
Bacterial Indole-3-Acetic Acid Influences Soil Nitrogen Acquisition in Barley and Chickpea

Abstract
:1. Introduction
2. Results
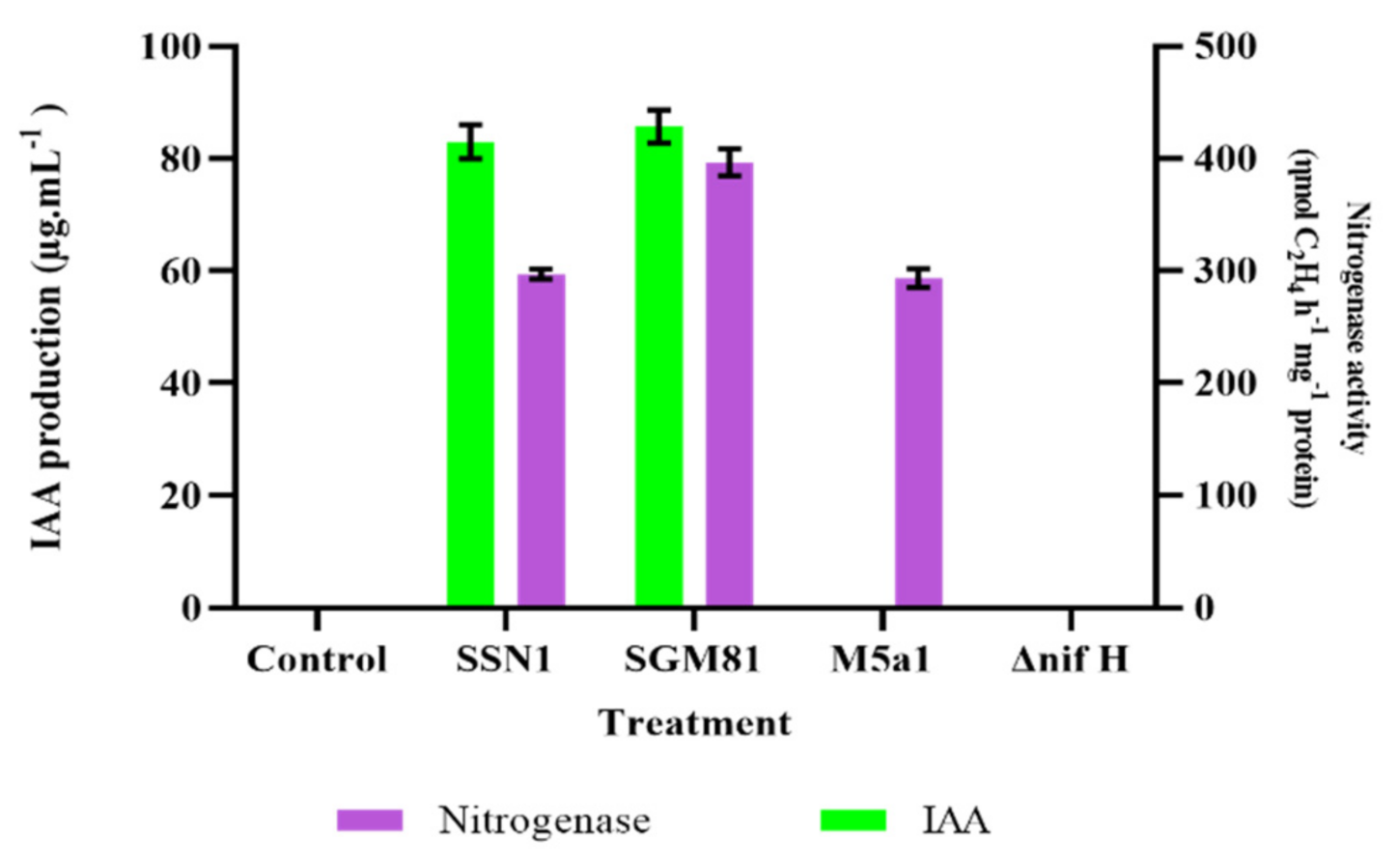
2.1. Characterization for Biological Nitrogen Fixation and Phytohormone Production in Klebsiella Strains
2.2. IAA Biosynthesis in Klebsiella Contributes to Enhanced Root Traits
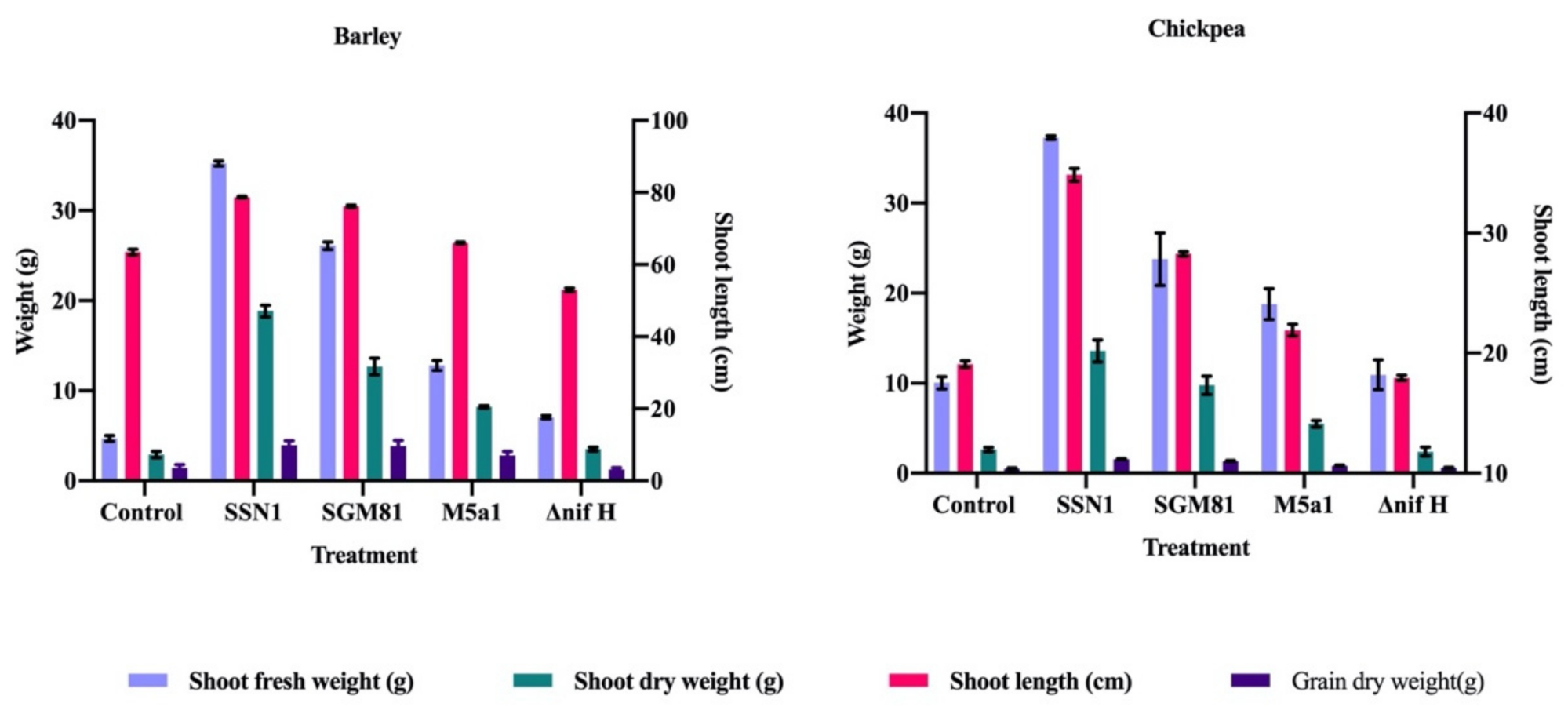
2.3. Klebsiella Improves Shoot Phenotype and Biochemical Profiling
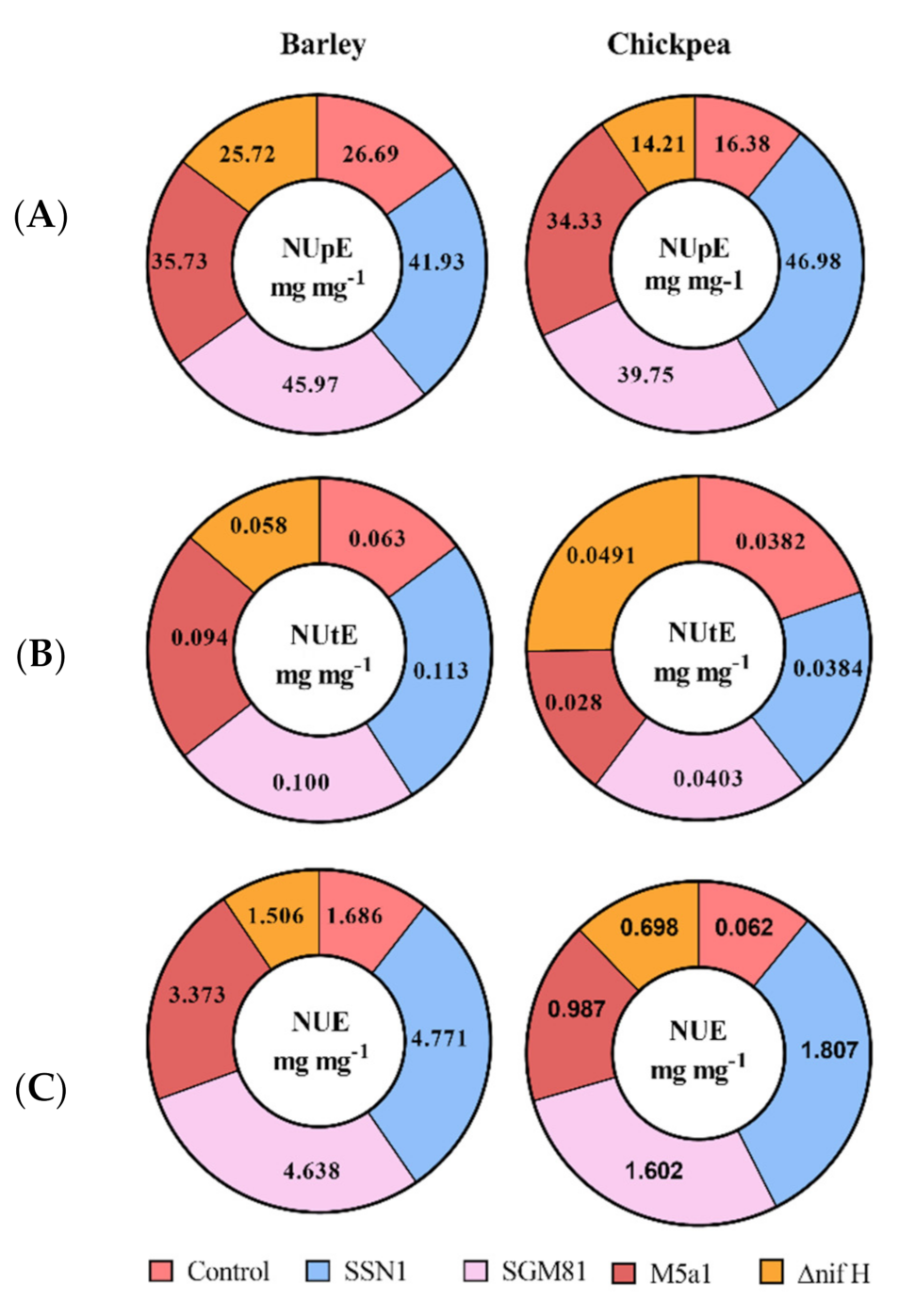
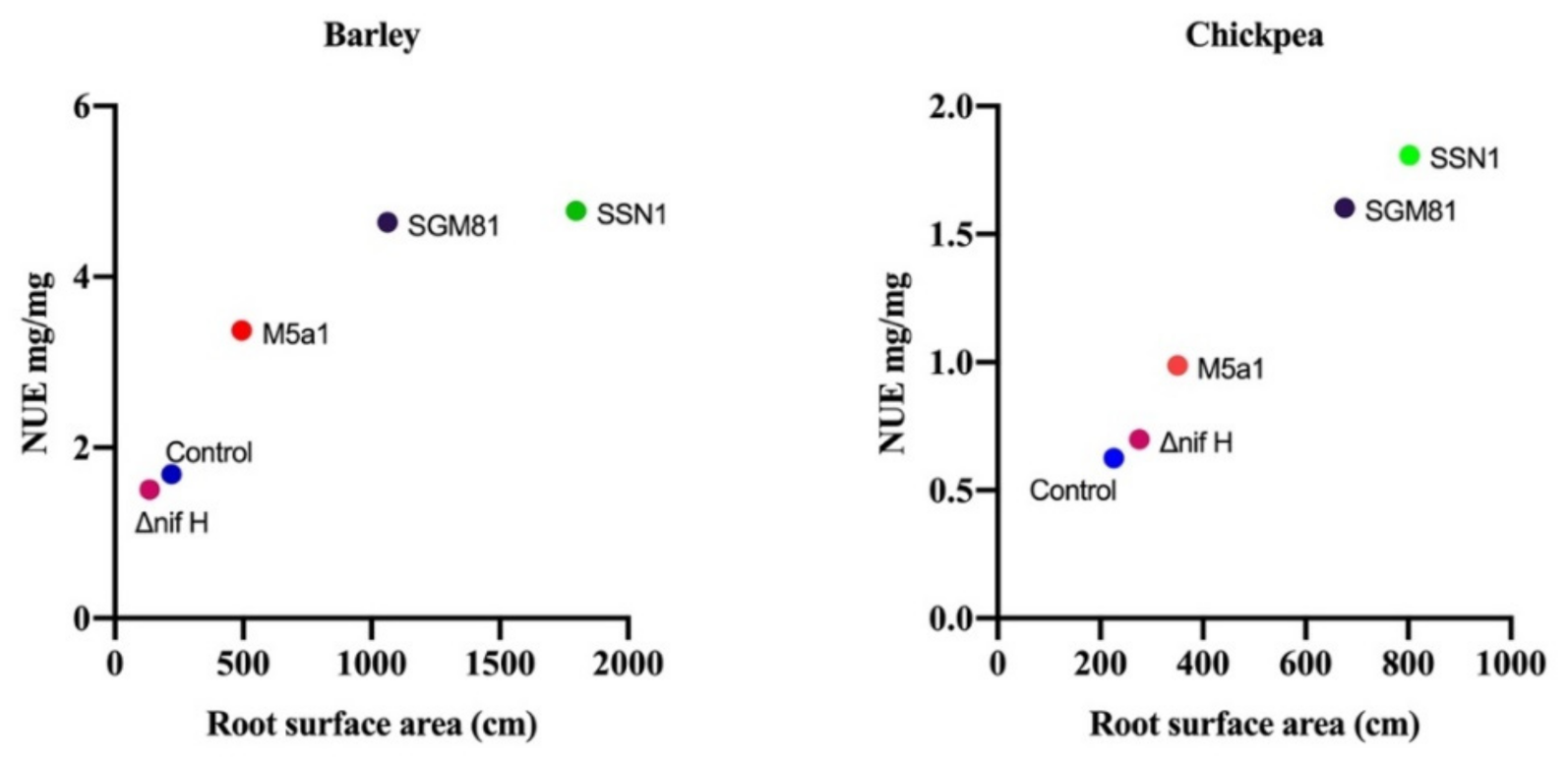
2.4. Klebsiella Demonstrates Effective Mobilization and Use of Nitrogen in Barley and Chickpea
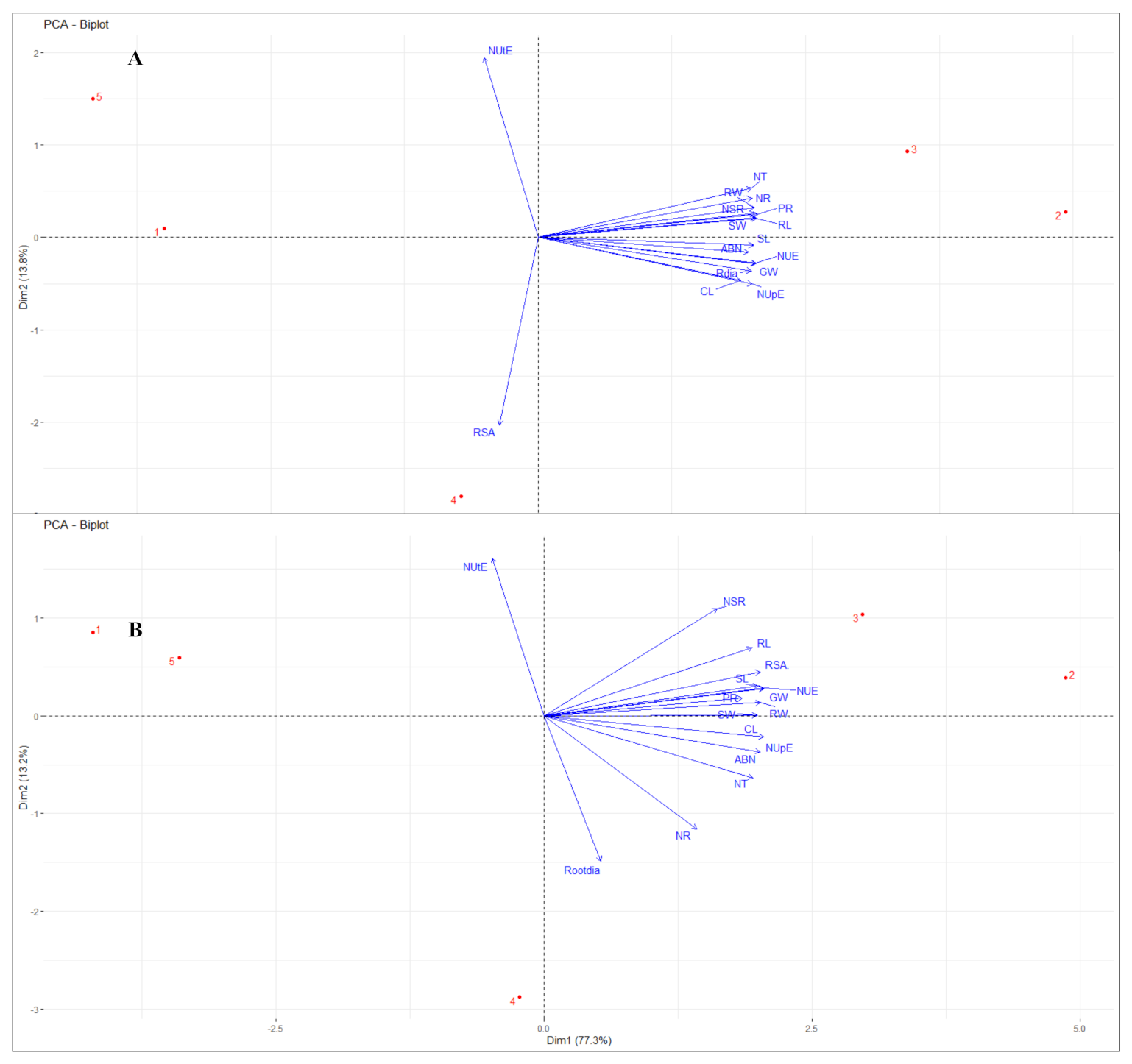
2.5. Principal Component Analysis
3. Discussion
4. Materials and Methods
4.1. Bacteria and Culture Conditions
4.2. Recombineering M5a1ΔnifH
4.3. Acetylene Reduction Assay
- C = concentration of ethylene in ηmoles.
- P = protein concentration of bacterial cell in mg.
- PS = peak height of sample.
- V = volume of air space in the assay vial.
- PStd = peak height of standard.
- T = time of incubation in h.
4.4. IAA Extraction and Determination by Salkowski Reagent Method
4.5. Field Experimental Design
4.5.1. Site Description
4.5.2. Plant Treatment and Growth
4.6. Plant Growth Parameters
- Root morphology traits comprising RL, RW, RDW, and NSR. These root traits were measured after uprooting the treated and control plants at the harvest stage after 90 days. Pictures of the roots were taken and processed in Image J for estimating the root diameter. In addition, RSA was calculated using the formula RSA = root length × root diameter × Π.
- Shoot morphology traits comprising of SL, SW, SDW, and GW per plant.
- Biochemical traits:
4.6.1. Chlorophyll Content
- Cl a (mg mL−1) = 12.7 A663–2.69 A645.
- Cl b (mg mL−1) = 22.9 A645–4.68 A663.
- A645 = absorbance at a wavelength of 645 nm.
- A663 = absorbance at a wavelength of 663 nm.
4.6.2. Total Protein
4.6.3. Plant Nitrogen Concentration
4.6.4. Nitrate and Nitrate Reductase Activity
4.7. Nitrogen Use Efficiency and Its Components
- NUpE: ABN/Ns.
- NUtE: Gw/ABN.
- NUE: Gw/Ns.
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hartman, K.; Tringe, S.G. Interactions between Plants and Soil Shaping the Root Microbiome under Abiotic Stress. Biochem. J. 2019, 476, 2705–2724. [Google Scholar] [CrossRef] [Green Version]
- FAO. Food and Agriculture Organization of the United Nations World Fertilizer Trends and Outlook to 2022; FAO: Rome, Italy, 2019. [Google Scholar]
- Andrews, M.; Lea, P. Our Nitrogen ‘Footprint’: The Need for Increased Crop Nitrogen Use Efficiency. Ann. Appl. Biol. 2013, 163, 165–169. [Google Scholar] [CrossRef]
- He, H.; Yang, R.; Li, Y.; Ma, A.; Cao, L.; Wu, X.; Chen, B.; Tian, H.; Gao, Y. Genotypic Variation in Nitrogen Utilization Efficiency of Oilseed Rape (Brassica Napus) under Contrasting N Supply in Pot and Field Experiments. Front. Plant Sci. 2017, 8, 1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakeem, K.R.; Mir, B.A.; Qureshi, M.I.; Ahmad, A.; Iqbal, M. Physiological Studies and Proteomic Analysis for Differentially Expressed Proteins and Their Possible Role in the Root of N-Efficient Rice (Oryza Sativa L.). Mol. Breed. 2013, 32, 785–798. [Google Scholar] [CrossRef] [Green Version]
- Kant, S.; Bi, Y.-M.; Rothstein, S.J. Understanding Plant Response to Nitrogen Limitation for the Improvement of Crop Nitrogen Use Efficiency. J. Exp. Bot. 2011, 62, 1499–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Benedetto, N.A.; Corbo, M.R.; Campaniello, D.; Cataldi, M.P.; Bevilacqua, A.; Sinigaglia, M.; Flagella, Z. The Role of Plant Growth Promoting Bacteria in Improving Nitrogen Use Efficiency for Sustainable Crop Production: A Focus on Wheat. AIMS Microbiol. 2017, 3, 413. [Google Scholar] [CrossRef]
- Moubayidin, L.; Di Mambro, R.; Sabatini, S. Cytokinin–Auxin Crosstalk. Trends Plant Sci. 2009, 14, 557–562. [Google Scholar] [CrossRef]
- Gang, S.; Saraf, M.; Waite, C.J.; Buck, M.; Schumacher, J. Mutualism between Klebsiella SGM 81 and Dianthus Caryophyllus in Modulating Root Plasticity and Rhizospheric Bacterial Density. Plant Soil 2017. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, H.-J.; George, E. Reduced15N-Nitrogen Transport through Arbuscular Mycorrhizal Hyphae to Triticum Aestivum L. Supplied with Ammonium vs. Nitrate Nutrition. Ann. Bot. 2001, 87, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Rosenblueth, M.; Ormeño-Orrillo, E.; López-López, A.; Rogel, M.A.; Reyes-Hernández, B.J.; Martínez-Romero, J.C.; Reddy, P.M.; Martínez-Romero, E. Nitrogen Fixation in Cereals. Front. Microbiol. 2018, 9, 1794. [Google Scholar] [CrossRef] [Green Version]
- Van Deynze, A.; Zamora, P.; Delaux, P.-M.; Heitmann, C.; Jayaraman, D.; Rajasekar, S.; Graham, D.; Maeda, J.; Gibson, D.; Schwartz, K.D. Nitrogen Fixation in a Landrace of Maize Is Supported by a Mucilage-Associated Diazotrophic Microbiota. PLoS Biol. 2018, 16, e2006352. [Google Scholar] [CrossRef] [Green Version]
- Kifle, M.H.; Laing, M.D. Isolation and Screening of Bacteria for Their Diazotrophic Potential and Their Influence on Growth Promotion of Maize Seedlings in Greenhouses. Front. Plant Sci. 2016, 6, 1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cakmakci, M.L.; Evans, H.; Seidler, R. Characteristics of Nitrogen-FixingKlebsiella Oxytoca Isolated from Wheat Roots. Plant Soil 1981, 61, 53–63. [Google Scholar] [CrossRef]
- Iniguez, A.L.; Dong, Y.; Triplett, E.W. Nitrogen Fixation in Wheat Provided by Klebsiella Pneumoniae 342. Mol. Plant-Microbe Interact. 2004, 17, 1078–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Li, Z.; Hu, C.; Zhang, X.; Chang, S.; Yang, L.; Li, Y.; An, Q. Plant Growth-Promoting Nitrogen-Fixing Enterobacteria Are in Association with Sugarcane Plants Growing in Guangxi, China. Microbes Environ. 2009, 27, 391–398. [Google Scholar] [CrossRef] [Green Version]
- Dixon, R.; Kahn, D. Genetic Regulation of Biological Nitrogen Fixation. Nat. Rev. Microbiol. 2004, 2, 621. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, M.; Vande Broek, A.; Dosselaere, F.; Vanderleyden, J. The IpdC Promoter Auxin-responsive Element of Azospirillum Brasilense, a Prokaryotic Ancestral Form of the Plant AuxRE? Mol. Microbiol. 1999, 32, 889–891. [Google Scholar] [CrossRef] [PubMed]
- Coulson, T.J.; Patten, C.L. The TyrR Transcription Factor Regulates the Divergent Akr-IpdC Operons of Enterobacter Cloacae UW5. PLoS ONE 2015, 10, e0121241. [Google Scholar] [CrossRef] [Green Version]
- Sachdev, D.P.; Chaudhari, H.G.; Kasture, V.M.; Dhavale, D.D.; Chopade, B.A. Isolation and Characterization of Indole Acetic Acid (IAA) Producing Klebsiella Pneumoniae Strains from Rhizosphere of Wheat (Triticum Aestivum) and Their Effect on Plant Growth. Indian J. Exp. Biol. 2009, 47, 993–1000. [Google Scholar]
- Jasim, B.; Jimtha, C.J.; Jyothis, M.; Radhakrishnan, E. Plant Growth Promoting Potential of Endophytic Bacteria Isolated from Piper Nigrum. Plant Growth Regul. 2013, 71, 1–11. [Google Scholar] [CrossRef]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.-L.; Touraine, B.; Moënne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Plant Growth-Promoting Rhizobacteria and Root System Functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-Acetic Acid in Microbial and Microorganism-Plant Signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-Acetic Acid in Plant–Microbe Interactions. Antonie van Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Li, X.; Zeng, R.; Liao, H. Improving Crop Nutrient Efficiency through Root Architecture Modifications. J. Integr. Plant Biol. 2016, 58, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Dhungana, S.A.; Itoh, K. Effects of Co-Inoculation of Indole-3-Acetic Acid-Producing and-Degrading Bacterial Endophytes on Plant Growth. Horticulturae 2019, 5, 17. [Google Scholar] [CrossRef]
- Kim, M.; Oh, H.-S.; Park, S.-C.; Chun, J. Towards a Taxonomic Coherence between Average Nucleotide Identity and 16S RRNA Gene Sequence Similarity for Species Demarcation of Prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Walch-Liu, P.; Filleur, S.; Gan, Y.; Forde, B.G. Signaling Mechanisms Integrating Root and Shoot Responses to Changes in the Nitrogen Supply. Photosynth. Res. 2005, 83, 239–250. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Z.; Wang, J.; Wang, Z.; Zhou, J. Plant Growth-Promoting Rhizobacteria Improve Shoot Morphology and Photosynthesis in Dryland Spring Wheat. Biomed. Eng. Environ. Eng. 2014, 145, 343–350. [Google Scholar]
- Khan, N.; Bano, A.M.; Babar, A. Impacts of Plant Growth Promoters and Plant Growth Regulators on Rainfed Agriculture. PLoS ONE 2020, 15, e0231426. [Google Scholar] [CrossRef] [Green Version]
- Bashan, Y.; Levanony, H. Current Status of Azospirillum Inoculation Technology: Azospirillum as a Challenge for Agriculture. Can. J. Microbiol. 1990, 36, 591–608. [Google Scholar] [CrossRef] [Green Version]
- Calvo, P.; Zebelo, S.; McNear, D.; Kloepper, J.; Fadamiro, H. Plant Growth-Promoting Rhizobacteria Induce Changes in Arabidopsis Thaliana Gene Expression of Nitrate and Ammonium Uptake Genes. J. Plant Interact. 2019, 14, 224–231. [Google Scholar] [CrossRef]
- Forde, B.G. Nitrate Transporters in Plants: Structure, Function and Regulation. Biochim. Biophys. Acta BBA-Biomembr. 2000, 1465, 219–235. [Google Scholar] [CrossRef]
- Raghuveer Rao, P.; Neeraja, C.; Vishnukiran, T.; Srikanth, B.; Vijayalakshmi, P.; Subhakara Rao, I.; Swamy, K.; Kondamudi, R.; Sailaja, N.; Subrahmanyam, D. Nitrogen Use Efficiency Ciency in Irrigated Rice for Climate Change—A Case Study; Directorate of Rice Research: Hyderabad, India, 2014. [Google Scholar]
- Perchlik, M.; Tegeder, M. Improving Plant Nitrogen Use Efficiency through Alteration of Amino Acid Transport Processes. Plant Physiol. 2017, 175, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Win, K.T.; Okazaki, K.; Ookawa, T.; Yokoyama, T.; Ohwaki, Y. Influence of Rice-Husk Biochar and Bacillus Pumilus Strain TUAT-1 on Yield, Biomass Production, and Nutrient Uptake in Two Forage Rice Genotypes. PLoS ONE 2019, 14, e0220236. [Google Scholar] [CrossRef] [Green Version]
- Ladha, J.; Pathak, H.J.; Krupnik, T.; Six, J.; van Kessel, C. Efficiency of Fertilizer Nitrogen in Cereal Production: Retrospects and Prospects. Adv Agron. 2005, 87, 85–156. [Google Scholar]
- Akter, Z.; Lupwayi, N.Z.; Balasubramanian, P. Nitrogen Use Efficiency of Irrigated Dry Bean (Phaseolus Vulgaris L.) Genotypes in Southern Alberta. Can. J. Plant Sci. 2017, 97, 610–619. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.A.; Kamiya, T.; Burritt, D.; Tran, L.-S.P.; Fujiwara, T. Plant Macronutrient Use Efficiency: Molecular and Genomic Perspectives in Crop Plants; Academic Press: Cambridge, MA, USA, 2017; ISBN 0-12-811294-8. [Google Scholar]
- Worku, M.; Bänziger, M.; Erley, G.S.A.; Friesen, D.; Diallo, A.O.; Horst, W.J. Nitrogen Uptake and Utilization in Contrasting Nitrogen Efficient Tropical Maize Hybrids. Crop Sci. 2007, 47, 519–528. [Google Scholar] [CrossRef] [Green Version]
- Rotundo, J.L.; Borrás, L.; De Bruin, J.; Pedersen, P. Soybean Nitrogen Uptake and Utilization in Argentina and United States Cultivars. Crop Sci. 2014, 54, 1153–1165. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-Step Inactivation of Chromosomal Genes in Escherichia Coli K-12 Using PCR Products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zumaquero, A.; Macho, A.P.; Rufián, J.S.; Beuzón, C.R. Analysis of the Role of the Type III Effector Inventory of Pseudomonas Syringae Pv. Phaseolicola 1448a in Interaction with the Plant. J. Bacteriol. 2010, 192, 4474–4488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushal, M.; Kaushal, R. Acetylene Reductase Activity and Molecular Characterization of Plant Growth Promoting Rhizobacteria to Know Efficacy in Integrated Nutrient Management System. Indian J. Biotechnol. 2015, 14, 221–227. [Google Scholar]
- He, F. Bradford Protein Assay. Bio-Protocol 2011, 101, e45. [Google Scholar] [CrossRef]
- Gang, S.; Sharma, S.; Saraf, M.; Buck, M.; Schumacher, J. Analysis of Indole-3-Acetic Acid (IAA) Production in Klebsiella by LC-MS/MS and the Salkowski Method. Bio-Protocol 2019, 9, e3230. [Google Scholar] [CrossRef]
- Staff, S. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014. [Google Scholar]
- Bent, A. Arabidopsis Thaliana Floral Dip Transformation Method. Agrobacterium Protoc. 2006, 87–104. [Google Scholar]
- Arnon, D.I. Copper Enzymes in Isolated Chloroplasts. Polyphenoloxidase in Beta Vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- AppliChem, P. Nitrogen Determination by Kjeldahl Method; PanReac AppliChem: Darmstadt, Germany, 2017. [Google Scholar]
- Zhao, L.; Wang, Y. Nitrate Assay for Plant Tissues. Biochem. Biophys. Res. Commun. 2017, 7, e2029. [Google Scholar] [CrossRef]
- Silveira, J.; Matos, J.; Cecatto, V.; Viegas, R.; Oliveira, J. Nitrate Reductase Activity, Distribution, and Response to Nitrate in Two Contrasting Phaseolus Species Inoculated with Rhizobium Spp. Environ. Exp. Bot. 2001, 46, 37–46. [Google Scholar] [CrossRef]
- Moll, R.; Kamprath, E.; Jackson, W. Analysis and Interpretation of Factors Which Contribute to Efficiency of Nitrogen Utilization 1. Agron. J. 1982, 74, 562–564. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants | Traits | Treatment | ||||
|---|---|---|---|---|---|---|
| Barley | Control | K.p. SSN1 | K.q. SGM81 | K.o. M5a1 | Δnif H | |
| Root Fresh weight (g) | 1.5 ± 0.039 *** | 6.58 ± 0.0084 *** | 5.117 ± 0.060 *** | 1.84 ± 0.0093 *** | 1.205 ± 0.085 *** | |
| Root dry weight (g) | 0.8 ± 0.050 *** | 4.217 ± 0.1249 *** | 3.4 ± 0.057 ** | 0.9200 ± 0.01125 *** | 0.300 ± 0.0057 *** | |
| Root length (cm) | 8.00 ± 0.392 *** | 22.12 ± 0.1195 *** | 18.08 ± 0.149 *** | 10.00 ± 0.1493 *** | 6.00 ± 0.0077 *** | |
| Root diameter (mm) | 9.583 ± 0.1417 * | 25.75 ±.6180 *** | 18.42 ± 0.3780 *** | 15.94 ± 0.5707 *** | 7.304 ± 0.3696 * | |
| Root surface area(cm) | 220.7 ± 16.02 ** | 1796 ± 31.35 *** | 1062.2 ± 22.90 *** | 494.5 ± 29.47 *** | 136.2 ± 88.39 ns | |
| Number of secondary roots | 10.33 ± 0.88 ** | 18.67 ± 1.202 ** | 22.00 ± 0.57 ** | 11.00 ± 0.5774 ns | 8.33±0.33 ns | |
| Chickpea | Root Fresh weight (g) | 1.117 ± 0.060 ** | 3.283 ± 0.070 *** | 2.5 ± 0.03 ** | 1.735 ± 0.04 * | 0.89 ± 0.008 * |
| Root dry weight (g) | 0.42 ± 0.012 ** | 1.78 ±0.018 *** | 1.43 ± 0.0095 *** | 0.52 ± 0.0079 *** | 0.266 ± 0.006 *** | |
| Root length (cm) | 7.117 ± 0.060 *** | 19.82 ± 0.1302 *** | 18.68 ± 0.364 *** | 8.017 ± 0.1108 ** | 7.033 ± 0.066 ns | |
| Root diameter (mm) | 10.26 ± 0.765 * | 12.72 ± 0.459 ns | 11.18 ± 0.415 ns | 13.66 ± 0.6018 ** | 12.74 ± 0.4287 * | |
| Root surface area (cm) | 226.3 ± 15.41 ** | 802.7 ± 26.66 *** | 676.2 ± 13.93 *** | 350.4 ± 16.50 ** | 276.0 ± 10.55 ns | |
| Number of secondary roots | 3.33 ± 0.88 ns | 5.00 ± 0.5 ns | 5.33 ± 0.3 ns | 3.33 ± 0.3 ns | 4.00 ± 0.57 ns | |
| Plants | Traits | Treatment | ||||
|---|---|---|---|---|---|---|
| Barley | Control | K.p. SSN1 | K.q. SGM81 | K.o. M5a1 | Δnif H | |
| Chlorophyll (mg mL−1) | 1.079 ± 0.0547 * | 1.530 ± 0.1281 ** | 1.372 ± 0.110 * | 1.362 ± 0.0614 * | 1.165 ± 0.04829 * | |
| Protein (mg g−1 FW h−1) | 1.200 ± 0.0774 * | 2.130 ± 0.1134 ** | 2.042 ± 0.1428 ** | 1.395 ± 0.1919 ** | 1.133 ± 0.1436 ns | |
| Nitrate reductase (µg g−1 FW h−1) | 10.22 ± 0.3156 ** | 14.20 ± 0.5447 *** | 14.60 ± 0.2781 *** | 10.93 ± 0.6344 * | 10.12 ± 0.4257* | |
| Nitrate (µg g−1) | 381.2 ± 450.0** | 604.2 ± 37.93*** | 593.5 ± 33.25*** | 424.2 ± 12.60ns | 395.1 ± 22.26 ns | |
| Total nitrogen (mg g−1) | 22.16 ± 2.482* | 34.81 ± 3.875** | 38.16 ± 1.447** | 29.66 ± 4.425* | 21.35 ± 1.384 ns | |
| Chickpea | Chlorophyll (mg mL−1) | 0.817 ±0.07417 *** | 1.685 ± 0045 ** | 1.450 ± 0.065 ** | 1.222 ± 0.1756 ** | 0.8275 ±0.060 * |
| Protein (mg g−1 FW h−1) | 1.295 ± 0.1350 * | 1.760 ± 0.1395 * | 1.967 ± 0.0928 ** | 1.577 ± 0.0954 ns | 1.433 ± 0.0954 ns | |
| Nitrate reductase (µg g−1 FW h−1) | 10.55 ± 0.4796 ** | 15.53 ± 1.322 *** | 16.57 ± 0.9735 *** | 17.35 ± 0.6020 ns | 13.67 ± 0.2257 ns | |
| Nitrate (µg g−1) | 353.1 ± 17.83 ** | 533.0 ± 34.62 ** | 484.2 ± 7.955 ** | 484.9 ± 17.21 ns | 397.3 ± 6.172 ns | |
| Total nitrogen (mg g−1) | 13.65 ± 2.724 ** | 39.00 ± 1.483 *** | 33.00 ± 3.183 *** | 28.50 ± 1.310 ** | 11.83 ±5.084 ns | |
| Trait | Barley | Chickpea | ||
|---|---|---|---|---|
| Dim.1 | Dim.2 | Dim.1 | Dim.2 | |
| Root fresh weight (RFW) | 0.9801 | −0.1458 | 0.9817 | 0.0430 |
| Root length (RL) | 0.9897 | −0.0874 | 0.9480 | 0.3087 |
| Shoot length (SL) | 0.9713 | 0.0506 | 0.9721 | 0.1266 |
| Total nitrogen (Nt) | 0.9438 | 0.0923 | 0.9705 | −0.2037 |
| Chlorophyll (Cl) | 0.9130 | 0.2394 | 0.9893 | −0.1326 |
| Nitrate reductase (NR) | 0.9650 | −0.1917 | 0.6626 | −0.5968 |
| Nitrate (NT) | 0.9664 | −0.2451 | 0.9306 | −0.3403 |
| Protein (PR) | 0.9918 | −0.1094 | 0.8870 | 0.0474 |
| Number of secondary roots (NSR) | 0.9806 | −0.1137 | 0.7895 | 0.4932 |
| Nitrogen use efficiency (NUE) | 0.9776 | 0.1497 | 0.9938 | 0.1065 |
| Nitrogen uptake efficiency (NUpE) | 0.9632 | 0.2575 | 0.9705 | −0.2037 |
| Nitrogen utilization efficiency (NUtE) | −0.2301 | −0.9407 | −0.2182 | 0.7675 |
| Grain weight (GW) | 0.9774 | 0.1509 | 0.9945 | 0.1042 |
| Root diameter (Rdia) | 0.9629 | 0.1877 | 0.2290 | −0.7412 |
| Root surface area (RSA) | −0.1874 | 0.9773 | 0.9815 | 0.1875 |
| Root dry weight (RDW) | 0.9760 | −0.1830 | 0.9616 | 0.2426 |
| Shoot dry weight (SDW) | 0.9821 | 0.0292 | 0.9883 | 0.0800 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gang, S.; Sharma, S.; Saraf, M.; Buck, M.; Schumacher, J. Bacterial Indole-3-Acetic Acid Influences Soil Nitrogen Acquisition in Barley and Chickpea. Plants 2021, 10, 780. https://doi.org/10.3390/plants10040780
Gang S, Sharma S, Saraf M, Buck M, Schumacher J. Bacterial Indole-3-Acetic Acid Influences Soil Nitrogen Acquisition in Barley and Chickpea. Plants. 2021; 10(4):780. https://doi.org/10.3390/plants10040780
Chicago/Turabian StyleGang, Shraddha, Sheetal Sharma, Meenu Saraf, Martin Buck, and Jorg Schumacher. 2021. "Bacterial Indole-3-Acetic Acid Influences Soil Nitrogen Acquisition in Barley and Chickpea" Plants 10, no. 4: 780. https://doi.org/10.3390/plants10040780
APA StyleGang, S., Sharma, S., Saraf, M., Buck, M., & Schumacher, J. (2021). Bacterial Indole-3-Acetic Acid Influences Soil Nitrogen Acquisition in Barley and Chickpea. Plants, 10(4), 780. https://doi.org/10.3390/plants10040780