholar] [CrossRef]
- Marcotuli, I.; Houston, K.; Schwerdt, J.G.; Waugh, R.; Fincher, G.B.; Burton, R.A.; Blanco, A.; Gadaleta, A. Genetic diversity and genome wide association study of β-glucan content in tetraploid wheat grains. PLoS ONE 2016, 11, e0152590. [Google Scholar] [CrossRef] [Green Version]
- Jaganathan, D.; Bohra, A.; Thudi, M.; Varshney, R.K. Fine mapping and gene cloning in the post-NGS era: Advances and prospects. Theor. Appl. Genet. 2020, 133, 1791–1810. [Google Scholar] [CrossRef] [Green Version]
- Sajjad, M.; Khan, S.H.; Shahzad, M. Patterns of allelic diversity in spring wheat populations by SSR-markers. Cytol. Genet. 2018, 52, 155–160. [Google Scholar] [CrossRef]
- Gill, H.S.; Li, C.; Sidhu, J.S.; Liu, W.; Wilson, D.; Bai, G.; Gill, B.S.; Sehgal, S.K. Fine Mapping of the Wheat Leaf Rust Resistance Gene Lr42. Int. J. Mol. Sci. 2019, 20, 2445. [Google Scholar] [CrossRef] [Green Version]
- Blanco, A.; De Giovanni, C.; Bellomo, M.P.; Cenci, A.; D’Ovidio, R.; Iacono, E.; Laddomada, B.; Pagnotta, M.A.; Porcedu, E.; Sciancalepore, A.; et al. A genetic linkage map of durum wheat. Theor. Appl. Genet. 1998, 97, 721–728. [Google Scholar] [CrossRef]
- Wenzl, P.; Carling, J.; Kudrna, D.; Jaccoud, D.; Huttner, E.; Kleinhofs, A.; Kilian, A. Diversity Arrays Technology (DArT) for whole-genome profiling of barley. Proc. Natl. Acad. Sci. USA 2004, 101, 9915–9920. [Google Scholar] [CrossRef] [Green Version]
- Marone, D.; Laidò, G.; Gadaleta, A.; Colasuonno, P.; Ficco, D.B.; Giancaspro, A.; Giove, S.; Panio, G.; Russo, M.A.; De Vita, P.; et al. A high-density consensus map of A and B wheat genomes. Theor. Appl. Genet. 2012, 125, 1619–1638. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Han, Y.; Zhao, Y.F.; Troggio, M.; Korban, S.S. A multi-population consensus genetic map reveals inconsistent marker order among maps likely attributed to structural variations in the apple genome. PLoS ONE 2012, 7, e47864. [Google Scholar] [CrossRef] [Green Version]
- Korzun, V.; Röder, M.S.; Wendekake, K.; Pasqualone, A.; Lotti, C.; Ganal, M.W.; Blanco, A. Integration of dinucleotide microsatellites from hexaploid bread wheat into a genetic linkage map of durum wheat. Theor. Appl. Genet. 1999, 98, 1202–1207. [Google Scholar] [CrossRef]
- Lotti, C.; Salvi, S.; Pasqualone, A.; Tuberosa, R.; Blanco, A. Integration of AFLP markers into an RFLP-based map of durum wheat. Plant Breed. 2000, 119, 393–401. [Google Scholar] [CrossRef]
- Nachit, M.; Elouafi, I.; Pagnotta, M.A.; El Saleh, A.; Iacono, E.; Labhilili, M.; Asbati, A.; Azrak, M.; Hazzam, H.; Benscher, D.; et al. Molecular linkage map for an intraspecific recombinant inbred population of durum wheat (Triticum turgidum L. var durum). Theor. Appl. Genet. 2001, 102, 177–186. [Google Scholar] [CrossRef]
- Blanco, A.; Simeone, R.; Cenci, A.; Gadaleta, A.; Tanzarella, O.A.; Porceddu, E.; Salvi, S.; Tuberosa, R.; Figliuolo, G.; Spagnoletti, P.; et al. Extension of the Messapia X dicoccoides linkage map of Triticum turgidum (L.) Thell. Cell. Mol. Biol. Lett. 2004, 9, 529–541. [Google Scholar]
- Elouafi, I.; Nachit, M.M. A genetic linkage map of the Durum x Triticum dicoccoides backcross population based on SSRs and AFLP markers.; and QTL analysis for milling traits. Theor. Appl. Genet. 2004, 108, 401–413. [Google Scholar] [CrossRef]
- Somers, D.J.; Fedak, G.; Clarke, J.; Cao, W. Mapping of FHB resistance QTLs in tetraploid wheat. Genome 2006, 49, 1586–1593. [Google Scholar] [CrossRef]
- Pozniak, C.J.; Knox, R.E.; Clarke, F.R.; Clarke, J.M. Identification of QTL and association of a phytoene synthase gene with endosperm colour in durum wheat. Theor. Appl. Genet. 2007, 114, 525–537. [Google Scholar] [CrossRef]
- Mantovani, P.; Maccaferri, M.; Sanguineti, M.C.; Tuberosa, R.; Catizone, I.; Wenzl, P.; Thompson, B.; Carling, J.; Huttner, E.; DeAmbrogio, E.; et al. An integrated DArT-SSR linkage map of durum wheat. Mol. Breed. 2008, 22, 629–648. [Google Scholar] [CrossRef]
- Peleg, Z.; Saranga, Y.; Suprunova, T.; Ronin, Y.; Röder, M.; Kilian, A.; Korol, A.B.; Fahima, T. High-density genetic map of durum wheat × wild emmer wheat based on SSR and DArT markers. Theor. Appl. Genet. 2008, 117, 103–115. [Google Scholar] [CrossRef]
- Zhang, W.; Dubcovsky, J. Association between allelic variation at the Phytoene synthase 1 gene and yellow pigment content in the wheat grain. Theor. Appl. Genet. 2008, 116, 635–645. [Google Scholar] [CrossRef] [Green Version]
- Marone, D.; Del Olmo, A.I.; Laido, G.; Sillero, J.C.; Emeran, A.A.; Russo, M.A.; Ferragonio, P.; Giovaniello, V.; Mazzucotelli, E.; De Leonardis, A.M. Genetic analysis of durable resistance against leaf rust in durum wheat. Mol. Breed. 2009, 24, 25–39. [Google Scholar] [CrossRef]
- Suprayogi, Y.; Pozniak, C.J.; Clarke, F.R.; Clarke, J.M.; Knox, R.E.; Singh, A.K. Identification and validation of quantitative trait loci for grain protein concentration in adapted Canadian durum wheat populations. Theor. Appl. Genet. 2009, 119, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.A.; Ficco, D.B.M.; Marone, D.; De Vita, P.; Vallega, V.; Rubies-Autonell, C.; Rati, C.; Ferragonio, P.; Giovaniello, V.; Pecchioni, N. A major QTL for resistance to soil-borne cereal mosaic virus derived from an old Italian durum wheat cultivar. J. Plant Interact. 2012, 7, 290–300. [Google Scholar] [CrossRef]
- Buerstmayr, M.; Huber, K.; Heckmann, J.; Steiner, B.; Nelson, J.; Buerstmayr, H. Mapping of QTL for Fusarium head blight resistance and morphological and developmental traits in three backcross populations derived from Triticum dicoccum × Triticum durum. Theor. Appl. Genet. 2012, 125, 1751–1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, Y.; Comeau, A.; Langevin, F.; Hucl, P.; Clarke, J.; Brule-Babel, A.; Pozniak, C. Identification of novel QTL for resistance to Fusarium head blight in a tetraploid wheat population. Genome 2012, 55, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Patil, R.; Tamhankar, S.; Oak, M.; Raut, A.; Honrao, B.; Rao, V.S.; Misra, S.C. Mapping of QTL for agronomic traits and kernel characters in durum wheat (Triticum durum Desf). Euphytica 2013, 190, 117–129. [Google Scholar] [CrossRef]
- Buerstmayr, M.; Alimari, A.; Steiner, B.; Buerstmayr, H. Genetic mapping of QTL for resistance to Fusarium head blight spread (type 2 resistance) in a Triticum dicoccoides × Triticum durum backcross-derived population. Theor. Appl. Genet. 2013, 126, 2825–2834. [Google Scholar] [CrossRef]
- Bansal, U.K.; Kazi, A.G.; Singh, B.; Hare, R.A.; Bariana, H.S. Mapping of durable stripe rust resistance in a durum wheat cultivar Wollaroi. Mol. Breed. 2013, 33, 51–59. [Google Scholar] [CrossRef]
- Faris, J.D.; Zhang, Q.; Chao, S.; Zhang, Z.; Xu, S.S. Analysis of agronomic and domestication traits in a durum × cultivated emmer wheat population using a high-density single nucleotide polymorphism-based linkage map. Theor. Appl. Genet. 2014, 127, 2333–2348. [Google Scholar] [CrossRef]
- Russo, M.A.; Ficco, D.B.M.; Laidò, G.; Marone, D.; Papa, R.; Blanco, A.; Gadaleta, A.; De Vita, P.; Mastrangelo, A.M. A dense durum wheat × T dicoccum linkage map based on SNP markers for the study of seed morphology. Mol. Breed. 2014, 34, 1579–1597. [Google Scholar] [CrossRef]
- Desiderio, F.; Guerra, D.; Rubiales, D.; Piarulli, L.; Pasquini, M.; Mastrangelo, A.M.; Simeone, R.; Blanco, A.; Cattivelli, L.; Valé, G. Identification and mapping of quantitative trait loci for leaf rust resistance derived from a tetraploid wheat Triticum dicoccum accession. Mol. Breed. 2014, 34, 1659–1675. [Google Scholar] [CrossRef]
- Alsaleh, A.; Baloch, F.S.; Derya, M.; Azrak, M.; Kilian, B.; Nachit, M.M. Genetic linkage map of Anatolian durum wheat derived from a cross of Kunduru- 1149 × Cham1. Plant Mol. Biol. Rep. 2015, 33, 209–220. [Google Scholar] [CrossRef]
- Prat, N.; Guilbert, C.; Prah, U.; Wachter, E.; Steiner, B.; Langin, T.; Robert, O.; Buerstmayr, H. QTL mapping of Fusarium head blight resistance in three related durum wheat populations. Theor. Appl. Genet. 2017, 130, 13–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, C.; Basnet, B.R.; Singh, R.P.; Huerta-Espino, J.; Herrera-Foessel, S.A.; Ren, Y.; Randhawa, M.S. Genetic analysis and mapping of adult plant resistance loci to leaf rust in durum wheat cultivar Bairds. Theor. Appl. Genet. 2017, 130, 609–619. [Google Scholar] [CrossRef]
- Nilsen, K.T.; N’Diaye, A.; MacLachlan, P.R.; Clarke, J.M.; Ruan, Y.; Cuthbert, R.D.; Knox, R.E.; Wiebe, K.; Cory, A.T.; Walkowiak, S.; et al. High density mapping and haplotype analysis of the major stem-solidness locus SSt1 in durum and common wheat. PLoS ONE 2017, 12, e0175285. [Google Scholar] [CrossRef]
- Marcotuli, I.; Gadaleta, A.; Mangini, G.; Signorile, A.M.; Zacheo, S.A.; Blanco, A.; Simeone, R.; Colasuonno, P. Development of a high-density SNP-based linkage map and detection of QTL for beta-glucans.; protein content.; grain yield per spike and heading time in durum wheat. Int. J. Mol. Sci. 2017, 18, 1329. [Google Scholar] [CrossRef]
- Giunta, F.; De Vita, P.; Mastrangelo, A.M.; Sanna, G.; Motzo, R. Environmental and genetic variation for yield-related traits of durum wheat as affected by development. Front. Plant Sci. 2018, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Leng, Y.; Chao, S.; Xu, S.S.; Zhong, S. Molecular mapping of QTL for Fusarium head blight resistance introgressed into durum wheat. Theor. Appl. Genet. 2018, 131, 1939–1951. [Google Scholar] [CrossRef]
- Sari, E.; Berraies, S.; Knox, R.E.; Singh, A.K.; Ruan, Y.; Cuthbert, R.D.; Pozniak, C.J.; Henriquez, M.A.; Kumar, S.; Burt, A.J.; et al. High density genetic mapping of Fusarium head blight resistance QTL in tetraploid wheat. PLoS ONE 2018, 13, e0204362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatiukha, A.; Filler, N.; Lupo, I.; Lidzbarsky, G.; Klymiuk, V.; Korol, A.B.; Pozniak, C.; Tzion, F.; Krugmar, T. Grain protein content and thousand kernel weight QTLs identified in a durum × wild emmer wheat mapping population tested in five environments. Theor. Appl. Genet. 2020, 133, 119–131. [Google Scholar] [CrossRef]
- Bassi, F.M.; Brahmi, H.; Sabraoui, A.; Amri, A.; Nsarellah, N.; Nachit, M.M.; Al-Abdallat, A.; Chen, M.S.; Lazraq, A.; El Bouhssini, E. Genetic identification of loci for Hessian fly resistance in durum wheat. Mol. Breed. 2019, 39, 24. [Google Scholar] [CrossRef] [Green Version]
- Pirseyedi, S.M.; Kumar, A.; Ghavami, F.; Hegstad, J.B.; Mergoum, M.; Mazaheri, M.; Elias, E.M. Mapping QTL for Fusarium head blight resistance in a Tunisian-derived durum wheat population. Cereal Res. Commun. 2019, 47, 78–87. [Google Scholar] [CrossRef]
- Desiderio, F.; Zarei, L.; Licciardello, S.; Cheghamirza, K.; Farshadfar, E.; Virzi, N.; Sciacca, F.; Bagnaresi, P.; Battaglia, R.; Guerra, D.; et al. Genomic regions from an Iranian landrace increase kernel size in durum wheat. Front. Plant Sci. 2019, 10, 448. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.S.; Katherine, L.D.; Running, K.L.D.; Xu, S.S.; Zhang, Q.; Peters Haugrud, A.R.; Sharma, S.; McClean, P.E.; Faris, J.D. Genetic analysis of threshability and other spike traits in the evolution of cultivated emmer to fully domesticated durum wheat. Mol. Genet. Genom. 2019, 294, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Aoun, M.; Kolmer, J.A.; Rouse, M.N.; Elias, E.M.; Breiland, M.; Bulbula, W.D.; Chao, S.; Acevedo, M. Mapping of novel leaf rust and stem rust resistance genes in the Portuguese durum wheat landrace PI 192051. G3 2019, 9, 2535–2547. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Salsman, E.; Wang, R.; Galagedara, N.; Zhang, Q.; Fiedler, J.D.; Liu, Z.; Xu, S.; Faris, J.D.; Li, X. Meta-QTL analysis of tan spot resistance in wheat. Theor. Appl. Genet. 2020, 133, 2363–2375. [Google Scholar] [CrossRef]
- Marcotuli, I.; Colasuonno, P.; Hsieh, Y.S.Y.; Fincher, G.B.; Gadaleta, A. Non-Starch Polysaccharides in Durum Wheat: A Review. Int. J. Mol. Sci. 2020, 21, 2933. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.C.; Andreescu, C.; Breseghello, F.; Finney, P.L.; Gualberto, D.G.; Bergman, C.J.; Peña, R.J.; Perretant, M.R.; Leroy, P.; Qualset, C.O.; et al. Quantitative trait locus analysis of wheat quality traits. Euphytica 2006, 149, 145–159. [Google Scholar] [CrossRef]
- Zhang, W.; Chao, S.; Manthey, F.; Chicaiza, O.; Brevis, J.C.; Echenique, V.; Dubcovsky, J. QTL analysis of pasta quality using a composite microsatellite and SNP map of durum wheat. Theor. Appl. Genet. 2008, 117, 1361–1377. [Google Scholar] [CrossRef]
- Mann, G.; Diffey, S.; Cullis, B.; Azanza, F.; Martin, D.; Kelly, A.; McIntyre, L.; Schmidt, A.; Ma, W.; Nath, Z.; et al. Genetic control of wheat quality: Interactions between chromosomal regions determining protein content and composition.; dough rheology.; and sponge and dough baking properties. Theor. Appl. Genet. 2009, 118, 1519–1537. [Google Scholar] [CrossRef]
- Raman, R.; Allen, H.; Raman, H.; Diffey, S.; Martin, P.; McKelvie, K. Localisation of quantitative trait loci for quality attributes in a doubled haploid population of wheat (Triticum aestivum L.). Genome 2009, 52, 701–715. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, J.; Crouch, J.H.; Xu, Y. Efficiency of selective genotyping for genetic analysis of complex traits and potential applications in crop improvement. Mol. Breed. 2010, 26, 493–511. [Google Scholar] [CrossRef]
- Nigro, D.; Gadaleta, A.; Mangini, G.; Colasuonno, P.; Marcotuli, I.; Giancaspro, A.; Giove, S.L.; Simeone, R.; Blanco, A. Candidate genes and genome-wide association study of grain protein content and protein deviation in durum wheat. Planta 2019, 249, 1157–1175. [Google Scholar] [CrossRef]
- Colasuonno, P.; Lozito, M.L.; Marcotuli, I.; Nigro, D.; Giancaspro, A.; Mangini, G.; De Vita, P.; Mastrangelo, A.M.; Pecchioni, N.; Houston, K.; et al. The carotenoid biosynthetic and catabolic genes in wheat and their association with yellow pigments. BMC Genom. 2017, 18, 122. [Google Scholar] [CrossRef] [Green Version]
- Colasuonno, P.; Marcotuli, I.; Blanco, A.; Maccaferri, M.; Condorelli, G.E.; Tuberosa, R.; Parada, R.; de Camargo, A.C.; Schwember, A.R.; Gadaleta, A. Carotenoid pigment contentin durum wheat (Triticum turgidum l var durum), an overview of quantitative trait loci and candidate genes. Front. Plant Sci. 2019, 10, 1347. [Google Scholar] [CrossRef] [Green Version]
- Marcotuli, I.; Colasuonno, P.; Cutillo, S.; Simeone, R.; Blanco, A.; Gadaleta, A. β-glucan content in a panel of Triticum and Aegilops genotypes. Genet. Resour. Crop Evol. 2019, 66, 897–907. [Google Scholar] [CrossRef]
- Colasuonno, P.; Marcotuli, I.; Cutillo, S.; Simeone, R.; Blanco, A.; Gadaleta, A. Effect of barley chromosomes on the β-glucan content of wheat. Genet. Resour. Crop Evol. 2020, 67, 561–567. [Google Scholar] [CrossRef]
- Seib, P.A. Wheat starch: Isolation, structure and properties. J. Appl. Glycoscience 1994, 41, 49–69. [Google Scholar]
- Limbalkar, O.M.; Sharma, J.B.; Jha, S.K.; Mallick, N.; Niranjana, M.; Vinod, N. Mapping of genes for leaf and stem rust resistance in bread wheat genotype selection 212. Indian J. Genet. Plant Breed. 2019, 79, 1–8. [Google Scholar] [CrossRef]
- Kthiri, D.; Loladze, A.; N’Diaye, A.; Nilsen, K.T.; Walkowiak, S.; Dreisigacker, S.; Ammar, K.; Pozniak, C.J. Mapping of genetic loci conferring resistance to leaf rust from three globally resistant durum wheat sources. Front. Plant Sci. 2019, 10, 1247. [Google Scholar] [CrossRef]
- Kang, Y.; Zhou, M.; Merry, A.; Barry, K. Mechanisms of powdery mildew resistance of wheat—A review of molecular breeding. Plant Pathol. 2020, 69, 601–617. [Google Scholar] [CrossRef]
- Gadaleta, A.; Colasuonno, P.; Give, S.L.; Blanco, A.; Giancaspro, A. Map-based cloning of QFhbmgb-2A identifies a WAK2 gene responsible for Fusarium Head Blight resistance in wheat. Sci. Rep. 2019, 9, 6929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royo, C.; Maccaferri, M.; Álvaro, F.; Moragues, M.; Sanguineti, M.C.; Tuberosa, R.; Maalouf, F.; García del Moral, L.F.; Demontis, A.; Rhouma, S.; et al. Understanding the relationships between genetic and phenotypic structures of a collection of elite durum wheat accessions. Field Crops Res. 2010, 119, 91–105. [Google Scholar] [CrossRef]
- Bergkamp, B.; Impa, S.; Asebedo, A.; Fritz, A.; Jagadish, S.K. Prominent winter wheat varieties response to post-flowering heat stress under controlled chambers and field based heat tents. Field Crops Res. 2018, 222, 143–152. [Google Scholar] [CrossRef]
- Hlaváčová, M.; Klem, K.; Rapantová, B.; Novotná, K.; Urban, O.; Hlavinka, P.; Smutna, P.; Horakova, V.; Skarpa, P.; Pohankova, E.; et al. Interactive effects of high temperature and drought stress during stem elongation.; anthesis and early grain filling on the yield formation and photosynthesis of winter wheat. Field Crops Res. 2018, 221, 182–195. [Google Scholar] [CrossRef]
- Ugarte, C.; Calderini, D.F.; Slafer, G.A. Grain weight and grain number responsiveness to pre-anthesis temperature in wheat, barley and triticale. Field Crops Res. 2007, 100, 240–248. [Google Scholar] [CrossRef]
- Royo, C.; Villegas, D.; Rharrabti, Y.; Blanco, R.; Martos, V.; Garcia del Moral, L.F. Grain growth and yield formation of durum wheat grown at contracting latitudes and water regimes in a Mediterranean environment. Cereal Res. Commun. 2006, 34, 1021–1028. [Google Scholar] [CrossRef]
- Altenbach, S.B. New insights into the effects of high temperature.; drought and post-anthesis fertilizer on wheat grain development. J. Cereal Sci. 2012, 56, 39–50. [Google Scholar] [CrossRef]
- Dolferus, R. To grow or not to grow, A stressful decision for plants. Plant Sci. 2014, 229, 247–261. [Google Scholar] [CrossRef]
- Reynolds, M.P.; Mujeeb-Kazi, A.; Sawkins, M. Prospects for utilizing plant-adaptive mechanisms to improve wheat and other crops in drought- and salinity-prone environments. Ann. Appl. Biol. 2005, 146, 239–259. [Google Scholar] [CrossRef]
- Salekdeh, G.H.; Reynolds, M.; Bennett, J.; Boyer, J. Conceptual framework for drought phenotyping during molecular breeding. Trends Plant Sci. 2009, 14, 488–496. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; Chenu, K.; Biddulph, B.; Moeller, C.; Deery, D.M.; Rattey, A.R.; Bennett, D.; Barrett-Lennard, E.G.; Mayer, J.E. A multisite managed environment facility for targeted trait and germplasm phenotyping. Funct. Plant Biol. 2013, 40, 1–13. [Google Scholar] [CrossRef]
- Graybosch, R.A.; Peterson, C.J. Genetic improvement in winter wheat yields in the Great Plains of North America, 1959–2008. Crop Sci. 2010, 50, 1882–1890. [Google Scholar] [CrossRef]
- Bechtold, N.; Bouchez, D. Planta Agrobacterium-Mediated Transformation of Adult Arabidopsis thaliana Plants by Vacuum Infiltration. In Gene Transfer to Plants Springer Lab Manual; Potrykus, I., Spangenberg, I.G., Eds.; Springer: Berlin, Germany, 1995; pp. 19–23. [Google Scholar]
- Zhang, Q.; Bartels, D. Molecular responses to dehydration and desiccation in desiccation-tolerant angiosperm plants. J. Exp. Bot. 2018, 69, 3211–3222. [Google Scholar] [CrossRef] [PubMed]
- Royo, C.; Villegas, D. Field measurements of canopy spectra for biomass assessment of small-grain cereals. In Biomass-Detection, Production and Usage; Matovic, D., Ed.; InTech: Rijeka, Croatia, 2007; ISBN 978-953-307-492-4. [Google Scholar]
- Tuberosa, R. Phenotyping for drought tolerance of crops in the genomics era. Front. Physiol. 2012, 3, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Hassouni, K.; Belkadi, B.; Filali-Maltouf, A.; Tidiane-Sall, A.; Al-Abdallat, A.; Nachit, M.; Bassi, F.M. Loci controlling adaptation to heat stress occurring at the reproductive stage in durum wheat. Agromomy 2019, 9, 414. [Google Scholar] [CrossRef] [Green Version]
- Bento, M.; Pereira, S.G.; Viegas, W.; Silva, M. Durum wheat diversity for heat stress tolerance during inflorescence emergence is correlated to TdHSP101C expression in early developmental stages. PLoS ONE 2017, 28, e0190085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Kouadria, R.; Bouzouina, M.; Azzouz, R.; Lotmani, B. Salinity stress resistance of durum wheat (Triticum durum) enhanced by fungi. Int. J. Biosci. 2018, 12, 70–77. [Google Scholar]
- Longin, F.; Sieber, A.N.; Reif, J.C. Combining frost tolerance, high grain yield and good pasta quality in durum wheat. Plant Breed. 2013, 132, 353–358. [Google Scholar] [CrossRef]
- Soriano, J.M.; Malosetti, M.; Roselló, M.; Sorrells, M.E.; Royo, C. Dissecting the old Mediterranean durum wheat genetic architecture for phenology.; biomass and yield formation by association mapping and QTL meta-analysis. PLoS ONE 2017, 12, e0178290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chardon, F.; Virlon, B.; Moreau, L.; Falque, M.; Joet, J.; Decousset, L.; Murigneux, A.; Charcosset, A. Genetic architecture of flowering time in maize as inferred from quantitative trait loci meta-analysis and synteny conservation with the rice genome. Genetics 2004, 168, 2169–2185. [Google Scholar] [CrossRef] [Green Version]
- Flint-Garcia, S.A.; Thornsberry, J.M.; Buckler, E.S. Structure of linkage disequilibrium in plants. Ann. Rev. Plant Biol. 2003, 54, 357–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breseghello, F.; Sorrells, M.S. Association mapping of kernel size and milling quality in wheat (Triticum aestivum L.) cultivars. Genetics 2006, 172, 1165–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maccaferri, M.; Sanguineti, M.C.; Demontis, A.; El-Ahmed, A.; Garcia del Moral, L.; Maalouf, F.; Nachit, M.; Nserallah, N.; Ouabbou, H.; Rhouma, S.; et al. Association mapping in durum wheat grown across a broad range of water regimes. J. Exp. Bot. 2011, 62, 409–438. [Google Scholar] [CrossRef] [Green Version]
- Laido, G.; Marone, D.; Russo, M.A.; Colecchia, S.A.; Mastrangelo, A.M.; De Vita, P.; Papa, R. Linkage disequilibrium and genome-wide association mapping in tetraploid wheat (Triticum turgidum L.). PLoS ONE 2014, 9, e95211. [Google Scholar] [CrossRef] [Green Version]
- Peleg, Z.; Cakmak, I.; Ozturk, L.; Yazici, A.; Jun, Y.; Budak, H.; Korol, A.B.; Fahima, T.; Saranga, Y. Quantitative trait loci conferring grain mineral nutrient concentrations in durum wheat × wild emmer wheat RIL population. Theor. Appl. Genet. 2009, 119, 353–369. [Google Scholar] [CrossRef] [Green Version]
- Graziani, M.; Maccaferri, M.; Royo, C.; Salvatorelli, F.; Tuberosa, R. QTL dissection of yield components and morpho-physiological traits in a durum wheat elite population tested in contrasting thermo-pluviometric conditions. Crop Pasture Sci. 2014, 65, 80–95. [Google Scholar] [CrossRef]
- Nagel, M.; Navakode, S.; Scheibal, V.; Baum, M.; Nachit, M.; Röder, M.S.; Börner, A. The genetic basis of durum wheat germination and seedling growth under osmotic stress. Biol. Plant 2014, 58, 681–688. [Google Scholar] [CrossRef]
- Maccaferri, M.; El-Feki, W.; Nazemi, G.; Salvi, S.; Cané, M.A.; Colalongo, C.; Stefanelli, S.; Tuberosa, R. Prioritizing quantitative trait loci for root system architecture in tetraploid wheat. J. Exp. Bot. 2016, 67, 1161–1178. [Google Scholar] [CrossRef]
- Iannucci, A.; Marone, D.; Russo, M.A.; De Vita, P.; Miullo, V.; Ferragonio, P.; Blanco, A.; Gadaleta, A.; Mastrangelo, A.M. Mapping QTL for Root and shoot morphological traits in a durum wheat × T dicoccum segregating population at seedling stage. Int. J. Genom. 2017, 6876393. [Google Scholar] [CrossRef] [Green Version]
- Sanguineti, M.C.; Li, S.; Maccaferri, M.; Corneti, S.; Rotondo, F.; Chiari, T.; Tuberosa, R. Genetic dissection of seminal root architecture in elite durum wheat germplasm. Ann. Appl. Biol. 2007, 151, 291–305. [Google Scholar] [CrossRef]
- Canè, M.A.; Maccaferri, M.; Nazemi, G.; Salvi, S.; Francia, R.; Colalongo, C.; Tuberosa, R. Association mapping for root architectural traits in durum wheat seedlings as related to agronomic performance. Mol. Breed. 2014, 34, 1629–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Condorelli, G.E.; Maccaferri, M.; Newcomb, M.; Andrade-Sanchez, P.; White, J.W.; French, A.N.; Sciara, G.; Ward, R.; Tuberosa, R. Comparative aerial and ground based high throughput phenotyping for the genetic dissection of NDVI as a proxy for drought adaptive traits in durum wheat. Front. Plant Sci. 2018, 9, 893. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stack, R.W.; Friesen, T.L.; Faris, J.D. Identification of a novel fusarium head blight resistance quantitative trait locus on chromosome 7A in tetraploid wheat. Phytopathol. 2007, 97, 592–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haile, J.K.; Nachit, M.M.; Hammer, K.; Badebo, A.; Röder, M.S. QTL mapping of resistance to race Ug99 of Puccinia graminis f.sp. tritici in durum wheat (Triticum durum Desf.). Mol. Breed. 2012, 30, 1479–1493. [Google Scholar]
- Maccaferri, M.; Francia, R.; Ratti, C.; Rubies-Autonell, C.; Colalongo, C.; Ferrazzano, G.; Tubersoa, R.; Sanguineti, M.C. Genetic analysis of Soil-Borne Cereal Mosaic Virus response in durum wheat: Evidence for the role of the major quantitative trait locus QSbm.ubo-2BS and of minor quantitative trait loci. Mol. Breed. 2012, 29, 973–988. [Google Scholar] [CrossRef]
- Ben-David, R.; Peleg, Z.; Dinoor, A.; Saranga, Y.; Korol, A.B.; Fahima, T. Genetic dissection of quantitative powdery mildew resistance loci in tetraploid wheat. Mol. Breed. 2014, 34, 1647–1658. [Google Scholar] [CrossRef]
- Zhang, Q.; Axtman, J.E.; Faris, J.D.; Shiaoman, C.; Zengcui, Z.; Timothy, L.F.; Zhong, S.; Cai, X.; Elias, E.M.; Xu, S.S. Identification and molecular mapping of quantitative trait loci for Fusarium head blight resistance in emmer and durum wheat using a single nucleotide polymorphism-based linkage map. Mol. Breed. 2014, 34, 1677–1687. [Google Scholar] [CrossRef]
- Kumar, S.; Knox, R.E.; Singh, A.K.; DePaw, R.M.; Campbell, H.R.; Isidro-Sanchez, J.; Clarke, F.R.; Pozniak, C.J.; N’Daye, A.; Meyer, B.; et al. High-density genetic mapping of a major QTL for resistance to multiple races of loose smut in a tetraploid wheat cross. PLoS ONE 2018, 13, e0192261. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; N’Diaye, A.; Walkowiak, S.; Nilsen, K.T.; Cory, A.T.; Haile, J.; Kutcher, H.R.; Ammar, K.; Loladze, A.; Huerta-Espino, J.; et al. Genetic analysis of resistance to stripe rust in durum wheat (Triticum turgidum L. var. durum). PLoS ONE 2018, 13, e0203283. [Google Scholar] [CrossRef]
- Gordon, A.; McCartney, C.; Knox, R.E.; Ereful, N.; Hiebert, C.W.; Konkin, D.J.; Hsueh, Y.C.; Bhadauria, V.; Sgroi, M.; O’Sullivan, D.M.; et al. Genetic and transcriptional dissection of resistance to Claviceps purpurea in the durum wheat cultivar Greenshank. Theor. Appl. Genet. 2020, 133, 1873–1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghavami, F.; Elias, E.M.; Mamidi, S.; Ansari, O.; Sargolzaei, M.; Adhikari, T.; Mergoum, M.; Kianian, S.F. Mixed model association mapping for fusarium head blight resistance in Tunisian-derived durum wheat populations. G3 2011, 1, 209–218. [Google Scholar] [CrossRef]
- Letta, T.; Olivera, P.; Maccaferri, M.; Jin, Y.; Ammar, K.; Badebo, A.; Salvi, S.; Noli, E.; Crossa, J.; Tuberosa, R. Association mapping reveals novel stem rust resistance loci in Durum wheat at the seedling stage. Plant Genome 2014, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Aoun, M.; Breiland, M.; Kathryn Turner, M.; Loladze, A.; Chao, S.; Xu, S.S.; Ammar, K.; Anderson, J.A.; Kolmer, J.A.; Acevedo, M. Genome-wide association mapping of leaf rust response in a durum wheat worldwide germplasm collection. Plant Genome 2016, 9, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidane, Y.G.; Hailemariam, B.N.; Mengistu, D.K.; Fadda, C.; Pè, M.E.; Dell’Acqua, M. Genome-Wide Association Study of Septoria tritici Blotch Resistance in Ethiopian Durum Wheat Landraces. Front. Plant Sci. 2017, 8, 1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Maccaferri, M.; Bulli, P.; Rynearson, S.; Tuberosa, R.; Chen, X.; Pumphrey, M. Genome-wide association mapping for seedling and field resistance to Puccinia striiformis f. sp. tritici in elite durum wheat. Theor. Appl. Genet. 2017, 130, 649–667. [Google Scholar] [CrossRef]
- Saccomanno, A.; Matny, O.; Marone, D.; Laidò, G.; Petruzzino, G.; Mazzucotelli, E.; Desiderio, F.; Blanco, A.; Gadaleta, A.; Pecchioni, N.; et al. Genetic mapping of loci for resistance to stem rust in a tetraploid wheat collection. Int. J. Mol. Sci. 2018, 19, 3907. [Google Scholar] [CrossRef] [Green Version]
- Steiner, B.; Buerstmayr, M.; Wagner, C.; Danler, A.; Eshonkulov, B.; Ehn, M.; Buerstmayr, H. Fine-mapping of the Fusarium head blight resistance QTL Qfhs.ifa-5A identifies two resistance QTL associated with anther extrusion. Theor. Appl. Genet. 2019, 132, 2039–2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, V.; Roncallo, P.F.; Beaufort, V.; Cervigni, G.L.; Miranda, R.; Jensen, C.A.; Echenique, V.C. Mapping of main and epistatic effect QTLs associated with grain protein and gluten strength using a RIL population of durum wheat. J. Appl. Genet. 2011, 52, 287–298. [Google Scholar] [CrossRef]
- Roncallo, P.F.; Cervigni, G.L.; Jensen, C.; Miranda, R.; Carrera, A.D.; Helguera, M.; Echenique, V. QTL analysis of main and epistatic effects for flour color traits in durum wheat. Euphytica 2012, 185, 77–92. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Xue, W.T.; Yang, R.Z.; Qin, H.B.; Zhao, G.; Tzion, F.; Cheng, J.P. Quantitative trait loci conferring grain selenium nutrient in durum wheat×wild emmer wheat RIL population. Czech. J. Genet. Plant Breed. 2018, 54, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Giancaspro, A.; Give, S.L.; Zacheo, S.A.; Blanco, A.; Gadaleta, A. Genetic Variation for Protein Content and Yield-Related Traits in a Durum Population Derived from an Inter-Specific Cross Between Hexaploid and Tetraploid Wheat Cultivars. Front. Plant Sci. 2019, 10, 1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, Y.; Yu, B.; Knox, R.E.; Singh, A.K.; DePauw, R.; Cuthbert, R.; Zhang, W.; Piche, I.; Gao, P.; Sharpe, A.; et al. High density mapping of quantitative trait loci conferring gluten strength in canadian durum wheat. Front. Plant Sci. 2020, 11, 170. [Google Scholar] [CrossRef] [Green Version]
- Reimer, S.; Pozniak, C.J.; Clarke, F.R.; Clarke, J.M.; Somers, D.J.; Knox, R.E.; Singh, A.K. Association mapping of yellow pigment in an elite collection of durum wheat cultivars and breeding lines. Genome 2008, 51, 1016–1025. [Google Scholar] [CrossRef]
- Marcotuli, I.; Houston, K.; Waugh, R.; Fincher, G.B.; Burton, R.A.; Blanco, A.; Gadaleta, A. Genome wide association mapping for arabinoxylan content in a collection of tetraploid wheats. PLoS ONE 2015, 10, e0132787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- N’Diaye, A.; Haile, J.K.; Cory, A.T.; Clarke, F.R.; Clarke, J.M.; Knox, R.E.; Pozniak, C.J. Single marker and haplotype-based association analysis of semolina and pasta colour in elite durum wheat breeding lines using a high-density consensus map. PLoS ONE 2017, 12, e0170941. [Google Scholar]
- N’Diaye, A.; Haile, J.K.; Nilsen, K.T.; Walkowiak, S.; Ruan, Y.; Singh, A.K.; Clarke, F.R.; Clarke, J.M.; Pozniak, C.J. Haplotype loci under selection in Canadian durum wheat germplasm over 60 years of breeding: Association with grain yield, quality traits, protein loss, and plant height. Front. Plant Sci. 2018, 9, 1–19. [Google Scholar] [CrossRef]
- Rosello, M.; Royo, C.; Alvaro, F.; Villegas, D.; Nazco, R.; Soriano, J.M. Pasta-making quality QTLome from Mediterranean durum wheat landraces. Front. Plant Sci. 2018, 9, 1512. [Google Scholar] [CrossRef]
- Goffinet, B.; Gerber, S. Quantitative Trait Loci: A metaanalysis. Genetics 2000, 155, 463–473. [Google Scholar]
- Löffler, M.; Schön, C.; Miedaner, T. Revealing the genetic architecture of FHB resistance in hexaploid wheat (Triticum aestivum L.) by QTL meta-analysis. Mol. Breed. 2009, 23, 473–488. [Google Scholar] [CrossRef]
- Hanocq, E.; Laperche, A.; Jaminon, O. Most significant genome regions involved in the control of earliness traits in bread wheat.; as revealed by QTL meta-analysis. Theor. Appl. Genet. 2007, 114, 569–584. [Google Scholar] [CrossRef] [PubMed]
- Goudemand, E.; Laurent, V.; Duchalais, L.; Ghaffary, S.M.; Kema, G.H.; Lonnet, P.; Margalé, E.; Robert, O. Association mapping and meta-analysis: Two complementary approaches for the detection of reliable Septoria tritici blotch quantitative resistance in bread wheat (Triticum aestivum L.). Mol. Breed. 2013, 32, 563–584. [Google Scholar] [CrossRef]
- Marone, D.; Russo, M.A.; Laidò, G.; De Vita, P.; Papa, R.; Blanco, A.; Gadaleta, A.; Rubiales, D.; Mastrangelo, A.M. Genetic basis of qualitative and quantitative resistance to powdery mildew in wheat: From consensus regions to candidate genes. BMC Genom. 2013, 14, 562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soriano, J.M.; Royo, C. Dissecting the genetic architecture of leaf rust resistance in wheat by QTL meta-analysis. Phytopathology 2015, 105, 1585–1593. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, S.; Simmonds, J.; Leverington, M.; Wang, Y.; Fish, L.J.; Sayers, L.; Alibert, L.; Orford, S.; Wingen, L.; Snape, J. Meta-QTL analysis of the genetic control of crop height in elite European winter wheat germplasm. Mol. Breed. 2012, 29, 159–171. [Google Scholar] [CrossRef]
- Quraishi, U.M.; Murat, F.; Abrouk, M.; Pont, C.; Confolent, C.; Oury, F.X.; Ward, J.; Boros, D.; Gebruers, K.; Delcour, J.A.; et al. Combined meta-genomics analyses unravel candidate genes for the grain dietary fiber content in bread wheat (Triticum aestivum L.). Funct. Integr. Genom. 2011, 11, 71–83. [Google Scholar] [CrossRef]
- Tyagi, S.; Mir, R.R.; Balyan, H.S.; Gupta, P.K. Interval mapping and meta-QTL analysis of grain traits in common wheat (Triticum aestivum L.). Euphytica 2015, 201, 367–380. [Google Scholar] [CrossRef]
- Darzi-Ramandi, H.; Shariati, J.V.; Tavakol, E.; Najafi-Zarini, H.; Bilgrami, S.S.; Razavi, K. Detection of consensus genomic regions associated with root architecture of bread wheat on groups 2 and 3 chromosomes using QTL meta-analysis. Aust. J. Crop Sci. 2017, 11, 777. [Google Scholar] [CrossRef]
- Soriano, J.M.; Alvaro, F. Discovering consensus genomic regions in wheat for root-related traits by QTL meta-analysis. Sci. Rep. 2019, 9, 10537. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, S.; Gupta, P.K. Meta-analysis of QTLs involved in pre-harvest sprouting tolerance and dormancy in bread wheat. Triticeae Genom. Genet. 2012, 3, 9–24. [Google Scholar] [CrossRef]
- Liu, R.; Jia, H.; Cao, X.; Huang, J.; Li, F.; Tao, Y.; Qiu, F.; Zheng, Y.; Zhang, Z. Fine mapping and candidate gene prediction of a pleiotropic quantitative trait locus for yield-related trait in Zea mays. PLoS ONE 2012, 7, e49836. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Gonzalez, R.H.; Segovia, V.; Bird, N.; Fenwick, P.; Holdgate, S.; Berry, S.; Jack, P.; Caccamo, M.; Uauy, C. RNA-Seq bulked segregant analysis enables the identification of high-resolution genetic markers for breeding in hexaploid wheat. Plant Biotechnol. 2015, 13, 613–624. [Google Scholar] [CrossRef]
- Zou, C.; Wang, P.; Xu, Y. Bulked sample analysis in genetics.; genomics and crop improvement. Plant Biotechnol. 2016, 10, 1941–1955. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xie, J.; Zhang, H.; Guo, B.; Ning, S.; Chen, Y.; Lu, P.; Wu, Q.; Li, M.; Zhang, D.; et al. Mapping stripe rust resistance gene YrZH22 in Chinese wheat cultivar Zhoumai 22 by bulked segregant RNA-Seq (BSR-Seq) and comparative genomics analyses. Theor. Appl. Genet. 2017, 130, 2191–2201. [Google Scholar] [CrossRef] [PubMed]
- Bassi, F.M.; Ghavami, F.; Hayden, M.J.; Wang, Y.; Forrest, K.L.; Kong, S.; Dizon, R.; Michalak de Jimenez, M.K.; Meinhardt, S.W.; Mergoum, M.; et al. Fast-forward genetics by radiation hybrids to saturate the locus regulating nuclear-cytoplasmic compatibility in Triticum. Plant Biotechnol. J. 2016, 14, 1716–1726. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.K.; Kulwal, P.L.; Mir, R.R. QTL mapping, methodology and applications in cereal breeding. In Cereal Genomics II; Gupta, P.K., Varshney, R.K., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 275–318. [Google Scholar]
- Huang, B.E.; Verbyla, K.L.; Verbyla, A.P. MAGIC populations in crops: Current status and future prospects. Theor. Appl. Genet. 2015, 128, 999–1017. [Google Scholar] [CrossRef] [PubMed]
- Milner, S.G.; Maccaferri, M.; Huang, B.E.; Mantovani, P.; Massi, A.; Frascaroli, E.; Tuberosa, R.; Salvi, S. A multiparental cross population for mapping QTL for agronomic traits in durum wheat (Triticum turgidum ssp. durum). Plant Biotechnol. J. 2015, 14, 735–748. [Google Scholar] [CrossRef] [Green Version]
- Vatter, T.; Maurer, A.; Kopahnke, D.; Perovic, D.; Ordon, F.; Pillen, K. A nested association mapping population identifies multiple small effect QTL conferring resistance against net blotch (Pyrenophora teres f teres) in wild barley. PLoS ONE 2017, 12, 0186803. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Fang, X.; Jiang, P.; Zhang, G.; Hu, J.; Wang, X.; Meng, Q.; Cui, W.; Lan, S.; Ma, X.; et al. Genetic architecture of nitrogen-deficiency tolerance in wheat seedlings based on a nested association mapping (NAM) population. Front. Plant Sci. 2018, 9, 845. [Google Scholar] [CrossRef]
- McMullen, M.D.; Kresovich, S.; Villeda, H.S.; Bradbury, P.; Li, H.; Sun, Q.; Flint-Garcia, S.; Thornsberry, J.; Acharya, C.; Bottoms, C.; et al. Genetic properties of the maize nested association mapping population. Science 2009, 325, 737–740. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Holland, J.B.; McMullen, M.D.; Buckler, E.S. Genetic design and statistical power of nested association mapping in maize. Genetics 2008, 178, 539–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Cao, Y.; He, J.; Zhao, T.; Gai, J. Detecting the QTL-allele system conferring flowering date in a nested association mapping population of soybean using a novel procedure. Theor. Appl. Genet. 2017, 130, 2297–2314. [Google Scholar] [CrossRef]
- Bouchet, S.; Olatoye, M.O.; Marla, S.R.; Perumal, R.; Tesso, T.; Yu, J.; Tuinstra, M.; Morris, G.P. Increased power to dissect adaptive traits in global sorghum diversity using a nested association mapping population. Genetics 2017, 206, 573–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nice, L.M.; Steffenson, B.J.; Blake, T.K.; Horsley, R.D.; Smith, K.P.; Muehlbauer, G.J. Mapping agronomic traits in a wild barley advanced backcross-nested association mapping population. Crop Sci. 2017, 57, 1199–2210. [Google Scholar] [CrossRef]
- Bajgain, P.; Rouse, M.N.; Tsilo, T.J.; Macharia, G.K.; Bhavani, S.; Jin, Y.; Anderson, J.A. Nested association mapping of stem rust resistance in wheat using genotyping by sequencing. PLoS ONE 2016, 11, e0155760. [Google Scholar] [CrossRef]
- Alahmad, S.; El Hassouni, K.; Bassi, F.M.; Dinglasan, E.; Youssef, C.; Quarry, G.; Aksoy, A.; Mazzucotelli, E.; Juhász, A.; Able, J.A.; et al. A Major Root Architecture QTL Responding to Water Limitation in Durum Wheat. Front. Plant Sci. 2019, 10, 436. [Google Scholar] [CrossRef] [Green Version]
- Kidane, Y.G.; Gesesse, C.A.; Hailemariam, B.N.; Desta, E.A.; Mengistu, D.K.; Fadda, C.; Pè, M.E.; Dell’Acqua, M. A large nested association mapping population for breeding and quantitative trait locus mapping in Ethiopian durum wheat. Plant Biotechnol. J. 2019, 17, 1380–1393. [Google Scholar] [CrossRef]
- Alahmad, S.; Kang, Y.; Dinglasan, E.; Mazzucotelli, E.; Voss-Fels, K.P.; Able, J.A.; Christopher, J.; Bassi, F.M.; Hickey, L.T. Adaptive Traits to Improve Durum Wheat Yield in Drought and Crown Rot Environments. Int. J. Mol. Sci. 2020, 21, 5260. [Google Scholar] [CrossRef]
- Soriano, J.; Villegas, D.; Aranzana, M.; García del Moral, L.; Royo, C. Genetic structure of modern durum wheat cultivars and mediterranean landraces matches with their agronomic performance. PLoS ONE 2016, 11, e0160983. [Google Scholar] [CrossRef] [Green Version]
- Soriano, J.M.; Villegas, D.; Sorrells, M.E.; Royo, C. Durum Wheat Landraces from East and West Regions of the Mediterranean Basin Are Genetically Distinct for Yield Components and Phenology. Front. Plant Sci. 2018, 9, 80. [Google Scholar] [CrossRef] [Green Version]
- Rommens, J.M.; Iannuzzi, M.C.; Kerem, B.; Drumm, M.L.; Melmer, G.; Dean, M.; Rozmahel, R.; Cole, J.L.; Kennedy, D.; Hidaka, N.; et al. Identification of the cystic fibrosis gene: Chromosome walking and jumping. Science 1989, 245, 1059–1065. [Google Scholar] [CrossRef]
- Bechtold, U.; Ferguson, J.N.; Mullineaux, P.M. To defend or to grow, lessons from Arabidopsis C24. J. Exp. Bot. 2018, 69, 2809–2821. [Google Scholar] [CrossRef] [PubMed]
- Byrne, P.F.; McMullen, M.D. Defining genes for agricultural traits, QTL analysis and the candidate gene approach. Probe 1996, 7, 24–27. [Google Scholar]
- De Vienne, D. La génomique pour l’étude des gènes contrôlant la variation des caractères d’intérêt agronomique. Oléagineux Corps Gras Lipides 1999, 6, 143–148. [Google Scholar]
- Pflieger, S.; Lefebvre, V.; Causse, M. The candidate gene approach in plant genetics: A review. Mol. Breed. 2001, 7, 275–291. [Google Scholar] [CrossRef]
- Faris, J.D.; Li, W.L.; Liu, D.J.; Chen, P.D.; Gill, B.S. Candidate gene analysis of quantitative disease resistance in wheat. Theor. Appl. Genet. 1999, 98, 219–225. [Google Scholar] [CrossRef]
- Marcotuli, I.; Colasuonno, P.; Blanco, A.; Gadaleta, A. Expression analysis of cellulose synthase-like genes in durum wheat. Sci. Rep. 2018, 8, 15675. [Google Scholar] [CrossRef] [PubMed]
- James, R.A.; von Caemmerer, S.; Condon, A.G.; Zwart, A.B.; Munns, R. Genetic variation in tolerance to the osmotic stress component of salinity stress in durum wheat. Funct. Plant Biol. 2008, 35, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-González, R.H.; Borrill, P.; Lang, D.; Harrington, S.A.; Brinton, J.; Venturini, L.; Davey, M.; Jacobs, J.; van Ex, F.; Pasha, A.; et al. The transcriptional landscape of polyploid wheat. Science 2018, 361, eaar6089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyers, B.T. Camoco: A net for the sea of candidate genes. Plant Cell 2018, 30, 2889. [Google Scholar] [CrossRef] [Green Version]
- Lin, F.; Lazarus, E.Z.; Rhee, S.Y. QTG-Finder2: A Generalized Machine-Learning Algorithm for Prioritizing QTL Causal Genes in Plants. G3 2020, 10, 2411–2421. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef]
- Gao, L.; Wang, S.; Li, X.Y.; Wei, X.J.; Zhang, Y.J.; Wang, H.Y.; Liu, D.Q. Expression and functional analysis of a pathogenesis-related protein 1 gene, TcLr19PR1, involved in wheat resistance against leaf rust fungus. Plant Mol. Biol. Rep. 2015, 33, 797–805. [Google Scholar] [CrossRef]
- Liu, W.; Frick, M.; Huel, R.; Nykiforuk, C.L.; Wang, X.; Gaudet, D.A.; Eudes, F.; Conner, R.L.; Kuzyk, A.; Chen, Q.; et al. The stripe rust resistance gene Yr10 encodes an evolutionary-conserved and unique CC-NBS-LRR sequence in wheat. Mol. Plant 2014, 7, 1740–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.N.; Shen, F.D.; Hong, Y.T.; Rong, W.; Du, L.P.; Liu, X.; Xu, H.; Ma, L.; Zhang, Z. The wheat calcium-dependent protein kinase TaCPK7-D positively regulates host resistance to sharp eyespot disease. Mol. Plant Pathol. 2016, 17, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, X.; Wang, X.; Zhou, M.; Zhou, X.; Ye, X.; Wei, X. An R2R3 MYB transcription factor in wheat, TaPIMP1, mediates host resistance to Bipolaris sorokiniana and drought stresses through regulation of defense- and stress-related genes. New Phytol. 2012, 196, 1155–1170. [Google Scholar] [CrossRef]
- Liang, Z.; Chen, K.; Li, T.; Zhang, Y.; Wang, Y.; Zhao, Q.; Liu, J.; Zhang, H.; Liu, C.; Ran, Y.; et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 2017, 8, 14261. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, Z.; Zong, Y.; Wang, Y.P.; Liu, J.X.; Chen, K.L.; Qiu, J.L.; Gao, C. Efficient and transgene-free genome editing in wheat through transient expression of CRISPR/Cas9 DNA or RNA. Nat. Commun. 2016, 7, 12617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, Y.; Wang, Y.; Li, C.; Zhang, R.; Chen, K.; Ran, Y.; Qiu, J.L.; Wang, D.; Gao, C. Precise base editing in rice, wheat and maize with a Cas9-cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 438–440. [Google Scholar] [CrossRef]
- Camerlengo, F.; Frittelli, A.; Sparks, C.; Doherty, A.; Martignago, D.; Larré, C.; Lupi, R.; Sestili, F.; Masci, F. CRISPR-Cas9 multiplex editing of the α-amylase/trypsin inhibitor genes to reduce allergen proteins in durum wheat. Front. Sustain. Food Syst. 2020, 4, 104. [Google Scholar] [CrossRef]
- Kalunke, R.M.; Tundo, S.; Sestili, F.; Camerlengo, F.; Lafiandra, D.; Lupi, R.; Larré, C.; Denery-Papini, S.; Islam, S.; Ma, W.; et al. Reduction of allergenic potential in bread wheat rnai transgenic lines silenced for cm3, cm16 and 0.28 ati genes. Int. J. Mol. Sci. 2020, 21, 5817. [Google Scholar] [CrossRef]
- Loss, S.P.; Siddique, K.H.M. Morphological and Physiological Traits Associated with Wheat Yield Increases in Mediterranean Environments. Adv. Agron. 1994, 52, 229–276. [Google Scholar]
- Sanchez-Garcia, M.; Royo, C.; Aparicio, N.; Martín-Sánchez, J.A.; Alvaro, F. Genetic improvement of bread wheat yield and associated traits in Spain during the 20th century. J. Agric. Sci. 2013, 151, 105–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Garcia, M.; Álvaro, F.; Peremarti, A.; Martín-Sánchez, J.A.; Royo, C. Changes in bread-making quality attributes of bread wheat varieties cultivated in Spain during the 20th century. Eur. J. Agron. 2015, 63, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Royo, C.; Martos, V.; Ramdani, A.; Villegas, D.; Rharrabti, Y.; García Del Moral, L.F. Changes in yield and carbon isotope discrimination of Italian and Spanish durum wheat during the 20th century. Agron. J. 2008, 100, 352–360. [Google Scholar] [CrossRef]
- Jackson, P.; Robertson, M.; Cooper, M.; Hammer, G. The role of physiological understanding in plant breeding; from a breeding perspective. Field Crops Res. 1996, 49, 1–37. [Google Scholar] [CrossRef]
- Goddard, M.; Hayes, B. Mapping genes for complex traits in domestic animals and their use in breeding programmes. Nat. Rev. Genet. 2009, 10, 381–391. [Google Scholar] [CrossRef]
- Isidro, J.; Akdemir, D.; Burke, J. Genomic selection. In The World Wheat Book: A History of Wheat Breeding; William, A., Alain, B., Maarten, V.G., Eds.; Lavoisier: Paris, France, 2016; pp. 1001–1023. [Google Scholar]
- Fiedler, J.D.; Salsman, E.; Liu, Y.; Michalak de Jiménez, M.; Hegstad, J.B.; Chen, B.; Manthey, F.A.; Chao, S.; Xu, S.; Elias, E.M.; et al. Genome-Wide Association and Prediction of Grain and Semolina Quality Traits in Durum Wheat Breeding Populations. Plant Genome 2017, 10, 3. [Google Scholar] [CrossRef] [Green Version]
- Haile, J.K.; N’Diaye, A.; Clarke, F.; Clarke, J.; Knox, R.; Rutkosky, J.; Bassi, F.M.; Pozniak, C.J. Genomic selection for grain yield and quality traits in durum wheat. Mol. Breed. 2018, 38, 75. [Google Scholar] [CrossRef]
- Zaïm, M.; Kabbaj, H.; Kehel, Z.; Gorjanc, G.; Filali-Maltouf, A.; Belkadi, B.; Nachit, M.M.; Bassi, F.M. Combining QTL Analysis and Genomic Predictions for Four Durum Wheat Populations Under Drought Conditions. Front. Genet. 2020, 11, 316. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Amores, J.; Michel, S.; Miedaner, T.; Longin, C.F.H.; Buerstmayr, H. Genomic predictions for Fusarium head blight resistance in a diverse durum wheat panel: An effective incorporation of plant height and heading date as covariates. Euphytica 2020, 216, 22. [Google Scholar] [CrossRef] [Green Version]
- Watson, A.; Ghosh, S.; Williams, M.; Cuddy, W.S.; Simmonds, J.; Rey, M.D.; Asyraf Md Hatta, M.; Hinchliffe, A.; Steed, A.; Reynolds, D.; et al. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alahmad, S.; Dinglasan, E.; Leung, K.M.; Riaz, A.; Derbal, N.; Voss-Fels, K.P.; Hickey, L.T. Speed breeding for multiple quantitative traits in durum wheat. Plant Methods 2018, 14, 36. [Google Scholar] [CrossRef] [PubMed]


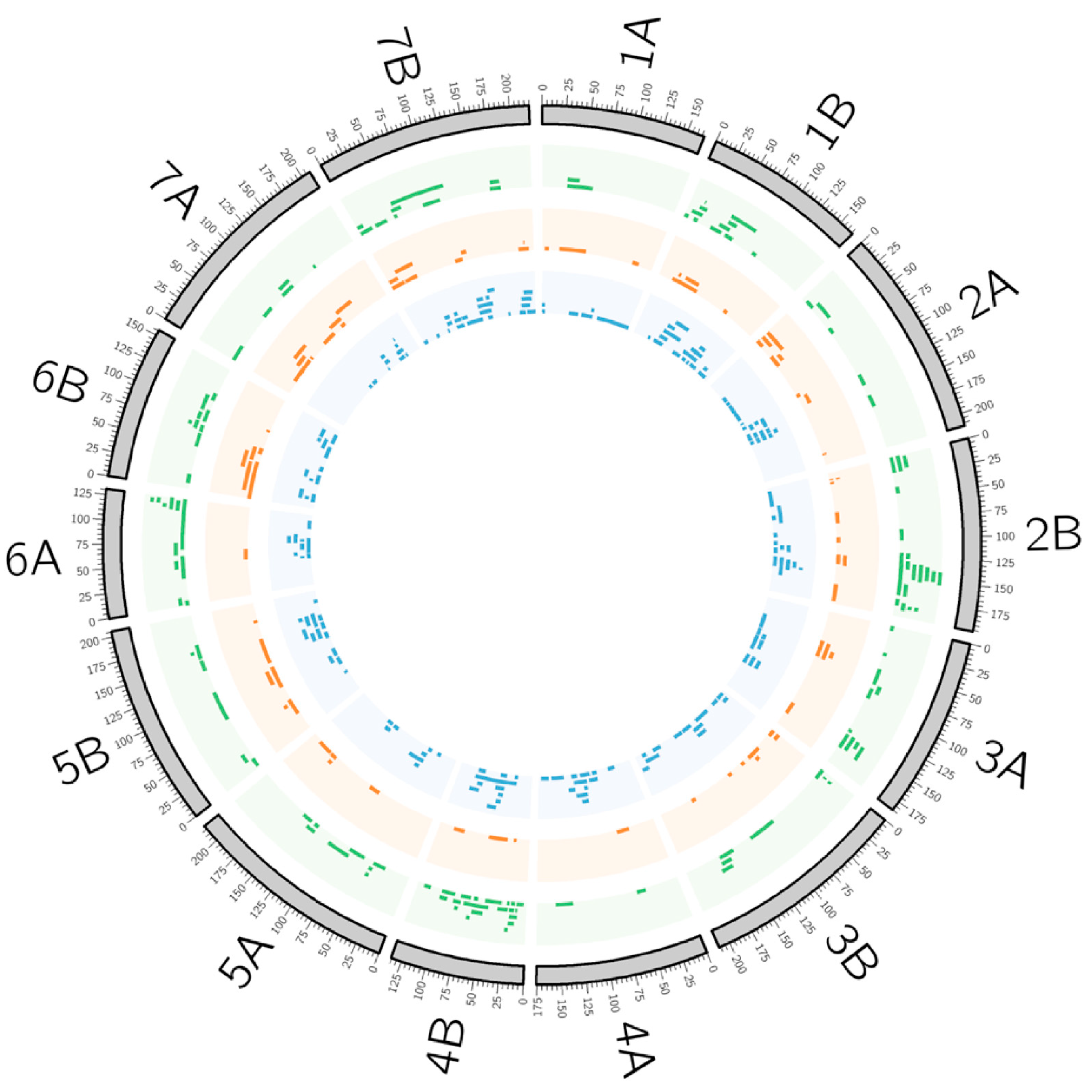{kind=link}
| Cross | Type | N Progenies | N Markers | Marker Type | Distance (cM) | Reference |
|---|---|---|---|---|---|---|
| Messapia × MG4343 | RIL | 65 | 213 | RFLP, biochemical | 1352 | [39] |
| Messapia × MG4343 | RIL | 65 | 292 | SSR, RFLP, biochemical | 2034 | [43] |
| Messapia × MG4343 | RIL | 65 | 293 | AFLP, RFLP, biochemical | 2063 | [44] |
| Jennah Khetifa × Cham 1 | RIL | 110 | 306 | AFLP, SSP, SSR | 3598 | [45] |
| Messapia × MG4343 | RIL | 65 | 458 | AFLP, RFLP, SSR, biochemical, morphological | 1352 | [46] |
| Omrabi 5 × 600545 | RIL | 114 | 295 | AFLP, SSP, SSR | 2289 | [47] |
| Strongfield × Blackbird | DH | 85 | 424 | SSR | 2052 | [48] |
| W9262-260D3 × Kofa | DH | 155 | 194 | DArT, SSR | 1400 | [49] |
| Colosseo × Lloyd | RIL | 176 | 554 | DArT, SSR | 2022 | [50] |
| Langdon × G1816 | RIL | 152 | 669 | DArT, SSR | 2317 | [51] |
| Kofa × UC1113 | RIL | 93 | 269 | SNP, SSR | 2140 | [52] |
| Svevo × Ciccio | RIL | 120 | 522 | SSR | 1605 | [11] |
| Creso × Pedroso | RIL | 123 | 456 | DArT, SSR | 1800 | [53] |
| DT695 × Strongfield | DH | 185 | 345 | DArT, SSR | 1474 | [54] |
| Latino × Primadur | RIL | 121 | 454 | DArT, SSR | 1172 | [10] |
| Neodur × Cirillo | RIL | 146 | 414 | DArT, SSR, STS | 1917 | [55] |
| Svevo × Ciccio | RIL | 120 | 833 | DArT, SSR, morphological | 1716 | [7] |
| DS × Td161 | BC | 134 | 368 | AFLP, SSR | 1921 | [56] |
| Floradur × Td161 | BC | 129 | 248 | AFLP, SSR | 1414 | [56] |
| Helidur × Td161 | BC | 126 | 239 | AFLP, SSR | 1515 | [56] |
| BGRC3487/2 × DT735 | RIL | 160 | 948 | DArT, SSR | 1089 | [57] |
| Rugby × Maier | DH | 105 | 228 | DArT, SSR | 801 | [6] |
| PDW233 × Bhalegaon 4 | RIL | 140 | 245 | SSR, ISSR, TRAP, SCAR, STS, biochemical | 2773 | [58] |
| Gerizim #36 × Helidur | RIL | 103 | 421 | AFLP, SSR | 1808 | [59] |
| Wollaroi × Bansi | RIL | 92 | 799 | DArT, SSR | 3859 | [60] |
| Svevo × Ciccio | RIL | 120 | 5670 | DArT, SNP, SSR | 1774 | [5] |
| Ben × PI41025 | RIL | 200 | 2593 | SNP, SSR, STS | 2444 | [61] |
| Simeto × Molise Colli | RIL | 136 | 9040 | SNP | 2879 | [62] |
| Latino × MG5323 | RIL | 110 | 10840 | SNP, SSR | 2363 | [63] |
| Kofa × Svevo | RIL | 249 | 311 | SNP, SSR | 1259 | [4] |
| Gallareta × Demetra | DH | 127 | 147 | DArT, SSR | 1017 | [4] |
| DT707 × DT696 | DH | 127 | 136 | DArT, SSR | 861 | [4] |
| DT712 × Blackbird | DH | 89 | 392 | SSR | 1848 | [4] |
| Lebsock × PI94749 | DH | 146 | 240 | SSR, biochemical | 1463 | [4] |
| PDW1216 × MvTD10-98 | RIL | 182 | 440 | SSR | 984 | [4] |
| W9292-260D3 × Kofa | DH | 155 | 3712 | SNP, SSR | 1685 | [31] |
| Svevo × Zavitan | RIL | 140 | 10911 | SNP | 2258 | [31] |
| Simeto × Levante | RIL | 180 | 5978 | DArT, SNP, SSR | 2185 | [31] |
| Mohawk × Cocorit69 | RIL | 81 | 5554 | SNP | 2013 | [31] |
| Meridiano × Claudio | RIL | 180 | 5970 | DArT, SNP, SSR | 2239 | [31] |
| Colosseo × Lloyd | RIL | 176 | 7946 | DArT, SNP, SSR | 2064 | [31] |
| Kunduru-1149 × Cham 1 | RIL | 141 | 395 | AFLP, SSP, SSR | 4854 | [64] |
| 02-5B-318 × Saragolla | RIL | 135 | 4366 | SNP | 4227 | [30] |
| Karur × DBC-480 | RIL | 110 | 1609 | DArTseq, SSR | 2806 | [65] |
| Durobonus × DBC-480 | RIL | 100 | 1052 | DArTseq, SSR | 1781 | [65] |
| SZD1029K × DBC-480 | RIL | 100 | 1006 | DArTseq, SSR | 2219 | [65] |
| Bairds × Atred#1 | RIL | 137 | 1150 | SNP, SSR | 2639 | [66] |
| W9262-260D3 × Kofa | DH | 155 | 4227 | SNP | 2282 | [67] |
| Duilio × Avonlea | RIL | 134 | 5444 | SNP | 1962 | [68] |
| Ofanto × Cappelli | RIL | 98 | 9267 | DArT, DArTseq, SSR | 2119 | [69] |
| Joppa × 10Ae564 | RIL | 205 | 5216 | SNP | 3428 | [70] |
| DT707 × DT696 | DH | 127 | 2943 | SNP | 1808 | [71] |
| Strongfield × Blackbird | DH | 85 | 9568 | SNP | 2763 | [71] |
| Svevo × Y12-3 | RIL | 208 | 4166 | SNP | 2169 | [72] |
| Faraj × Gidara 2 | RIL | 143 | 2067 | SNP | 2578 | [73] |
| Tunisian 108 × Ben | BIL | 178 | 329 | DArT, SSR | 1888 | [74] |
| Zardak × Iran249 | RIL | 118 | 6195 | SNP, SSR | 2884 | [75] |
| Rusty × PI193883 | RIL | 190 | 9346 | SNP, SSR | 2440 | [76] |
| Rusty × PI192051-1 | RIL | 180 | 1138 | SNP | 1436 | [77] |
| Rusty × PI 387336 | RIL | 200 | 2894 | SNP | 2858 | [78] |
| Rusty × PI 387696 | RIL | 200 | 2059 | SNP | 2724 | [78] |
| Rusty × PI 466979 | RIL | 200 | 3692 | SNP | 2776 | [78] |
| Rusty × Iumillo | RIL | 200 | 2911 | SNP | 2968 | [78] |
| Reference | Cross | Type | N Genotypes | N QTL | Traits |
|---|---|---|---|---|---|
| Abiotic stress | |||||
| [121] | Langdon × G18-16 | RIL | 156 | 31 | CIR, OP, CC, FLRI |
| [122] | Kofa × Svevo | RIL | 247 | 12 | PDL, SPAD, NDVI |
| [123] | Omrabi5 × Belikh2 | RIL | 114 | 6 | CL, RRT |
| [124] | Colosseo × Lloyd | RIL | 176 | 28 | RRT |
| [124] | Meridiano × Caludio | RIL | 181 | 32 | RRT |
| [125] | Simeto × Mollise Colli | RIL | 136 | 18 | RRT |
| [126] | Elite cultivars | GWAS | 57 | 4 | RRT |
| [127] | Elite cultivars | GWAS | 183 | 2 | RRT |
| [124] | Elite cultivars | GWAS | 183 | 31 | RRT |
| [128] | UNIBO-DP | GWAS | 248 | 73 | DB, NDVI, SPAD |
| Biotic stress | |||||
| [48] | Strongfield × Blackbird | DH | 85 | 2 | FHB |
| [129] | LDN × LDN-Dic7A | RIL | 118 | 1 | FHB |
| [8] | Colosseo × Lloyd | RIL | 176 | 1 | LR |
| [119] | Meridiano × Claudio | RIL | 181 | 1 | SBCMV |
| [56] | DS × Td161 | BC | 134 | 1 | FHB |
| [56] | Floradur × Td161 | BC | 129 | 3 | FHB |
| [56] | Helidur × Td161 | BC | 126 | 1 | FHB |
| [130] | Kristal × Sebatel | RIL | 85 | 7 | SR |
| [131] | Simeto × Levante | RIL | 180 | 7 | SBCMV |
| [57] | BGRC3487 × 2 * DT735 | RIL | 160 | 2 | FHB |
| [55] | Neodur × Cirillo | RIL | 146 | 2 | SBCMV |
| [60] | Wollaroi × Bansi | RIL | 92 | 2 | YR |
| [59] | Gerizim × Helidur | RIL | 103 | 1 | FHB |
| [132] | Langdon × G18-16 | RIL | 157 | 4 | PM |
| [63] | Latino × MG5323 | RIL | 110 | 3 | LR |
| [133] | Ben × PI41025 | RIL | 200 | 3 | FHB |
| [94] | Sumai-3 × Saragolla | RIL | 135 | 11 | FHB |
| [65] | Karur × DBC-480 | RIL | 111 | 1 | FHB |
| [134] | Strongfield × Blackbird | DH | 90 | 2 | LS |
| [135] | Kofa × W9262-260D3 | DH | 155 | 1 | YR |
| [70] | Joppa × 10Ae564 | RIL | 205 | 3 | FHB |
| [77] | Rusty × PI 192051-1 | RIL | 180 | 5 | LR |
| [74] | Ben × Tunisian 108 | BIL | 171 | 3 | FHB |
| [136] | Greenshank × AC Avonlea | DH | 132 | 4 | CP |
| [137] | Different sources | GWAS | 323 | 3 | FHB |
| [138] | Elite cultivars | GWAS | 183 | 8 | SR |
| [139] | Worldwide collection | GWAS | 496 | 50 | LR |
| [140] | Ethiopian landraces | GWAS | 318 | 20 | STB |
| [141] | Elite cultivars | GWAS | 250 | 16 | YR |
| [135] | Elite cultivars | GWAS | 92 | 1 | YR |
| [142] | Tetraploid panel | GWAS | 230 | 37 | SR |
| [143] | Spring lines | GWAS | 228 | 7 | FHB |
| Quality | |||||
| [81] | UC1113 × Kofa | BP | 93 | 5 | YPC |
| [121] | Langdon × G18-16 | RIL | 152 | 55 | GCaC, GCuC, GFeC, GKC, GMgC, GMnC, GPC, GSC, GZnC, PGC |
| [54] | DT695 × Strongfield | DH | 185 | 6 | GPC |
| [10] | Latino × Primadur | BP | 121 | 4 | YPC |
| [144] | UC1113 × Kofa | RIL | 93 | 18 | GPC, SV |
| [145] | UC1113 × Kofa | BP | 93 | 13 | F, YPC |
| [5] | Svevo × Ciccio | BP | 120 | 7 | YPC |
| [68] | Duilio × Avonlea | RIL | 134 | 2 | BG |
| [146] | Langdon × G18-16 | RIL | 152 | 15 | GSeC, GSeY |
| [87] | Colosseo × Lloyd | BP | 176 | 12 | YPC |
| [87] | Kofa × Svevo | BP | 249 | 4 | YPC |
| [87] | Meridiano × Claudio | BP | 181 | 6 | YPC |
| [72] | Svevo × Y12-3 | RIL | 208 | 9 | GPC |
| [147] | Saragolla × 02-5B-318 | RIL | 135 | 9 | GPC |
| [148] | Pelissier × Strongfield | DH | 162 | 6 | SV |
| [149] | Worldwide elite collection | GWAS | 93 | 20 | YPC |
| [150] | Agrogen | GWAS | 104 | 19 | AX |
| [35] | Agrogen | GWAS | 230 | 7 | BG |
| [86] | Durum collection | GWAS | 124 | 6 | YPC |
| [151] | Canadian durum wheats | GWAS | 169 | 6 | YPC |
| [152] | Canadian durum lines | GWAS | 192 | 28 | YPC |
| [153] | Mediterranean landraces | GWAS | 172 | 14 | GPC, GS, TW, YPC |
| Linkage QTL Mapping | GWAS |
|---|---|
| Two known ancestors (parents) | Multiple (unknown) ancestors |
| Short known recombination history | Long (unknown) recombination history |
| Simple population structure | Complex population structure |
| LD caused by linkage | LD caused by different genetic events |
| Requires construction of specific maps | Existing maps can be used |
| Contrasting genetic background | Diverse genetic background |
| Phenotyping is required for new populations | Phenotyping data might already be available |
| Trait | 1A | 1B | 2A | 2B | 3A | 3B | 4A | 4B | 5A | 5B | 6A | 6B | 7A | 7B | Total | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Abiotic | CC | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 7 | |||||||
| CIR | 1 | 1 | 2 | 1 | 1 | 6 | ||||||||||
| CL | 1 | 1 | 2 | 1 | 5 | |||||||||||
| DB | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | |||||||
| FLRI | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 9 | |||||||
| NDVI | 1 | 3 | 3 | 6 | 1 | 6 | 2 | 3 | 3 | 3 | 4 | 3 | 4 | 5 | 47 | |
| OP | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | ||||||
| PDL | 1 | 1 | 1 | 1 | 4 | |||||||||||
| RRT | 10 | 0 | 6 | 17 | 14 | 4 | 17 | 7 | 3 | 14 | 9 | 7 | 8 | 116 | ||
| SPAD | 1 | 2 | 2 | 2 | 1 | 5 | 2 | 3 | 3 | 1 | 1 | 2 | 25 | |||
| Total | 15 | 7 | 13 | 32 | 19 | 18 | 6 | 28 | 15 | 15 | 21 | 16 | 13 | 19 | 237 | |
| Biotic | CP | 1 | 1 | 1 | 1 | 4 | ||||||||||
| FHB | 1 | 2 | 7 | 3 | 3 | 5 | 2 | 2 | 4 | 2 | 4 | 4 | 3 | 42 | ||
| LR | 6 | 1 | 7 | 9 | 4 | 6 | 4 | 3 | 5 | 1 | 4 | 2 | 4 | 3 | 59 | |
| LS | 1 | 1 | 2 | |||||||||||||
| PM | 1 | 1 | 1 | 1 | 4 | |||||||||||
| SBCMV | 1 | 3 | 1 | 1 | 1 | 1 | 1 | 1 | 10 | |||||||
| SR | 1 | 3 | 2 | 6 | 1 | 4 | 10 | 0 | 1 | 3 | 6 | 3 | 4 | 8 | 52 | |
| STB | 7 | 1 | 10 | 1 | 1 | 20 | ||||||||||
| YR | 2 | 3 | 1 | 2 | 1 | 1 | 2 | 1 | 4 | 3 | 20 | |||||
| Total | 16 | 9 | 21 | 24 | 19 | 18 | 16 | 7 | 11 | 12 | 12 | 13 | 17 | 18 | 213 | |
| Quality | AX | 2 | 2 | 2 | 1 | 1 | 1 | 1 | 3 | 1 | 1 | 3 | 1 | 19 | ||
| BG | 1 | 3 | 2 | 1 | 2 | 9 | ||||||||||
| Fb | 2 | 1 | 1 | 1 | 2 | 7 | ||||||||||
| GCaC | 1 | 4 | 5 | |||||||||||||
| GCuC | 2 | 5 | 3 | 10 | ||||||||||||
| GFeC | 1 | 5 | 3 | 1 | 10 | |||||||||||
| GKC | 8 | 8 | ||||||||||||||
| GMgC | 2 | 2 | ||||||||||||||
| GMnC | 2 | 2 | ||||||||||||||
| GPC | 2 | 4 | 6 | 4 | 1 | 2 | 4 | 3 | 2 | 3 | 1 | 1 | 1 | 5 | 39 | |
| GS | 1 | 1 | 1 | 3 | ||||||||||||
| GSC | 1 | 4 | 5 | |||||||||||||
| GSeC | 2 | 2 | 1 | 4 | 9 | |||||||||||
| GseY | 1 | 2 | 1 | 1 | 1 | 6 | ||||||||||
| GZnC | 6 | 6 | ||||||||||||||
| PGC | 2 | 1 | 3 | |||||||||||||
| SV | 2 | 2 | 2 | 2 | 3 | 1 | 1 | 2 | 1 | 16 | ||||||
| TW | 1 | 1 | 1 | 3 | ||||||||||||
| YPC | 7 | 11 | 5 | 1 | 10 | 6 | 7 | 7 | 6 | 10 | 3 | 22 | 13 | 108 | ||
| Total | 15 | 22 | 23 | 18 | 10 | 20 | 18 | 19 | 15 | 17 | 15 | 16 | 31 | 31 | 270 | |
| 46 | 38 | 57 | 74 | 48 | 56 | 40 | 54 | 41 | 44 | 48 | 45 | 61 | 68 | 720 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Colasuonno, P.; Marcotuli, I.; Gadaleta, A.; Soriano, J.M. From Genetic Maps to QTL Cloning: An Overview for Durum Wheat. Plants 2021, 10, 315. https://doi.org/10.3390/plants10020315
Colasuonno P, Marcotuli I, Gadaleta A, Soriano JM. From Genetic Maps to QTL Cloning: An Overview for Durum Wheat. Plants. 2021; 10(2):315. https://doi.org/10.3390/plants10020315
Chicago/Turabian StyleColasuonno, Pasqualina, Ilaria Marcotuli, Agata Gadaleta, and Jose Miguel Soriano. 2021. "From Genetic Maps to QTL Cloning: An Overview for Durum Wheat" Plants 10, no. 2: 315. https://doi.org/10.3390/plants10020315
APA StyleColasuonno, P., Marcotuli, I., Gadaleta, A., & Soriano, J. M. (2021). From Genetic Maps to QTL Cloning: An Overview for Durum Wheat. Plants, 10(2), 315. https://doi.org/10.3390/plants10020315