
Microalgal Cell Biofactory—Therapeutic, Nutraceutical and Functional Food Applications


Abstract
:1. Introduction
2. Potential of Microalgae
3. Bioactive Compounds
3.1. Chlorophyll
3.2. Carotenoids
3.3. Lipids
3.4. Carbohydrates
3.5. Proteins
3.6. Vitamins
3.7. Sterols
4. Therapeutical Function of Microalgae
4.1. Antioxidant Activity
4.2. Antimicrobial Activity
4.2.1. Antibacterial Activity
4.2.2. Antiviral Activity
4.2.3. Antifungal Activity
4.3. Anticancer Activity
4.4. Anti-Inflammatory
4.5. Antiaging
4.6. Other Activities
5. Functional Food/Feed
5.1. Functional Food
5.2. Feed—Animal and Aquaculture
6. Microalgae Cultivation
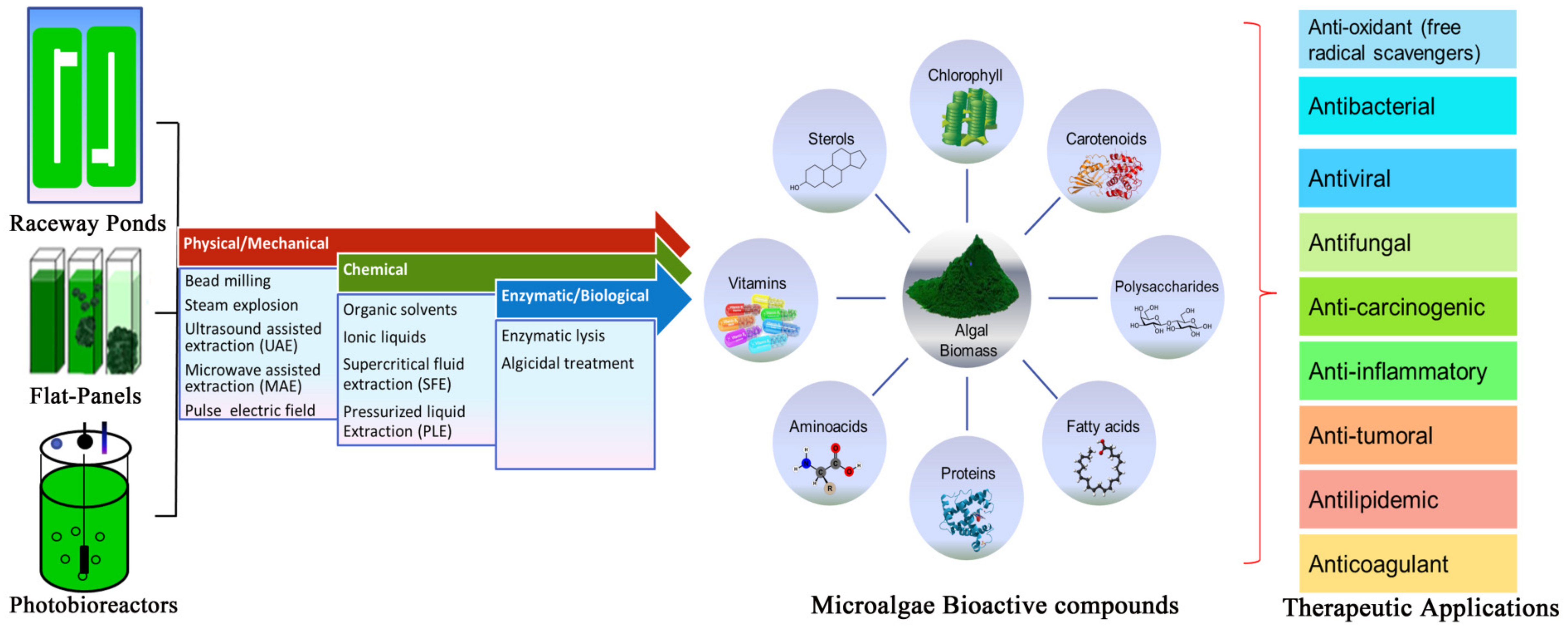
7. Harvesting and Downstream Process of Microalgae
7.1. Mechanical/Physical Pre-Treatment
7.2. Chemical Pre-Treatment
7.3. Enzymatic/Biological Pre-Treatment
8. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- United Nations Department of Economic and Social Affairs (UNDES). World Population Projected to Reach 9.8 Billion in 2050, and 11.2 Billion in 2100. 2017. Available online: https://www.un.org/development/desa/en/news/population/world-population-prospects-2017.html (accessed on 19 March 2018).
- Sreeharsha, R.V.; Venkata Mohan, S.V. Symbiotic integration of bioprocesses to design a self-sustainable life supporting ecosystem in a circular economy framework. Bioresour. Technol 2021, 124712. [Google Scholar] [CrossRef]
- FAO. World Agriculture: Towards 2015/2030, Summary Report (2002). Available online: http://www.fao.org/docrep/004/Y3557E/y3557e03.htm (accessed on 19 March 2018).
- FAO. Protein Sources for the Animal Feed Industry. 2004. Available online: http://www.fao.org/docrep/007/y5019e/y5019e03.htm (accessed on 23 March 2018).
- Sudhakar, M.P.; Kumar, B.R.; Mathimani, T.; Arunkumar, K. A review on bioenergy and bioactive compounds from microalgae and macroalgae-sustainable energy perspective. J. Clean. Prod. 2019, 228, 1320–1333. [Google Scholar] [CrossRef]
- Sosa-Hernández, J.E.; Escobedo-Avellaneda, Z.; Iqbal, H.; Welti-Chanes, J. State-of-the-art extraction methodologies for bioactive compounds from algal biome to meet bio-economy challenges and opportunities. Molecules 2018, 23, 2953. [Google Scholar] [CrossRef] [Green Version]
- Torres-Tiji, Y.; Fields, F.J.; Mayfield, S.P. Microalgae as a future food source. Biotechnol. Adv. 2020, 107536. [Google Scholar] [CrossRef] [PubMed]
- Venkata Mohan, S.; Hemalatha, M.; Chakraborty, D.; Chatterjee, S.; Ranadheer, P.; Kona, R. Algal biorefinery models with self-sustainable closed loop approach. Trends and prospective for blue-bioeconomy. Bioresour. Technol. 2020, 122128. [Google Scholar] [CrossRef] [PubMed]
- Venkata Mohan, S.; Rohit, M.V.; Chiranjeevi, P.; Hariprasad, R.; Arora, S. Integrating microalgae cultivation with wastewater treatment for biodiesel production. In Algal Biorefinery: An Integrated Approach; Springer: Cham, Switzerland, 2015; pp. 321–337. [Google Scholar]
- dos Santos Alves Figueiredo Brasil, B.; Gonçalves de Siqueira, F.; Chan Salum, T.F.; Zanette, C.M.; Spier, M.R. Microalgae and cyanobacteria as enzyme biofactories. Algal Res. 2017, 25, 76–89. [Google Scholar]
- Levasseur, W.; Perré, P.; Pozzobon, V. A review of high value-added molecules production by microalgae in light of the classification. Biotechnol. Adv. 2020, 107545. [Google Scholar] [CrossRef]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for high-value products towards human health and nutrition. Mar. Drugs. 2019, 17, 304. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Zhu, L.; Li, S.; Hu, T.; Chu, R.; Mo, F.; Hu, D.; Liu, C.; Li, B. A comprehensive review on cultivation and harvesting of microalgae for biodiesel production: Environmental pollution control and future directions. Bioresour. Technol. 2020, 301, 122804. [Google Scholar] [CrossRef]
- Koyande, A.K.; Chew, K.W.; Rambabu, K.; Tao, Y.; Chu, D.T.; Show, P.L. Microalgae: A potential alternative to health supplementation for humans. Food Sci. Hum. Wellness 2019, 8, 16–24. [Google Scholar] [CrossRef]
- Globe Newswire. Herbal Nutraceutical Supplements in Vogue as Consumer Preference for Natural Soars. Available online: http://in.style.yahoo.com/herbal-nutraceutical-supplements-vogue-consumer-143000805.html (accessed on 18 December 2018).
- Williams, R. Microscopic Algae Produce Half the Oxygen we Breathe. ABC News. October 2013. Available online: http://www.abc.Net.au/radionational/programs/scienceshow/microscopic-algae-produce-half-the-oxygen-we-breathe/5041338. (accessed on 19 May 2019).
- Da Silva Vaz, B.; Moreira, J.B.; de Morais, M.G.; Costa, J.A.V. Microalgae as a new source of bioactive compounds in food supplements. Curr. Opin. Food Sci. 2016, 7, 73–77. [Google Scholar]
- Kılınç, B.; Cirik, S.; Turan, G.; Tekogul, H.; Koru, E. Seaweeds for food and industrial applications. InTechOpen 2013, 735–747. [Google Scholar] [CrossRef] [Green Version]
- Jacob-Lopes, E.; Maroneze, M.M.; Deprá, M.C.; Sartori, R.B.; Dias, R.R.; Zepka, L.Q. Bioactive food compounds from microalgae: An innovative framework on industrial biorefineries. Curr. Opin. Food Sci. 2019, 25, 1–7. [Google Scholar] [CrossRef]
- Wilson, G.M., Jr.; Gorgich, M.; Corrêa, P.S.; Martins, A.A.; Mata, T.M.; Caetano, N.S. Microalgae for biotechnological applications. Cultivation, harvesting and biomass processing. Aquaculture 2020, 735562. [Google Scholar] [CrossRef]
- Vuppaladadiyam, A.K.; Prinsen, P.; Raheem, A.; Luque, R.; Zhao, M. Microalgae cultivation and metabolites production: A comprehensive review. Biofuel Bioprod. Biorefin. 2018, 12, 304–324. [Google Scholar] [CrossRef]
- Tang, D.Y.Y.; Khoo, K.S.; Chew, K.W.; Tao, Y.; Ho, S.H.; Show, P.L. Potential utilization of bioproducts from microalgae for the quality enhancement of natural products. Bioresour. Technol. 2020, 122997. [Google Scholar] [CrossRef]
- Onofrejova, L.; Vasickova, J.; Klejdus, B.; Stratil, P.; Misurcova, L.; Kracmar, S.; Kopecky, J.; Vacek., J. Bioactive phenols in algae: The application of pressurized-liquid and solid-phase extraction techniques. J. Pharm. Biomed. Anal. 2010, 51, e464–e470. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, V.; Gunasegavan, R.D.; Mustar, S.; Lee, J.C.; Mohd Noh, M.F. Isolation of industrial important bioactive compounds from microalgae. Molecules 2021, 26, 943. [Google Scholar] [CrossRef]
- Rosas, J.G.; Gómez, N.; Cara-Jiménez, J.; González-Arias, J.; Olego, M.Á.; Sánchez, M.E. Evaluation of Joint Management of Pine Wood Waste and Residual Microalgae for Agricultural Application. Sustainability 2021, 13, 53. [Google Scholar] [CrossRef]
- Odjadjare, E.C.; Mutanda, T.; Olaniran, A.O. Potential biotechnological application of microalgae: A critical review. Crit. Rev. Biotechnol. 2017, 37, 37–52. [Google Scholar] [CrossRef]
- Mishra, V.K.; Bacheti, R.K.; Husen, A. Medicinal uses of chlorophyll: A critical overview. In Chlorophyll: Structure, Function and Medicinal; Le, H., Salcedo, E., Eds.; Nova Science Publishers: Hauppauge, NY, USA, 2011; pp. 177–196. [Google Scholar]
- Khanra, S.; Mondal, M.; Halderl, G.; Tiwari, O.N.; Gayen, K.; Bhowmick, T.K. Downstream processing of microalgae for pigments, protein and carbohydrate in industrial application: A review. Food Bioprod. Process 2018, 110, 60–84. [Google Scholar] [CrossRef]
- Rajesh, K.; Rohit, G.; Venkata Mohan, S. Microalgae-based carotenoids production. In Algal Green Chemistry; Rastogi, R.P., Madamwar, D., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 139–147. [Google Scholar]
- Galasso, C.; Gentile, A.; Orefice, I.; Ianora, A.; Bruno, A.; Noonan, D.M.; Sansone, C.; Albini, A.; Brunet, C. Microalgal derivatives as potential nutraceutical and food supplements for human health: A focus on cancer prevention and interception. Nutrients 2019, 11, 1226. [Google Scholar] [CrossRef] [Green Version]
- Gong, M.; Bassi, A. Carotenoids from microalgae: A review of recent developments. Biotechnol. Adv. 2016, 34, 1396–1412. [Google Scholar] [CrossRef]
- Capelli, R.; Talbott, S.; Ding, L. Astaxanthin sources: Suitability for human health and nutrition. Funct. Foods Health Dis. 2019, 9, 430–445. [Google Scholar] [CrossRef] [Green Version]
- Talukdar, J.; Dasgupta, S.; Nagle, V.; Bhadra, B. COVID-19: Potential of Microalgae Derived Natural Astaxanthin as Adjunctive Supplement in Alleviating Cytokine Storm. 2020. Available online: https://dx.doi.org/10.2139/ssrn.3579738 (accessed on 18 April 2020).
- Lin, J.H.; Lee, D.J.; Chang, J.S. Lutein production from biomass: Marigold flowers versus microalgae. Bioresour. Technol. 2015, 184, 421–428. [Google Scholar] [CrossRef]
- Sanzo, G.D.; Mehariya, S.; Martino, M.; Larocca, V.; Casella, P.; Chianese, S.; Musmarra, D.; Balducchi, R.; Molino, A. Supercritical carbon dioxide extraction of astaxanthin, lutein, and fatty acids from Haematococcus pluvialis microalgae. Mar. Drugs 2018, 16, 334. [Google Scholar] [CrossRef] [Green Version]
- Abu-Ghosh, S.; Dubinsky, Z.; Verdelho, V.; Iluz, D. Unconventional high-value products from microalgae. A review. Bioresour. Technol. 2021, 124895. [Google Scholar] [CrossRef]
- Folarin, O.; Sharma, L. Algae as a functional food. Int. J. Home Sci. 2017, 3, 166–170. [Google Scholar]
- Katiyar, R.; Arora, A. Health promoting functional lipids from microalgae pool: A review. Algal Res. 2020, 46, e101800. [Google Scholar] [CrossRef]
- Sathasivam, R.; Radhakrishnan, R.; Hashem, A.; Abd-Allah, E.F. Microalgae metabolites: A rich source for food and medicine. Saudi J. Biol. Sci. 2019, 26, 709–722. [Google Scholar] [CrossRef]
- Rohit, M.V.; Venkata Mohan, S. Quantum Yield and Fatty Acid Profile Variations with Nutritional Mode during Microalgae Cultivation. Front. Bioeng. Biotechnol. 2018, 6, 111. [Google Scholar] [CrossRef]
- Charles, C.N.; Msagati, T.; Swai, H.; Chacha, M. Microalgae: An alternative natural source of bioavailable omega-3 DHA for promotion of mental health in East Africa. Sci. African. 2019, 6, e00187. [Google Scholar] [CrossRef]
- Bhalamurugan, G.L.; Valerie, O.; Mark, L.; Chacha, M. Valuable bioproducts obtained from microalgal biomass and their commercial applications: A review. Environ. Eng. Res. 2018, 23, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Chiranjeevi, P.; Venkata Mohan, S. Critical parametric influence on microalgae cultivation towards maximizing biomass growth with simultaneous lipid productivity. Renew. Energy 2016, 98, 64–71. [Google Scholar] [CrossRef]
- Cesário, M.T.; da Fonseca, M.M.R.; Marques, M.M.; de Almeida, M.C.M. Marine algal carbohydrates as carbon sources for the production of biochemicals and biomaterials. Biotechnol. Adv. 2018, 36, 798–817. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Huang, G. The antiviral activity of polysaccharides and their derivatives. Int. J. Biol. Macromol. 2018, 115, 77–82. [Google Scholar] [CrossRef] [PubMed]
- De Morais, M.G.; Vaz, B.D.S.; de Morais, E.G.; Costa, J.A.V. Biologically active metabolites synthesized by microalgae. BioMed Res. Int. 2015, 835761. [Google Scholar] [CrossRef] [Green Version]
- Gardeva, E.; Toshkova, R.; Minkova, K.; Gigova, L. Cancer protective action of polysaccharide, derived from red microalga Porphyridium cruentum—A biological background. Biotechnol. Biotechnol. Equip. 2009, 23, 783–787. [Google Scholar] [CrossRef]
- Lekshmi, V.S.; Kurup, G.M. Anticoagulant activities of sulfated polysaccharides from the edible marine algae Padina tetrastromatica and Ulva fasciata: A combined in vitro and in vivo approach. J. Pharmacogn. Phytochem. 2019, 8, 693–698. [Google Scholar]
- Gaikwad, M.; Pawar, Y.; Nagle, V.; Dasgupta, S. Marine red alga Porphyridium sp. as a source of sulfated polysaccharides (SPs) for combating against COVID-19. Preprints 2020, 2020040168, in press. [Google Scholar]
- Caporgno, M.P.; Mathys, A. Trends in microalgae incorporation into innovative food products with potential health benefits. Front. Nutr. 2018, 5, 58. [Google Scholar] [CrossRef]
- Hempel, F.; Lau, J.; Klingl, A.; Maier, U.G. Algae as protein factories: Expression of a human antibody and the respective antigen in the diatom Phaeodactylum tricornutum. PLoS ONE 2011, 6, e28424. [Google Scholar]
- Paliwal, C.; Mitra, M.; Bhayani, K.; Bharadwaj, S.V.; Ghosh, T.; Dubey, S.; Mishra, S. Abiotic stresses as tools for metabolites in microalgae. Bioresour. Technol. 2017, 244, 1216–1226. [Google Scholar] [CrossRef]
- Hempel, F.; Maier, U.G. Microalgae as solar-powered protein factories. In Advanced Technologies for Protein Complex Production and Characterization; Springer: Cham, Switzerland, 2016; pp. 241–262. [Google Scholar]
- Gregory, J.A.; Topol, A.B.; Doerner, D.Z.; Mayfield, S. Alga-produced cholera toxin-Pfs25 fusion proteins as oral vaccines. Appl. Environ. Microbiol. 2013, 79, 3917–3925. [Google Scholar] [CrossRef] [Green Version]
- Specht, E.; Miyake-Stoner, S.; Mayfield, S. Micro-algae come of age as a platform for recombinant protein production. Biotechnol. Lett. 2010, 32, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Andrade, L.M.; Andrade, C.J.; Dias, M.; Nascimento, C.A.O.; Mendes, M.A. Chlorella and Spirulina Microalgae as Sources of Functional Foods, Nutraceuticals, and Food Supplements; an overview. MOJ Food Process Technol. 2018, 6, 45–58. [Google Scholar]
- Marzieh Hosseini, S.; Shahbazizadeh, S.; Khosravi-Darani, K.; Reza Mozafari, M. Spirulina paltensis: Food and function. Curr. Nutr. Food Sci. 2013, 9, 189–193. [Google Scholar]
- Tarento, T.D.; McClure, D.D.; Vasiljevski, E.; Schindeler, A.; Dehghani, F.; Kavanagh, J.M. Microalgae as a source of vitamin K1. Algal Res. 2018, 36, 77–87. [Google Scholar]
- Giammanco, M.; Di Majo, D.; La Guardia, M.; Aiello, S.; Crescimannno, M.; Flandina, C.; Tumminello, F.M.; Leto, G. Vitamin D in cancer chemoprevention. Pharm. Biol. 2015, 53, 1399–1434. [Google Scholar]
- Bong, S.C.; Loh, S.P. A study of fatty acid composition and tocopherol content of lipid extracted from marine microalgae, Nannochloropsis oculata and Tetraselmis suecica, using solvent extraction and supercritical fluid extraction. Int. Food Res. J. 2013, 20, 721–729. [Google Scholar]
- Lopes, G.; Sousa, C.; Valentao, P.; Andrade, P.B. Sterols in algae and health. In Bioactive Compounds from Marine Foods: Plant and Animal Sources; Hernandez-Ledesma, B., Herrero, M., Eds.; John Wiley & Sons: Hoboken, New Jersey, USA, 2013; pp. 173–191. ISBN 9781118412893. [Google Scholar]
- Luo, X.; Su, P.; Zhang, W. Advances in microalgae-derived phytosterols for functional food and pharmaceutical applications. Mar. Drugs 2015, 13, 4231–4254. [Google Scholar]
- Ahmed, F.; Zhou, W.; Schenk, P.M. Pavlova lutheri is a high-level producer of phytosterols. Algal Res. 2015, 10, 210–217. [Google Scholar]
- Koller, M.; Muhr, A.; Braunegg, G. Microalgae as versatile cellular factories for valued products. Algal Res. 2014, 6, 52–63. [Google Scholar] [CrossRef]
- Cai, X.; Chen, Y.; Xie, X.; Yao, D.; Ding, C.; Chen, M. Astaxanthin prevents against lipopolysaccharide-induced acute lung injury and sepsis via inhibiting activation of MAPK/NF-κB. Am. J. Transl. Res. 2019, 11, 1884. [Google Scholar]
- Chang, C.H.; Chen, C.Y.; Chiou, J.Y.; Peng, R.Y.; Peng, C.H. Astaxanthine secured apoptotic death of PC12 cells induced by β-amyloid peptide 25–35: Its molecular action targets. J. Med. Food 2010, 13, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Chyun, J.H.; Kim, Y.K.; Line, L.L.; Chew, B.P. Astaxanthin decreased oxidative stress and inflammation and enhanced immune response in humans. Nutr. Metab. 2010, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Zhang, X.S.; Wang, H.D.; Zhang, X.; Yu, Q.; Li, W.; Zhou, M.L.; Wang, X.L. Astaxanthin activates nuclear factor erythroid-related factor 2 and the antioxidant responsive element (Nrf2-ARE) pathway in the brain after subarachnoid hemorrhage in rats and attenuates early brain injury. Mar. Drugs 2014, 12, 6125–6141. [Google Scholar] [CrossRef]
- Liu, T.; Liu, W.H.; Zhao, J.S.; Meng, F.Z.; Wang, H. Lutein protects against β-amyloid peptide-induced oxidative stress in cerebrovascular endothelial cells through modulation of Nrf-2 and NF-κb. Cell Biol. Toxicol. 2017, 33, 57–67. [Google Scholar]
- Rasmussen, H.M.; Johnson, E.J. Nutrients for the aging eye. Clin. Interv. Aging 2013, 8, 741–748. [Google Scholar]
- Sansone, C.; Galasso, C.; Orefice, I.; Nuzzo, G.; Luongo, E.; Cutignano, A.; Romano, G.; Brunet, C.; Fontana, A.; Esposito, F.; et al. The green microalga Tetraselmis suecica reduces oxidative stress and induces repairing mechanisms in human cells. Sci. Rep. 2017, 7, 1–12. [Google Scholar]
- Morokutti-Kurz, M.; Graf, C.; Prieschl-Grassauer, E. Amylmetacresol/2, 4-dichlorobenzyl alcohol, hexylresorcinol, or carrageenan lozenges as active treatments for sore throat. Int. J. Gen. Med. 2017, 10, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Therapeutic potentials of microalgae in the treatment of Alzheimer’s disease. Molecules 2017, 22, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raposo, M.F.D.J.; Morais, A.M. Bioactivity and applications of polysaccharides from marine microalgae. Mar. Drugs. 2014, 11, 233–252. [Google Scholar]
- Long, S.F.; Kang, S.; Wang, Q.Q.; Xu, Y.T.; Pan, L.; Hu, J.X.; Li, M.; Piao, X.S. Dietary supplementation with DHA-rich microalgae improves performance, serum composition, carcass trait, antioxidant status, and fatty acid profile of broilers. Poult. Sci. 2018, 97, 1881–1890. [Google Scholar] [CrossRef]
- Raghukumar, S. Thraustochytrid marine protists: Production of PUFAs and other emerging technologies. Mar. Biotechnol. 2008, 10, 631–640. [Google Scholar] [CrossRef]
- Teng, E.; Taylor, K.; Bilousova, T.; Weiland, D.; Pham, T.; Zuo, X.; Yang, F.; Chen, P.P.; Glabe, C.G.; Takacs, A.; et al. Dietary DHA supplementation in an APP/PS1 transgenic rat model of AD reduces behavioral and Aβ pathology and modulates Aβ oligomerization. Neurobiol. Dis. 2015, 82, 552–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mourelle, M.L.; Gómez, C.P.; Legido, J.L. The potential use of marine microalgae and cyanobacteria in cosmetics and thalassotherapy. Cosmetics 2017, 4, 46. [Google Scholar] [CrossRef] [Green Version]
- Day, A.G.; Brinkmann, D.; Franklin, S.; Espina, K.; Rudenko, G.; Roberts, A.; Howse, K.S. Safety evaluation of a high-lipid algal biomass from Chlorella protothecoides. Regul. Toxicol. Pharmacol. 2009, 55, 166–180. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Yim, J.H.; Kim, S.Y.; Kim, H.S.; Lee, W.G.; Kim, S.J.; Kang, P.S.; Lee, C.K. In vitro inhibition of influenza A virus infection by marine microalga-derived sulfated polysaccharide p-KG03. Antivir. Res. 2012, 93, 253–259. [Google Scholar] [CrossRef]
- Márquez-Escobar, V.A.; Bañuelos-Hernández, B.; Rosales-Mendoza, S. Expression of a Zika virus antigen in microalgae: Towards mucosal vaccine development. J Biotechnol. 2018, 282, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, M. Pharmaceutically valuable bioactive compounds of algae. Asian J. Pharm. Clin. Res. 2016, 9, 43–47. [Google Scholar] [CrossRef]
- Kim, S.K.; Kang, K.H. Medicinal effects of peptides from marine microalgae. Adv. Food Nutr. Res. 2011, 64, 313–323. [Google Scholar]
- Lawrence, K.P.; Long, P.F.; Young, A.R. Mycosporine-like amino acids for skin photoprotection. Curr. Med. Chem. 2018, 25, 5512–5527. [Google Scholar]
- Becker, E. Microalgae for Human and Animal Nutrition. In Handbook of Microalgae Culture: Applied Phycology and Biotechnology, 2nd ed.; Richmond, A., Ed.; Blackwell Science: London, UK, 2004; pp. 461–503. ISBN 978-0-4709-9528-0. [Google Scholar]
- Desaloie, J.; Rossier, J.; Haeni, L.; Rothen-Rutishauser, B.; Zobi, F. Slow-targeted release of a ruthenium anticancer agent from vitamin B 12 functionalized marine diatom microalgae. Dalton Trans. 2018, 47, 17221–17232. [Google Scholar] [CrossRef]
- Kattappagari, K.K.; Teja, C.R.; Kommalapati, R.K.; Poosarla, C.; Gontu, S.R.; Reddyu, B.V.R. Role of antioxidants in facilitating the body functions: A review. J. Orofac. Sci. 2015, 7, 71–75. [Google Scholar] [CrossRef]
- Goiris, K.; Van Colen, W.; Wilches, I.; León-Tamariz, F.; De Cooman, L.; Muylaert, K. Impact of nutrient stress on antioxidant production in three species of microalgae. Algal Res. 2015, 7, 51–57. [Google Scholar] [CrossRef]
- Mostafa, S.S. Microalgal biotechnology: Prospects and applications. Plant Sci. 2012, 12, 276–314. [Google Scholar]
- Shanab, S.M.; Mostafa, S.S.; Shalaby, E.A.; Mahmoud, G.I. Aqueous extracts of microalgae exhibit antioxidant and anticancer activities. Asian Pac. J. Trop. Biomed. 2012, 2, 608–615. [Google Scholar] [CrossRef] [Green Version]
- Suparmi, S.; Fasitasari, M.; Martosupono, M.; Mangimbulude, J.C. Comparisons of curative effects of chlorophyll from Sauropus androgynus (L.) Merr leaf extract and Cu-chlorophyllin on sodium nitrate-induced oxidative stress in rats. J. Toxicol. 2016, 7, 8515089. [Google Scholar]
- Matsukawa, R.; Hotta, M.; Masuda, Y.; Chihara, M.; Karube, I. Antioxidants from carbon dioxide fixing Chlorella sorokiniana. J Appl. Phycol. 2000, 12, 263–267. [Google Scholar] [CrossRef]
- Rodrigues, D.B.; Menezes, C.R.; Mercadante, A.Z.; Jacob-Lopes, E.; Zepka, L.Q. Bioactive pigments from microalgae Phormidium autumnale. Food Res. Int. 2015, 77, 273–279. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; de Morais, R.M.S.C.; de Morais, A.M.M.B. Health applications of bioactive compounds from marine microalgae. Life Sci. 2013, 93, 479–486. [Google Scholar] [CrossRef]
- Shannon, E.; Abu-Ghannam, N. Antibacterial derivatives of marine algae: An overview of pharmacological mechanisms and applications. Mar. Drugs 2016, 14, 81. [Google Scholar] [CrossRef]
- Pratt, R.; Daniels, T.C.; Eiler, J.J.; Gunnison, J.B.; Kumler, W.D.; Oneto, J.F.; Strait, L.A.; Spoehr, H.A.; Hardin, G.J.; Milner, H.W.; et al. Chlorellin, an antibacterial substance from Chlorella. Science 1944, 351–352. [Google Scholar] [CrossRef]
- Najdenski, H.M.; Gigova, L.G.; Iliev, I.I.; Pilarski, P.S.; Lukavský, J.; Tsvetkova, I.V.; Ninova, M.S.; Kussovski, V.K. Antibacterial and antifungal activities of selected microalgae and cyanobacteria. Int. J. Sci. Food Technol. 2013, 48, 1533–1540. [Google Scholar]
- Smith, V.J.; Desbois, A.P.; Dyrynda, E.A. Conventional and unconventional antimicrobials from fish, marine invertebrates and micro-algae. Mar. Drugs 2010, 8, 1213–1262. [Google Scholar] [CrossRef] [Green Version]
- Amaro, H.M.; Guedes, A.C.; Malcata, F.X. Antimicrobial activities of microalgae: An invited review. In Science Against Microbial Pathogens: Communicating Current Research and Technological Advances; Mendez-Vilas, A., Ed.; Formatex Microbiology Book Series: Badajoz, Spain, 2011; pp. 1272–1284. ISBN 978-8-4939-8432-8. [Google Scholar]
- Cannell, R.J.; Owsianka, A.M.; Walker, J.M. Results of a large-scale screening programme to detect antibacterial activity from freshwater algae. Br. J. Phycol. 1988, 23, 41–44. [Google Scholar] [CrossRef]
- Falaise, C.; François, C.; Travers, M.A.; Morga, B.; Haure, J.; Tremblay, R.; Turcotte, F.; Pasetto, P.; Gastineau, R.; Hardivillier, Y.; et al. Antimicrobial compounds from eukaryotic microalgae against human pathogens and diseases in aquaculture. Mar. Drugs 2016, 14, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudimu, O.; Rybalka, N.; Bauersachs, T.; Born, J.; Friedl, T.; Schulz, R. Biotechnological screening of microalgal and cyanobacterial strains for biogas production and antibacterial and antifungal effects. Metabolites 2014, 4, 373–393. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P.; Mearns-Spragg, A.; Smith, V.J. A fatty acid from the diatom Phaeodactylum tricornutum is antibacterial against diverse bacteria including multi-resistant Staphylococcus aureus (MRSA). Mar. Biotechnol. 2009, 11, 45–52. [Google Scholar] [CrossRef]
- Rodríguez-Meizoso, I.; Jaime, L.; Santoyo, S.; Señoráns, F.J.; Cifuentes, A.; Ibáñez, E. Subcritical water extraction and characterization of bioactive compounds from Haematococcus pluvialis microalga. J. Pharm. Biomed. 2010, 51, 456–463. [Google Scholar]
- Prakash, J.W.; Marimuthu, J.; Jeeva, S. Antimicrobial activity of certain fresh water microalgae from Thamirabarani River, Tamil Nadu, South India. Asian Pac. J. Trop. Biomed. 2011, 1, S170–S173. [Google Scholar] [CrossRef]
- Pane, G.; Cacciola, G.; Giacco, E.; Mariottini, G.L.; Coppo, E. Assessment of the antimicrobial activity of algae extracts on bacteria responsible of external otitis. Mar. Drugs 2015, 13, 6440–6452. [Google Scholar] [CrossRef] [PubMed]
- Rendueles, O.; Ghigo, J.M. Multi-species biofilms: How to avoid unfriendly neighbors. FEMS Microbiol. Rev. 2012, 36, 972–989. [Google Scholar] [CrossRef]
- Yasuhara-Bell, J.; Lu, Y. Marine compounds and their antiviral activities. Antivir. Res. 2010, 86, 231–240. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Menakha, M. Pharmaceutical applications of cyanobacteria—A review. J. Acute Med. 2015, 5, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Gerber, P.; Dutcher, J.D.; Adams, E.V.; Sherman, J.H. Protective effect of seaweed extracts for chicken embryos infected with influenza B or mumps virus. Proc. Soc. Exp. Biol. Med. 1958, 99, 590–593. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Hayashi, K.; Maeda, M.; Kojima, I. Calcium spirulan, an inhibitor of enveloped virus replication, from a blue-green alga Spirulina platensis. J. Nat. Prod. 1996, 59, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.; Salomon, P.S.; Hamerski, L.; Walter, J.; Menezes, R.B.; Siqueira, J.E.; Santos, A.; Santos, J.A.M.; Ferme, N.; Guimarães, T.; et al. Inhibitory effect of microalgae and cyanobacteria extracts on influenza virus replication and neuraminidase activity. PeerJ 2018, 6, e5716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoyo, S.; Jaime, L.; Plaza, M.; Herrero, M.; Rodriguez-Meizoso, I.; Ibañez, E.; Reglero, G. Antiviral compounds obtained from microalgae commonly used as carotenoid sources. J. Appl. Phycol. 2012, 24, 731–741. [Google Scholar] [CrossRef] [Green Version]
- Huheihel, M.; Ishanu, V.; Tal, J.; Arad, S.M. Activity of Porphyridium sp. polysaccharide against herpes simplex viruses in vitro and in vivo. J. Biochem. Biophys. Methods 2002, 50, 189–200. [Google Scholar] [CrossRef]
- Talyshinsky, M.M.; Souprun, Y.Y.; Huleihel, M.M. Anti-viral activity of red microalgal polysaccharides against retroviruses. Cancer Cell Int. 2002, 2, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radonić, A.; Thulke, S.; Achenbach, J.; Kurth, A.; Vreemann, A.; König, T.; Walter, C.; Possinger, K.; Nitsche, A. Anionic polysaccharides from phototrophic microorganisms exhibit antiviral activities to Vaccinia virus. J. Antivir. Antiretrovir. 2011, 2, 51–55. [Google Scholar]
- Lee, J.B.; Hayashi, K.; Hirata, M.; Kuroda, E.; Suzuki, E.; Kubo, Y.; Hayashi, T. Antiviral sulfated polysaccharide from Navicula directa, a diatom collected from deep-sea water in Toyama Bay. Biol. Pharm. Bull. 2006, 29, 2135–2139. [Google Scholar] [CrossRef] [Green Version]
- Graf, C.; Bernkop-Schnürch, A.; Egyed, A.; Koller, C.; Prieschl-Grassauer, E.; Morokutti-Kurz, M. Development of a nasal spray containing xylometazoline hydrochloride and iota-carrageenan for the symptomatic relief of nasal congestion caused by rhinitis and sinusitis. Int. J. Gen. Med. 2018, 11, 275–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alangaden, G.J. Nosocomial fungal infections: Epidemiology, infection control, and prevention. Infect. Dis. Clin. N. Am. 2011, 25, 201–225. [Google Scholar] [CrossRef]
- Manuel, R.J.; Kibbler, C.C. The epidemiology and prevention of invasive Aspergillosis. J. Hospit. Infect. 1998, 39, 95–109. [Google Scholar] [CrossRef]
- Wey, S.B.; Mori, M.; Pfaller, M.A.; Woolson, R.F.; Wenzel, R.P. Hospital-acquired candidemia: The attributable mortality and excess length of stay. Arch. Intern. Med. 1988, 148, 2642–2645. [Google Scholar] [CrossRef]
- Washida, K.; Koyama, T.; Yamada, K.; Kita, M.; Uemura, D. Karatungiols A and B, two novel antimicrobial polyol compounds, from the symbiotic marine dinoflagellate Amphidinium sp. Tetrahedron Lett. 2006, 47, 2521–2525. [Google Scholar] [CrossRef]
- Gueho, E.; Pesando, D.; Barelli, M. Proprietes antifongiques d’une diatomee Chaetoceros lauderi ralfs CC. Mycopathologia 1977, 60, 105–107. [Google Scholar]
- Walter, C.S.; Mahesh, R. Antibacterial and antifungal activities of some marine diatoms in culture. Indian J. Mar. Sci. 2000, 29, 238–242. [Google Scholar]
- Ghannoum, M.A.; Rice, L.B. Antifungal agents: Mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clin. Microbiol. Rev. 1999, 12, 501–517. [Google Scholar] [CrossRef] [Green Version]
- Gastineau, R.; Hardivillier, Y.; Leignel, V.; Tekaya, N.; Morançais, M.; Fleurence, J.; Davidovich, N.; Jacquette, B.; Gaudin, P.; Hellio, C.; et al. Greening effect on oysters and biological activities of the blue pigments produced by the diatom Haslea karadagensis (Naviculaceae). Aquaculture 2012, 368, 61–67. [Google Scholar]
- Ghasemi, Y.; Moradian, A.; Mohagheghzadeh, A.; Shokravi, S.; Morowvat, M.H. Antifungal and antibacterial activity of the microalgae collected from paddy fields of Iran: Characterization of antimicrobial activity of Chroococcus dispersus. J. Biol. Sci. 2007, 7, 904–910. [Google Scholar] [CrossRef] [Green Version]
- Anand, P.; Kunnumakkara, A.B.; Sundaram, C.; Harikumar, K.B.; Tharakan, S.T.; Lai, O.S.; Sung, B.; Aggarwal, B.B. Cancer is a preventable disease that requires major lifestyle changes. Pharm. Res. 2008, 25, 2097–2116. [Google Scholar] [CrossRef]
- Chinery, R.; Brockman, J.A.; Peeler, M.O.; Shyr, Y.; Beauchamp, R.D.; Coffey, R.J. Antioxidants enhance the cytotoxicity of chemotherapeutic agents in colorectal cancer: A p53-independent induction of p21 WAF1/CIP1 via C/EBPβ. Nat. Med. 1997, 3, 1233–1241. [Google Scholar] [CrossRef] [PubMed]
- Burja, A.M.; Banaigs, B.; Abou-Mansour, E.; Burgess, J.G.; Wright, P.C. Marine cyanobacteria—A prolific source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar] [CrossRef]
- Singh, R.; Parihar, P.; Singh, M.; Bajguz, A.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Uncovering potential applications of cyanobacteria and algal metabolites in biology, agriculture and medicine: Current status and future prospects. Front. Microbiol. 2017, 8, 515. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liu, Y.; Gao, X.; Carter, C.L.; Liu, Z.R. The recombinant β subunit of C-phycocyanin inhibits cell proliferation and induces apoptosis. Cancer Lett. 2007, 247, 150–158. [Google Scholar] [CrossRef]
- Laroche, C.; Michaud, P. New developments and prospective applications for β (1, 3) glucans. Recent Pat. Biotechnol. 2007, 1, 59–73. [Google Scholar] [CrossRef]
- Sun, L.; Wang, L.; Zhou, Y. Immunomodulation and antitumor activities of different-molecular-weight polysaccharides from Porphyridium cruentum. Carbohydr. Polym. 2012, 87, 1206–1210. [Google Scholar] [CrossRef]
- Geresh, S.; Mamontov, A.; Weinstein, J. Sulfation of extracellular polysaccharides of red microalgae: Preparation, characterization and properties. J. Biochem. Biophys. Methods 2002, 50, 179–187. [Google Scholar] [CrossRef]
- Sheih, I.C.; Fang, T.J.; Wu, T.K.; Lin, P.H. Anticancer and antioxidant activities of the peptide fraction from algae protein waste. J. Agric. Food Chem. 2010, 58, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Talero, E.; García-Mauriño, S.; Ávila-Román, J.; Rodríguez-Luna, A.; Alcaide, A.; Motilva, V. Bioactive compounds isolated from microalgae in chronic inflammation and cancer. Mar. Drugs 2015, 13, 6152–6209. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.B.; Lichtman, A.H. Ch.2 Innate Immunity. In Basic Immunology. Functions and Disorders of the Immune System, 3rd ed.; Saunders (Elsevier): Philadelphia, PA, USA, 2009; p. 312. ISBN 978-1-4160-4688-2. [Google Scholar]
- Jellinger, K.A. Recent advances in our understanding of neurodegeneration. J. Neural. Transm. 2009, 116, 1111–1162. [Google Scholar]
- Popa-Wagner, A.; Mitran, S.; Sivanesan, S.; Chang, E.; Buga, A.M. ROS and brain diseases: The good, the bad, and the ugly. Oxid. Med. Cell. Longev. 2013, 963520. [Google Scholar] [CrossRef]
- Blagojević, D.; Babić, O.; Rašeta, M.; Šibul, F.; Janjušević, L.; Simeunović, J. Antioxidant activity and phenolic profile in filamentous cyanobacteria: The impact of nitrogen. J. Appl. Phycol. 2018, 30, 2337–2346. [Google Scholar] [CrossRef]
- Khan, M.N.A.; Cho, J.Y.; Lee, M.C.; Kang, J.Y.; Park, N.G.; Fujii, H.; Hong, Y.K. Isolation of two anti-inflammatory and one pro-inflammatory polyunsaturated fatty acids from the brown seaweed Undaria pinnatifida. J. Agric. Food Chem. 2007, 55, 6984–6988. [Google Scholar] [CrossRef] [PubMed]
- Steffens, D.; Lersch, M.; Rosa, A.; Scher, C.; Crestani, T.; Morais, M.G.; Costa, J.A.V.; Pranke, P. A new biomaterial of nanofibers with the microalga Spirulinaas scaffolds to cultivate with stem cells for use in tissue engineering. J. Biomed. Nanotechnol. 2013, 9, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Díaz, M, M.T.; Pérez, C.; Sánchez, C.I.; Lauzurica, S.; Cañeque, V.; González, C.; De La Fuente, J. Feeding microalgae increases omega 3 fatty acids of fat deposits and muscles in light lambs. J. Food Compos. Anal. 2017, 56, 115–123. [Google Scholar] [CrossRef]
- Nauroth, J.M.; Liu, Y.C.; Van Elswyk, M.; Bell, R.; Hall, E.B.; Chung, G.; Arterburn, L.M. Docosahexaenoic acid (DHA) and docosapentaenoic acid (DPAn-6) algal oils reduce inflammatory mediators in human peripheral mononuclear cells in vitro and paw edema in vivo. Lipids 2010, 45, 375–384. [Google Scholar] [CrossRef]
- Ohgami, K.; Shiratori, K.; Kotake, S.; Nishida, T.; Mizuki, N.; Yazawa, K.; Ohno, S. Effects of astaxanthin on lipopolysaccharide-induced inflammation in vitro and in vivo. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2694–2701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of antioxidants and natural products in inflammation. Oxid. Med. Cell Longev. 2016, 5276130. [Google Scholar] [CrossRef] [Green Version]
- Robertson, R.C.; Guihéneuf, F.; Bahar, B.; Schmid, M.; Stengel, D.B.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. The anti-inflammatory effect of algae-derived lipid extracts on lipopolysaccharide (LPS)-stimulated human THP-1 macrophages. Mar. Drugs 2015, 13, 5402–5424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.M.; Kim, K.N.; Lee, S.H.; Ahn, G.; Cha, S.H.; Kim, A.D.; Yang, X.D.; Kang, M.C.; Jeon, Y.J. Anti-inflammatory activity of polysaccharide purified from AMG-assistant extract of Ecklonia cava in LPS-stimulated RAW 264.7 macrophages. Carbohydr. Polym. 2011, 85, 80–85. [Google Scholar] [CrossRef]
- Zhao, P.; Zang, Z.; Xie, X.; Huang, A.; Wang, G. The influence of different flocculants on the physiological activity and fucoxanthin production of Phaeodactylum tricornutum. Process Biochem. 2014, 49, 681–687. [Google Scholar] [CrossRef]
- Senni, K.; Pereira, J.; Gueniche, F.; Delbarre-Ladrat, C.; Sinquin, C.; Ratiskol, J.; Godeau, G.; Fischer, A.M.; Helley, D.; Colliec-Jouault, S. Marine polysaccharides: A source of bioactive molecules for cell therapy and tissue engineering. Mar. Drugs 2011, 9, 1664–1681. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Lee, J.E.; Kim, K.H.; Kang, N.J. Beneficial effects of marine algae-derived carbohydrates for skin health. Mar. Drugs 2018, 16, 459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilal, M.; Rasheed, T.; Ahmed, I.; Iqbal, H.M.N. High value compounds from microalgae with industrial exploitability—A review. Front. Biosci. 2017, 9, 319–342. [Google Scholar]
- Muñoz-Durango, N.; Fuentes, C.A.; Castillo, A.E.; González-Gómez, L.M.; Vecchiola, A.; Fardella, C.E.; Kalergis, A.M. Role of the renin-angiotensin-aldosterone system beyond blood pressure regulation: Molecular and cellular mechanisms involved in end-organ damage during arterial hypertension. Int. J. Mol. Sci. 2016, 17, 797. [Google Scholar]
- Solomon, S.D.; Skali, H.; Bourgoun, M.; Fang, J.; Ghali, J.K.; Martelet, M.; Wojciechowski, D.; Ansmite, B.; Skards, J.; Laks, T.; et al. Effect of angiotensin-converting enzyme or vasopeptidase inhibition on ventricular size and function in patients with heart failure: The Omapatrilat Versus Enalapril Randomized Trial of Utility in Reducing Events (OVERTURE) echocardiographic study. Am. Heart J. 2005, 150, 257–262. [Google Scholar]
- Ko, S.C.; Kang, N.; Kim, E.A.; Kang, M.C.; Lee, S.H.; Kang, S.M.; Lee, J.B.; Jeon, B.T.; Kim, S.K.; Park, S.J.; et al. A novel angiotensin I-converting enzyme (ACE) inhibitory peptide from a marine Chlorella ellipsoidea and its antihypertensive effect in spontaneously hypertensive rats. Process Biochem. 2012, 47, 2005–2011. [Google Scholar] [CrossRef]
- Samarakoon, K.W.; Kwon, O.N.; Ko, J.Y.; Lee, J.H.; Kang, M.C.; Kim, D.; Lee, J.B.; Lee, J.S.; Jeon, Y.J. Purification and identification of novel angiotensin-I converting enzyme (ACE) inhibitory peptides from cultured marine microalgae (Nannochloropsis oculata) protein hydrolysate. J. Appl. Phycol. 2013, 25, 1595–1606. [Google Scholar]
- Raposo, M.F.D.J.; Oliveira, S.E.; Castro, P.M.; Bandarra, N.M.; Morais, R.M. On the utilization of microalgae for brewery effluent treatment and possible applications of the produced biomass. J. Inst. Brew. 2010, 116, 285–292. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Hayashi, T.; Lee, J.B.; Ozawa, T.; Sakuragawa, N. Activation of heparin cofactor II by calcium Spirulan. J. Biol. Chem. 2000, 275, 11379–11382. [Google Scholar] [CrossRef] [Green Version]
- Taylor, V.F.; Jackson, B.P. Concentrations and speciation of arsenic in New England seaweed species harvested for food and agriculture. Chemosphere 2016, 163, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Das, B.K.; Pradhan, J.; Pattnaik, P.; Samantaray, B.R.; Samal, S.K. Production of antibacterials from the freshwater alga Euglena viridis (Ehren). World J. Microbiol. Biotechnol. 2005, 21, 45–50. [Google Scholar] [CrossRef]
- Bhagavathy, S.; Sumathi, P.; Bell, I.J.S. Green algae Chlorococcum humicola -a new source of bioactive compounds with antimicrobial activity. Asian Pac. J. Trop Biomed. 2011, 1, S1–S7. [Google Scholar] [CrossRef]
- Hasui, M.; Matsuda, M.; Okutani, K.; Shigeta, S.; Hasui, M. In vitro antiviral activities of sulfated polysaccharides from a marine microalga (Cochlodinium polykrikoides) against human immunodeficiency virus and other enveloped viruses. Int. J. Biol. Macromol. 1995, 17, 293–297. [Google Scholar] [CrossRef]
- Li, H.B.; Cheng, K.W.; Wong, C.C.; Fan, K.W.; Chen, F.; Jiang, Y. Evaluation of antioxidant capacity and total phenolic content of different fractions of selected microalgae. Food Chem. 2007, 102, 771–776. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration (HHS). FDA Summary: Substances Generally Regarded as Safe (Final Rule). 2016. Available online: http://www.fda.gov/AboutFDA/ReportsManualsForms/Reports/EconomicAnalyses/ucm517103.htm (accessed on 24 October 2016).
- Wijffels, R.H. Potential of sponges and microalgae for marine biotechnology. Trends Biotechnol. 2008, 26, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Europen Food Safety Auhtority (EFSA). EU Novel Food Catalogue (v 1.1.). 2020. Available online: http://ec.europa.eu/food/safety/novel_food/catalogue_en (accessed on 16 April 2020).
- Kay, R.A.; Barton, L.L. Microalgae as food and supplement. Crit. Rev. Food. Sci. Nutr. 1991, 30, 555–573. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.R.; Deviram, G.; Mathimani, T.; Duc, P.A.; Pugazhendhi, A. Microalgae as rich source of polyunsaturated fatty acids. Biocatal. Agric. Biotechnol. 2019, 17, 583–588. [Google Scholar] [CrossRef]
- Hemaiswarya, S.; Raja, R.; Kumar, R.R.; Ganesan, V.; Anbazhagan, C. Microalgae: A sustainable feed source for aquaculture. World J. Microbiol. Biotechnol. 2011, 27, 1737–1746. [Google Scholar] [CrossRef]
- Apandi, N.M.; Mohamed, R.M.S.R.; Al-Gheethi, A.; Kassim, A.H.M. Microalgal biomass production through phycoremediation of fresh market wastewater and potential applications as aquaculture feeds. Environ. Sci. Pollut. Res. 2019, 26, 3226–3242. [Google Scholar] [CrossRef]
- Yucetepe, A.; Saroglu, O.; Bildik, F.; Ozcelik, B.; Daskaya-Dikmen, C. Optimisation of ultrasound-assisted extraction of protein from Spirulina platensis using RSM. Czech J. Food Sci. 2018, 36, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Madeira, M.S.; Cardoso, C.; Lopes, P.A.; Coelho, D.; Afonso, C.; Bandarra, N.M.; Prates, J.A. Microalgae as feed ingredients for livestock production and meat quality: A review. Livest. Sci. 2017, 205, 111–121. [Google Scholar] [CrossRef]
- Sui, Y.; Vlaeminck, S.E. Dunaliella microalgae for nutritional protein: An undervalued asset. Trends Biotechnol. 2020, 38, 10–12. [Google Scholar] [CrossRef]
- Yaakob, Z.; Ali, E.; Zainal, A.; Mohamad, M.; Takriff, M.S. An overview: Biomolecules from microalgae for animal feed and aquaculture. J. Biol. Res. 2014, 21, 6. [Google Scholar]
- Nonwachai, T.; Purivirojkul, W.; Limsuwan, C.; Chuchird, N.; Velasco, M.; Dhar, A.K. Growth, nonspecific immune characteristics, and survival upon challenge with Vibrio harveyi in Pacific white shrimp (Litopenaeus vannamei) raised on diets containing algal meal. Fish Shellfish Immun. 2010, 29, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Dineshbabu, G.; Goswami, G.; Kumar, R.; Sinha, A.; Das, D. Microalgae–nutritious, sustainable aqua-and animal feed source. J. Funct Foods. 2019, 62, 103545. [Google Scholar]
- Guedes, A, A.C.; Malcata, F.X. Nutritional value and uses of microalgae in aquaculture. Aquaculture 2012, 10, 59–78. [Google Scholar]
- Neupert, J.; Shao, N.; Lu, Y.; Bock, R. Genetic transformation of the model green alga Chlamydomonas reinhardtii. In Transgenic Plants; Dunwell, J., Wetten, A., Eds.; Springer: Cham, Switzerland, 2012. [Google Scholar]
- Costa, D.F.A.; Quigley, S.P.; Isherwood, P.; McLennan, S.R.; Poppi, D.P. Supplementation of cattle fed tropical grasses with microalgae increases microbial protein production and average daily gain. J. Anim. Sci. 2016, 94, 2047–2058. [Google Scholar] [CrossRef]
- Kulpys, J.; Paulauskas, E.; Pilipavicius, V.; Stankevicius, R. Influence of cyanobacteria Arthrospira (Spirulina) platensis biomass additive towards the body condition of lactation cows and biochemical milk indexes. Agron. Res. 2009, 7, 823–835. [Google Scholar]
- Kholif, A.E.; Morsy, T.A.; Matloup, O.H.; Anele, U.Y.; Mohamed, A.G.; El-Sayed, A.B. Dietary Chlorella vulgaris microalgae improves feed utilization, milk production and concentrations of conjugated linoleic acids in the milk of Damascus goats. J. Agric. Sci. 2017, 155, 508–518. [Google Scholar] [CrossRef]
- Shimkiene, A.; Bartkevichiute, Z.; Chernauskiene, J.; Shimkus, A.; Chernauskas, A.; Ostapchuk, A.; Nevitov, M.; Shimkiene, A. The influence of Spirulina platensis and concentrates on lambs’ growth. Zhivotnov’dni Nauki 2010, 47, 9–14. [Google Scholar]
- Ekmay, R.; Gatrell, S.; Lum, K.; Kim, J.; Lei, X.G. Nutritional and metabolic impacts of a defatted green marine microalgal (Desmodesmus sp.) biomass in diets for weanling pigs and broiler chickens. J. Agric. Food Chem. 2014, 62, 9783–9791. [Google Scholar]
- Wu, Y.B.; Li, L.; Wen, Z.G.; Yan, H.J.; Yang, P.L.; Tang, J.; Xie, M.; Hou, S.S. Dual functions of eicosapentaenoic acid-rich microalgae: Enrichment of yolk with n-3 polyunsaturated fatty acids and partial replacement for soybean meal in diet of laying hens. Poult. Sci. 2019, 98, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Bruneel, C.; Lemahieu, C.; Fraeye, I.; Ryckebosch, E.; Muylaert, K.; Buyse, J.; Foubert, I. Impact of microalgal feed supplementation on omega-3 fatty acid enrichment of hen eggs. J. Funct. Foods 2013, 5, 897–904. [Google Scholar] [CrossRef]
- Gatrell, S.; Lum, K.; Kim, J.; Lei, X.G. Nonruminant Nutrition Symposium: Potential of defatted microalgae from the biofuel industry as an ingredient to replace corn and soybean meal in swine and poultry diets. J. Anim. Sci. 2014, 92, 1306–1314. [Google Scholar]
- Qiu, S.; Wang, S.; Xiao, C.; Ge, S. Assessment of microalgae as a new feeding additive for fruit fly Drosophila melanogaster. Sci. Total Environ. 2019, 667, 455–463. [Google Scholar] [CrossRef]
- Daneshvar, E.; Ok, Y.S.; Tavakoli, S.; Sarkar, B.; Shaheen, S.M.; Hong, H.; Luo, Y.; Rinklebe, J.; Song, H.; Bhatnagar, A. Insights into upstream processing of microalgae: A review. Bioresour. Technol. 2021, 329, 124870. [Google Scholar] [CrossRef] [PubMed]
- Narala, R.R.; Garg, S.; Sharma, K.K.; Thomas-Hall, S.R.; Deme, M.; Li, Y.; Schenk, P.M. Comparison of microalgae cultivation in photobioreactor, open raceway pond, and a two-stage hybrid system. Front. Energy Res. 2016, 4, 29. [Google Scholar] [CrossRef]
- Coêlho, D.D.F.; Tundisi, L.L.; Cerqueira, K.S.; Rodrigues, J.R.D.S.; Mazzola, P.G.; Tambourgi, E.B.; Souza, R.R.D. Microalgae: Cultivation aspects and bioactive compounds. Braz. Arch. Biol. Technol. 2019, 62, e19180343. [Google Scholar]
- Ranadheer, P.; Kona, R.; Sreeharsha, R.V.; Venkata Mohan, S. Non-lethal nitrate supplementation enhances photosystem II efficiency in mixotrophic microalgae towards the synthesis of proteins and lipids. Bioresour. Technol. 2019, 283, 373–377. [Google Scholar] [CrossRef]
- Chiranjeevi, P.; Venkata Mohan, S. Diverse acidogenic effluents as feedstock for microalgae cultivation: Dual phase metabolic transition on biomass growth and lipid synthesis. Bioresour. Technol. 2017, 242, 191–196. [Google Scholar] [CrossRef]
- Suparmaniam, U.; Lam, M.K.; Uemura, Y.; Lim, J.W.; Lee, K.T.; Shuit, S.H. Insights into the microalgae cultivation technology and harvesting process for biofuel production: A review. Renew. Sust. Energ. Rev. 2019, 115, 109361. [Google Scholar] [CrossRef]
- Chaisutyakorn, P.; Praiboon, J.; Kaewsuralikhit, C. The effect of temperature on growth and lipid and fatty acid composition on marine microalgae used for biodiesel production. J. Appl. Phycol. 2018, 30, 37–45. [Google Scholar] [CrossRef]
- Khalil, Z.I.; Asker, M.M.; El-Sayed, S.; Kobbia, I.A. Effect of pH on growth and biochemical responses of Dunaliella bardawil and Chlorella ellipsoidea. World J. Microbiol. Biotechnol. 2010, 26, 1225–1231. [Google Scholar] [CrossRef]
- Venkata Mohan, S.; Devi, M.P. Salinity stress induced lipid synthesis to harness biodiesel during dual mode cultivation of mixotrophic microalgae. Bioresour. Technol. 2014, 165, 288–294. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, C.Y.B.; Viegas, T.L.; da Silva, M.F.O.; Fracalossi, D.M.; Lopes, R.G.; Derner, R.B. Effect of trace metals on growth performance and accumulation of lipids, proteins, and carbohydrates on the green microalga Scenedesmus obliquus. Aquac. Int. 2020; 1–10. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Madamwar, D.; Nakamoto, H.; Incharoensakdi, A. Resilience and self-regulation processes of microalgae under UV radiation stress. J. Photochem. Photobiol. C Photochem. Rev. 2019, 100322. [Google Scholar] [CrossRef]
- Singh, G.; Patidar, S.K. Microalgae harvesting techniques: A review. J. Environ. Manag. 2018, 217, 499–508. [Google Scholar] [CrossRef]
- Saha, S.K.; Murray, P. Exploitation of microalgae species for nutraceutical purposes: Cultivation aspects. Fermentation 2018, 4, 46. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.; Mahfuzur, R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-producing green microalga Haematococcus pluvialis: From single cell to high value commercial products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemalatha, M.; Sravan, J.S.; Min, B.; Venkata Mohan, S. Microalgae-biorefinery with cascading resource recovery design associated to dairy wastewater treatment. Bioresour. Technol. 2019, 284, 424–429. [Google Scholar] [CrossRef]
- Venkata Mohan, S.; Devi, M.P.; Mohanakrishna, G.; Amarnath, N.; Babu, M.L.; Sarma, P.N. Potential of mixed microalgae to harness biodiesel from ecological water-bodies with simultaneous treatment. Bioresour. Technol. 2011, 102, 1109–1117. [Google Scholar] [CrossRef]
- Aziz, M.M.A.; Kassim, K.A.; Shokravi, Z.; Jakarni, F.M.; Liu, H.Y.; Zaini, N.; Tan, L.S.; Islam, A.S.; Shokravi, H. Two-stage cultivation strategy for simultaneous increases in growth rate and lipid content of microalgae: A review. Renew. Sust. Energ. Rev. 2020, 119, 109621. [Google Scholar] [CrossRef]
- Devi, M.P.; Subhash, G.V.; Venkata Mohan, S. Heterotrophic cultivation of mixed microalgae for lipid accumulation and wastewater treatment during sequential growth and starvation phases: Effect of nutrient supplementation. Renew Energy 2012, 43, 276–283. [Google Scholar] [CrossRef]
- Gangl, D.; Zedler, J.A.; Rajakumar, P.D.; Martinez, E.M.; Riseley, A.; Włodarczyk, A.; Purton, S.; Sakuragi, Y.; Howe, C.J.; Jensen, P.E.; et al. Biotechnological exploitation of microalgae. J. Exp. Bot. 2015, 66, 6975–6990. [Google Scholar]
- Caetano, N.S.; Martins, A.A.; Gorgich, M.; Gutiérrez, D.M.; Ribeiro, T.J.; Ribeiro, T.J.; Mata, T.M. Flocculation of Arthrospira maxima for improved harvesting. Energy Rep. 2020, 6, 423–428. [Google Scholar] [CrossRef]
- Monte, J.; Sá, M.; Galinha, C.F.; Costa, L.; Hoekstra, H.; Brazinha, C.; Crespo, J.G. Harvesting of Dunaliella salina by membrane filtration at pilot scale. Sep. Purif. Technol. 2018, 190, 252–260. [Google Scholar] [CrossRef]
- Sharma, K.K.; Garg, S.; Li, Y.; Malekizadeh, A.; Schenk, P.M. Critical analysis of current microalgae dewatering techniques. Biofuels 2013, 4, 397–407. [Google Scholar] [CrossRef]
- Okoro, V.; Azimov, U.; Munoz, J.; Hernandez, H.H.; Phan, A.N. Microalgae cultivation and harvesting: Growth performance and use of flocculants—A review. Renew. Sustain. Energy Rev. 2019, 115, 109364. [Google Scholar] [CrossRef]
- Mathimani, T.; Mallick, N. A comprehensive review on harvesting of microalgae for biodiesel–key challenges and future directions. Renew. Sustain. Energy Rev. 2018, 91, 1103–1120. [Google Scholar] [CrossRef]
- Safi, C.; Frances, C.; Ursu, A.V.; Laroche, C.; Pouzet, C.; Vaca-Garcia, C.; Pontalier, P.Y. Understanding the effect of cell disruption methods on the diffusion of Chlorella vulgaris proteins and pigments in the aqueous phase. Algal Res. 2015, 8, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Phong, W.N.; Show, P.L.; Ling, T.C.; Juan, J.C.; Ng, E.P.; Chang, J.S.; Pouzet, C.; Vaca-Garcia, C. Mild cell disruption methods for bio-functional proteins recovery from microalgae—Recent developments and future perspectives. Algal Res. 2018, 31, 506–516. [Google Scholar] [CrossRef]
- Günerken, E.; D’Hondt, E.; Eppink, M.H.M.; Garcia-Gonzalez, L.; Elst, K.; Wijffels, R.H. Cell disruption for microalgae biorefineries. Biotechnol. Adv. 2015, 33, 243–260. [Google Scholar]
- Lorente, E.; Hapońska, M.; Clavero, E.; Torras, C.; Salvadó, J. Microalgae fractionation using steam explosion, dynamic and tangential cross-flow membrane filtration. Bioresour. Technol. 2017, 237, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Postma, P.R.; Miron, T.L.; Olivieri, G.; Barbosa, M.J.; Wijffels, R.H.; Eppink, M.H.M. Mild disintegration of the green microalgae Chlorella vulgaris using bead milling. Bioresour. Technol. 2015, 184, 297–304. [Google Scholar]
- Cuellar-Bermudez, S.P.; Aguilar-Hernandez, I.; Cardenas-Chavez, D.L.; Ornelas-Soto, N.; Romero-Ogawa, M.A.; Parra-Saldivar, R. Extraction and purification of high-value metabolites from microalgae: Essential lipids, astaxanthin and phycobiliproteins. Microb. Biotechnol. 2015, 8, 190–209. [Google Scholar] [CrossRef]
- Tiwari, B.K. Ultrasound: A clean, green extraction technology. Trends Anal. Chem. 2015, 71, 100–109. [Google Scholar] [CrossRef]
- Mittal, R.; Tavanandi, H.A.; Mantri, V.A.; Raghavarao, K.S.M.S. Ultrasound assisted methods for enhanced extraction of Phycobiliproteins from marine macro-algae, Gelidium pusillum (Rhodophyta). Ultrason. Sonochem. 2017, 38, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Sutanto, H.; Suzery, M. Phyocyanin extraction from microalgae Spirulina platensis assisted by ultrasound irradiation: Effect of time and temperature. Songklanakarin J. Sci. Technol. 2016, 38, 391–398. [Google Scholar]
- Tan, J.S.; Lee, S.Y.; Chew, K.W.; Lam, M.K.; Lim, J.W.; Ho, S.H.; Show, P.L. A review on microalgae cultivation and harvesting, and their biomass extraction processing using ionic liquids. Bioengineered 2020, 11, 116–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapoore, R.V.; Butler, T.O.; Pandhal, J.; Vaidyanathan, S. Microwave-assisted extraction for microalgae: From biofuels to biorefinery. Biology 2018, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Cooney, M.; Young, G.; Nagle, N. Extraction of bio-oils from microalgae. Sep. Purif. Rev. 2016, 38, 291–325. [Google Scholar] [CrossRef]
- Rodriguez-Jasso, R.M.; Mussatto, S.I.; Pastrana, L.; Aguilar, C.N.; Teixeira, J.A. Microwave-assisted extraction of sulfated polysaccharides (fucoidan) from brown seaweed. Carbohydr. Polym. 2011, 86, 1137–1144. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.Y.; Vijayan, D.; Praveenkumar, R.; Han, J.I.; Lee, K.; Park, J.Y.; Chang, W.S.; Lee, J.S.; Oh, Y.K. Cell-wall disruption and lipid/astaxanthin extraction from microalgae: Chlorella and Haematococcus. Bioresour. Technol. 2016, 199, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Molino, D.A.; Mehariya, S.; Di Sanzo, G.; Larocca, V.; Martino, M.; Leone, G.P.; Marino, T.; Chianese, S.; Balducchi, R.; Musmarra, D. Recent developments in supercritical fluid extraction of bioactive compounds from microalgae: Role of key parameters, technological achievements and challenges. J. CO2 Util. 2020, 36, 196–209. [Google Scholar]
- Singh, D.; Gupta, A.; Wilkens, S.L.; Mathur, A.S.; Tuli, D.K.; Barrow, C.J.; Puri, M. Understanding response surface optimisation to the modeling of Astaxanthin extraction from a novel strain Thraustochytrium sp. S7. Algal Res. Algal Res. 2015, 11, 113–120. [Google Scholar]
- Negi, S.; Pandey, A.K. Ionic liquid pretreatment. Pretreat. Biomass Process. Technol. 2014, 137–155. [Google Scholar] [CrossRef]
- Lee, S.Y.; Vicente, F.A.; De Silva, F.A.; Sintra, T.E.; Taha, M.; Khoiroh, I.; Coutinho, J.A.; Show, P.L.; Ventura, S.P. Evaluating self-buffering ionic liquids for biotechnological applications. ACS Sustain. Chem Eng. 2015, 3, 3420–3428. [Google Scholar] [CrossRef]
- Zinnai, A.; Sanmartin, C.; Taglieri, I.; Andrich, G.; Venturi, F. Supercritical fluid extraction from microalgae with high content of LC-PUFAs. A case of study: Sc-CO2 oil extraction from Schizochytrium sp. J. Supercrit. Fluids 2016, 116, 126–131. [Google Scholar]
- Leone, G.P.; Balducchi, R.; Mehariya, S.; Martino, M.; Larocca, V.; Di Sanzo, G.; Iovine, A.; Casella, P.; Marino, T.; Karatza, D.; et al. Selective Extraction of ω-3 fatty acids from Nannochloropsis sp. using supercritical CO2 extraction. Molecules 2019, 24, 2406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molino, A.; Martino, M.; Larocca, V.; Di Sanzo, G.; Spagnoletta, A.; Marino, T.; Karatza, D.; Iovine, A.; Mehariya, S.; Musmarra, D. Eicosapentaenoic acid extraction from Nannochloropsis gaditana using carbon dioxide at supercritical conditions. Mar. Drugs 2019, 17, 132. [Google Scholar]
- Herrero, M.; Sanchez-Camargo, A.P.; Cifuentes, A.; Ibanez, E. Plants, seaweeds, micro-algae and food by-products as natural sources of functional ingredients obtained using pressurized liquid extraction and supercritical fluid extraction. Trends Anal. Chem. 2015, 71, 26–38. [Google Scholar] [CrossRef]
- Gallego, R.; Montero, L.; Cifuentes, A.; Ibáñez, E.; Herrero, M. Green extraction of bioactive compounds from microalgae. J. Anal. Test. 2018, 2, 109–123. [Google Scholar] [CrossRef]
- Gilbert-López, B.; Barranco, A.; Herrero, M.; Cifuentes, A.; Ibáñez, E. Development of new green processes for the recovery of bioactives from Phaeodactylum tricornutum. Food Res. Int. 2017, 99, 1056–1065. [Google Scholar] [CrossRef] [Green Version]
- Gerken, H.G.; Donohoe, B.; Knoshaug, E.P. Enzymatic cell wall degradation of Chlorellavulgaris and other microalgae for biofuels production. Planta 2013, 237, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Hettiarachchy, N.S.; Shellhammer, T.H. Protein extraction from heat-stabilized defatted rice bran. 1. Physical processing and enzyme treatments. J. Agric. Food Chem. 2002, 50, 7444–7448. [Google Scholar] [CrossRef]
- Kang, K.H.; Qian, Z.J.; Ryu, B.; Karadeniz, F.; Kim, D.; Kim, S.K. Antioxidant peptides from protein hydrolysate of microalgae Navicula incerta and their protective effects in HepG2/CYP2E1 cells induced by ethanol. Phyther. Res. 2012, 26, 1555–1563. [Google Scholar] [CrossRef]
- Taher, H.; Al-Zuhair, S.; Al-Marzouqi, A.H.; Haik, Y.; Farid, M. Effective extraction of microalgae lipids from wet biomass for biodiesel production. Biomass Bioenergy 2014, 66, 159–167. [Google Scholar] [CrossRef]


{kind=link}
| S. No. | Microalgae Compounds | Pigment | Microalgae Species | Activity on Human Health | Reference |
|---|---|---|---|---|---|
| 1. | Chlorophyll | Chlorophyll | Chlorella sp., Sanropus androgynous, Green algae | Food additive, antioxidant activity, immune activators, cytotoxic towards tumoral cells, | Khanra et al., 2018 [28]; Koller et al., 2014 [64]; Mishra et al., 2011 [27]; Odjadjare et al., 2017 [26]; www.oligae.com (accessed on 15 March 2021) |
| 2. | Carotenoids | β-carotene | Botryococcus braunii, Chlamydomonas nivalis, Chlamydocapsa sp., Chlorella sorokiniana, Chlorococcum sp., Chondria striolata, Dunaliella salina, Dunaliella tertiolecta, and Paeonia obovate | Anti-aging, cancer, immune control, coronary disease prevention, retinal and sensory disability enhancement and low-density lipoprotein oxidation inhibition | Andrade et al., 2018 [56]; Barkia et al., 2019 [12]; Galasso et al., 2019 [30]; Gong and Bassi 2016 [31] |
| Astaxanthin | Ankistrodesmus braunii, Chlamydomonasnivalis, Chlorella vulgaris, Chondria striolata, Haematococcus lacustris, Haematococcus pluvialis, Haematococcus sp., Monoraphidium sp., Scenedesmus obliquus | Cancer defense, inflammation, metabolic syndrome, diabetes, neurodegenerative and ocular diseases, lung injury, repressed alveolar wall swelling and myeloperoxidase activity | Cai et al., 2019 [65]; Capelli et al., 2019 [32]; Chang et al., 2010 [66]; Galasso et al., 2019 [30]; Park et al., 2010 [67]; Talukdar et al., 2020 [33]; Wu et al., 2014 [68] | ||
| Lutein | Ankistrodesmus braunii, Chlamydomonas acidophila, Chlorella fusca, Chlorella sorokiniana, Chlorococcum sp., Tetraselmis suecica | Antioxidant and anticancer activity, prevents macular degeneration, Cataract, atherosclerosis, diabetic retinopathy, and age-related retinal degeneration | Liu et al., 2017 [69]; Rasmussen and Johnson 2013 [70]; Sansone et al., 2017 [71] | ||
| Violaxanthin | Anti-proliferative activity | Koller et al., 2014 [64] | |||
| Canthaxanthin | Chlamydocapsa sp., Chlamydomonas nivalis, Chlorella vulgaris, Chlorococcum sp., Chlorella zofingiensis, Neospongiococcum sp. | Increases Vitamin E, antioxidative, anti-inflammatory and neuroprotective properties | Sathasivam et al., 2019 [41] | ||
| Fucoxanthin and Zeaxanthin | Chaetoceros gracilis, Chlamydomonasnivalis, Dunaliella salina Isochrysis sp., Ochromonas sp., Odontella aurita, Phaeodactylum tricornutum, Prymnesium parvum, Salpingoeca marina | Anti-cancer and anti-proliferative properties, prevention of osteoporosis, rheumatoid arthritis and diabetic diseases, suppressing insulin and hyperglycemia | Gong et al., 2016 [31]; Liu et al., 2017 [69] | ||
| 3. | Polysaccharides | Polysaccharides | Chlorella stigmatophora, Phaeodactylum tricornutum, Porphyridium cruentum, Rhodella reticulata | Antioxidant and tumoricidal activity, Reduces free radicals, atherosclerosis, blood cholesterol, gastric ulcers, sores, constipation and hypercholesterolemia | Chen et al., 2018 [45]; de Gardeva et al., 2009 [47]; Morais et al., 2015 [46] |
| Sulfonated polysaccharides | Chlorophyta, Phaeophyta and Rhodophyta | Anticancer, antifungal, hepatoprotective, antihelminthic, anti-protozoal, anti-inflammatory, anti-coagulant, immunomodulation and enhanced skin tissue regeneration. Reducing coronary heart disease and acts as coating material on the sanitary items for COVID-19 prevention | Gaikwad et al., 2020 [49]; Lekshmi and Krupa 2019 [48]; Morokutti-Kurz et al., 2017 [72]; Olasehinde et al., 2017 [73]; Raposo et al., 2014 [74] | ||
| 4. | Polyunsaturated Fatty Acids (PUFAs) | Chaetoceros calcitrans, Crythecodinium cohnii, Isochrysis galbana, Monodus subterraneus, Nannochloropsis sp., Pavlova salina, Phaeodactylum tricornutum, and Porphyridium cruentum | Reduces occurrence of chronic diseases such as obesity, arthritis, diabetes, cardiovascular diseases, hypercholesterolemia and improves brain function | Bhalamurugan et al., 2018 [42]; Katiyar and Arora 2020 [40]; Mourelle et al., 2017 [37] | |
| Eicosapentaenoic acid (EPA) | Chlorella vulgaris, Nannochloropsis sp., Pavlova sp., Tetraselmis sp. | Immune activator, blood clotting, regulation of blood pressure and prevents thrombosis, atherosclerosis and beneficial for coronal heart diseases | Charles et al., 2019 [43]; Levassuer et al., 2020 [11] | ||
| Docosahexaeonic acid (DHA) | Crypthecodiuimu sp., Pyramimonas sp., Schizochytrium sp., Thraustochytrid strain 12B | Anti-inflammatory, anticancer function, used in food for pregnant, nursing and cardiovascular patients as dietary supplements | Long et al., 2018 [75]; Raghukumar, 2008 [76]; Teng et al., 2015 [77] | ||
| Arachidonic acid (ARA) | Navicula atomus, Pediastrum boryanum, Porphyridium sp. | Platelet aggregators, vasoconstrictor, vasodilators and have antiaggregative action on the endothelium in neutrophils | de Morais et al., 2015 [46]; Paliwal et al., 2017 [52]; | ||
| γ-linoleic acid (GLA) | Anthrospira, Chlorococcum sp., Dunaliella primolecta, Spirulina sp. | Relieves from breast cancer, skin allergies, alcoholism, obesity, rheumatoid arthritis, blood pressure, heart diseases, premenstrual syndrome, sclerosis, hyperactivity attention deficit disorder (ADHD), diabetes-related neural issues | Koller et al., 2014 [64]; Mourelle et al., 2017 [78] | ||
| Linolenic acid | Botryococcus braunii, Chlorococcum sp., Dunaliella primolecta, Scenedesmus obliqus, Tetraselmis suecica | Anti-inflammatory, acne reductive and moisture retention | Day et al., 2009 [79] | ||
| 5. | Proteins | Glycoprotein | Alexandrium minutum, Chlorella sp., Dunaliella sp., Schizochytrium sp., Spirulina | Antihypertensive and Angiotensin I inhibitory activities, appetite suppression and reduction of LDL-cholesterol. It is also used as dietary supplements (tablets, powder and paste) | Andrade et al., 2018 [56]; Caporgno and Mathys 2018 [50]; da Silva Viz et al., 2016 [17]; Galasso et al., 2019 [30]; Hempel et al., 2011 [51]; Kim et al., 2012 [80];Marquez-Escobar et al., 2018 [81] |
| Phycobiliprotein | Cyanobacteria | Bio-sensor, neuroprotective, anti-nephrolithe, anti-hyperglycemic, immunomodulatory and hepatoprotective properties. Used as fluorescent labels in antibodies and receptors, flow-cytometry, immunohistochemistry | Bhattacharjee 2016 [82]; Paliwal et al., 2017 [52] | ||
| Recombinant protein | Chlamydomonas elipsoidea, Chlamydomonas reinhardtii; | Anemia treatment, wound healing, anti-malarial vaccines, antibody against anthrax, Herpes simplex virus, human papilloma virus, white spot syndrome virus, foot and mouth disease virus, immunotoxins against B-cell lymphoma and increase resistance to UV-induced stress | Brasil et al., 2017 [10]; Hempel et al., 2016 [53]; Specht et al., 2010 [55]; http://www.olson.com.br/ (accessed on 15 March 2021) | ||
| 6. | Amino acid | Mycosporine-like amino acid (MAA) | Chlorella pyrenoidosa, Chlorella vulgaris, Microcystis aeruginosa, Nitzschia incerta | Antioxidant properties, prevents atherosclerosis, cancer, coronary diseases and used in photo-aging protective formulations | Gregory et al., 2013 [54]; Kim et al., 2011 [83]; Lawrence et al., 2018 [84] |
| 7. | Vitamins | Vitamin A | Chlorella sp., Eisenia arborea, P. cruentum | Involved in vision, reproduction, immune function and cellular communication | Andrade et al., 2018 [56]; Koyande et al., 2019 [14]; |
| Vitamin B | Chlorella, Spirulina, Pavlova, Tetraselmis | Anticancer activity, reduces cholesterol, regeneration of blood cells, DNA repair, histone methylation, preservation of skin and mucous membranes and cardiovascular disease | Becker et al., 2004 [85]; Delasoie et al., 2018 [86] | ||
| Vitamin E | Chaetoceros calcitrans, Dunaliella tertiolecta, Nannochloropsis oculata, Porphyridium cruentum and Tetraselmis suecica | Protects membrane lipids from oxidative damage and prevent coronary, atherosclerosis as well as neurodegenerative diseases. Improves endothelial function, vascular health and inhibits prostate cancer cell growth | Bong et al., 2013 [60]; Giammanco et al., 2015 [59] | ||
| Vitamin K | Anabaena cylindrica | Protect against toxic pollutants, prevention of chronic diseases | Tarento et al., 2018 [58] | ||
| 8. | Sterols | Brassicasterol Stigmasterol Phytosterols | Chlorella vulgaris, Pavlova lutheria, Nanochloropsis sp. BR2 and Tetraselmis sp. M8 | Anti-inflammatory and anticancer activities, stabilizes phospholipid bilayers, reducing blood cholesterol levels in hyper and normocholesterolemic people and inhibit colon cancer development | Ahmed et al., 2015 [63]; Lopes et al., 2013 [61]; Luo et al., 2015 [62] |
| Antibacterial Activity | ||||
|---|---|---|---|---|
| S. No. | Microalgae species | Compound/Fraction | Targeted Microorganism | References |
| 1. | Bacillariophyceae and Chrysophyceae | Methanolic extracts, lysed cells, phycobiliproteins, beta-ionone and neophytadiene | Escherichia coli, Pseudomonas aeruginosa, Salmonella typhimurium, Enterobacter aerogenes, Klebsiella pneumoniae, Vibrio cholera and Proteus vulgaris | Cannell et al., 1988 [100]; Falaise et al., 2016 [101]; Mudimu et al., 2014 [102]; Najdenski et al., 2013 [97] |
| 2. | Chlorella sp. | Chlorellin | Gram-positive (G+) and Gram-negative (G−) bacteria | Pratt et al., 1944 [96] |
| 3. | Fischerella ambigua | Parsiguine | S. epidermidis PTCC 1114 and C. krusei ATCC 44507 | Najdenski et al., 2013 [97] |
| 4. | Chlorococcum strain HS-101, Dunaliella primolecta, Phaeodactylum tricornutum | Eicosapentaenoic acid (EPA), hexadecatrienoic acid (HTA) and palmitoleic acid (PA) | Methicillin-resistant S. aureus (MRSA), B. subtilis, Bacillus cereus, S. aureus, Enterobacter aerogenes | Desbois et al., 2008 [103] |
| 5. | Euglena viridis | Organic solvent extracts | Edwardsiella tarda, Aeromonas hydrophila, Pseudomonas tarda Pseudomonas fluorescens, Pseudomonas aeruginosa, Vibrio alginolyticus putida, V. anguillarum, Vibrio alginolyticus, Vibrio anguillarum Vibrio harveyi, fluvialis, Vibrio parahaemolyticus | Das et al., 2005 [161] |
| 6. | Scenedesmus obliquus, Haematococcus pluvialis | Fatty acids | E. coli and S. aureus | Rodríguez-Meizoso et al., 2010 [104] |
| 7. | Marine microalgae | Decadienal | Human pathogens MRSA and Haemophilus influenza | Mostafa et al., 2012 [89]; Smith et al., 2010 [98] |
| 8. | Spirogyra grantiana and Oscillatoria sancta | Ethanolic and methanolic extract | E. coli, P. vulgaris and P. mirabilis | Prakash et al., 2011 [105] |
| 9. | Dunaliella salina and Pseudokirchneriella subcapitata | Methanolic extract | S. aureus, P. aeruginosa, Escherichia coli and Klebsiella sp. | Pane et al., 2015 [106] |
| 10. | Porphyridium aerugineum | Phycobiliproteins | S. aureus, B. subtilis, S. pyogenes | Shannon et al., 2016 [95] |
| Antifungal Activity | ||||
| S. No. | Microalgae species | Compound/Fraction | Targeted microorganism | References |
| 1. | Chaetoceros sp., Chlorella vulgaris, Haematococcus pluvialis, Porphyridium purpureum, Rhodella reticulata, Scenedesmus quadricauda | Polysaccharides, organic solvent extracts, pigments and lipid fractions | A. fumigatus, A. niger, Penicillium sp., C. albicans, C. neoformans, S. cerevisiae, Microsporum sp., E. floccosum and T. mentagrophytes | Gueho et al., 1977 [123]; Mudimu et al., 2014 [102]; Washida et al., 2006 [122]; |
| 2. | Gambierdiscus toxicus, Thalassiothrix frauenfeldii | Gambieric acid | Penicillium, Aspergillus oryzae, Penicillium chrysogenum citrinum, Variotii paecilomyces, T. Mentagrophytes | Walter and Mahesh 2000 [124] |
| 3. | Chlorococcum humicola, Porphyridium aerugineum | Beta-carotene, chlorophyll-a and b | C. albicans, A. flavus and A. niger | Bhagavathy et al., 2011 [162] |
| 4. | Amphidinium klebsii | Polyols: karatungiols A(1) | A. niger | Ghannoum et al., 1999 [125] |
| 5. | Haslea. karadagensis | Pigments | Corollospora maritima, Dendryphiella salina and Lulworthia sp. | Gastineau et al., 2012 [126] |
| 6. | Chaetoceros lauderi, Chlorella vulgaris and Gambierdiscus toxicus | Organic extracts | Aspergillus fumigatus | Ghasemi et al., 2007 [127] |
| 7. | Scenedesmus quadricauda | Organic solvent extracts | C. albicans, S. cerevisiae, A. flavus, A. niger, P. herquei, A. brassicae, F. moniliforme, Helminthosporium sp. | Alangaden 2011 [119] |
| Antiviral Activity | ||||
| S. No. | Microalgae species | Compound/Fraction | Targeted virus | References |
| 1. | Gelidium cartilagenium | Polysaccharides | Influenza B and mumps viruses | Gerber et al., 1950 |
| 2. | Dunaliella sp., Spirulina platensis | Acyclovir®, Spirulan | Type 1 Herpes simplex virus (HSV-1) and Type 1 Human Immunodeficiency Virus (HIV-1) | Hayashi et al., 1996 [111]; Raposo et al., 2014 [74] |
| 3. | Arthrospira platensis, Cochlodinium polykrikoides, Gymnodinium impudicum Porphyridium sp., and Rhodomonas reticulate | Polysaccharides | Varicella zoster viruses (VZV), human cytomegalovirus (HCMV), murine leukemia virus (MuLV), Flu-A viruses, Hepatitis B virus (HBV); viral hemorrhagic septicemia virus (VHSV); African swine fever virus (ASFV), vaccinia virus (VACV), vesicular stomatitis virus (VSV), Encephalomyocarditis virus, Flu-A and Flu-B, respiratory syncytial virus types A (RSV-A) and B (RSV-B), measles, mumps | Falaise et al., 2016 [101]; Huleihel et al., 2002 [114]; Raposo et al., 2014 [74]; Santoyo et al., 2011 [113]; Silva et al., 2018 [112]; Talyshinsky et al., 2002 [115] |
| 4. | Gyrodinium impudicum | Sulphated polysaccharide | Encephalomyocarditis RNA virus (EMCV) | Huheihel et al., 2002 [114]; Kim et al., 2012 [80] |
| 5. | Arthrospira platensis, Porphyridium. purpureum | Polysaccharide TK-V3 Exopolysaccharides | Vaccinia and Ectromelia orthopoxvirus | Radonic et al., 2010 [116] |
| 6. | Cochlodinium polykrikoides Navicula directa | Naviculan | HIV-1,HSV-1 and influenza virus type A (IFV-A) | Lee et al., 2006 [117] |
| 7. | Chlorella autotrophica, Dunaliella tertiolecta, Ellipsoidon sp., Isochrysis galbana var. Tiso and Porphyridium cruentum | Endocellular extracts | Inhibited the viral infection of epithelioma papulosum cyprinid (EPC) cells | Graf et al., 2018 [118]; Talyshinsky et al., 2002 [115] |
| 8. | Dunaliella salina, Haematococcus pluvialis | Short chain fatty acids, β-ionone, phytol, neophytadiene, palmitic and α-linolenic acids | Herpes simplex virus type 1 (HSV-1) | Santoyo et al., 2011 [113] |
| 9. | Cochlodiniumpolykrikoides | Extracellular sulphated polysaccharides: A1 and A2 | Flu-A and B-Flu, Syncytial breathing Forms of Viruses A (RSV-A) A, and B (RSV-B), HIV-1, HSV-1, Parainfluenza type 2(PFluV-2) | Hasui et al., 1995 [163]; Morokutti-Kurz et al., 2017 [72]; |
| Antioxidant Activity | ||||
| S. No. | Microalgae species | Compound/Fraction | Application | References |
| 1. | Anabaena flos aquae, Anabaena oryzae, Nostoc humifusum, Chlorella vulgaris, Nostoc muscorum, Oscillatoria sp., Phormedium fragile, Spirulina platensis, and Wollea saccata | Algal extracts | Antioxidant activity is higher compared to standard Butylated hydroxytoluene (BHT) antioxidant | Barkia et al., 2019 [12]; Kattappagari et al., 2015 [87]; Lawrence et al., 2018 [84]; Shanab et al., 2012 [90] |
| 2. | Chlorella sorokiniana Sanropus androgynous | β-carotene, ά tocopherol, Chlorophyll and lutein | High radical scavenging activity | de Morais et al., 2015 [46]; Matsukawa et al., 2000 [92]; Suparmi et al., 2016 [91]; |
| 3. | Porphyridium cruentum | Exopolysaccharides | Scavenging hydroxyl, superoxide anion, and DPPH free radicals | Li et al., 2007 [164]; Wu et al., 2014 [68] |
| 4. | Dunaliella sp., Rhodella reticulata | Glutathione, Sulfated polysaccharides | Scavenges ROS and prevents cellular damage | de Jesus Raposo et al., 2013 [94]; Li et al., 2007 [164]; Mostafa 2012 [89]; Raposo et al., 2014 [74] |
| Anticancer Activity | ||||
| S. No. | Microalgae species | Compound/Fraction | Application | References |
| 1. | Nostoc muscorum | Phycobilins, phenolic compounds and polysaccharides | Anticancer activity against cell lines of Ehrlich Ascites Carcinoma and Human hepatocellular cancer | Shanab et al., 2012 [90]; Sheih et al., 2010 [136] |
| 2. | Red algae | c-phycocyanin | Anti-proliferation | Kim et al., 2011 [83] |
| 3. | G. impudicum | sulphate polysaccharide (p-KG03) | Prevents tumor cell growth | Wang et al., 2007 [132] |
| 4. | Chlorella vulgaris | β-(1,3)-glucan | Antitumor agent | Delasoie et al., 2018 [86]; Laroche and Michaud 2007 [133] |
| 5. | Porphyridium sp. | Sulphated polysaccharides | Tumor cell inhibition and proliferation of colon cancer in rats | Geresh et al. 2002 [135]; Sun et al. 2012 [134] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiran, B.R.; Venkata Mohan, S. Microalgal Cell Biofactory—Therapeutic, Nutraceutical and Functional Food Applications. Plants 2021, 10, 836. https://doi.org/10.3390/plants10050836
Kiran BR, Venkata Mohan S. Microalgal Cell Biofactory—Therapeutic, Nutraceutical and Functional Food Applications. Plants. 2021; 10(5):836. https://doi.org/10.3390/plants10050836
Chicago/Turabian StyleKiran, Boda Ravi, and S. Venkata Mohan. 2021. "Microalgal Cell Biofactory—Therapeutic, Nutraceutical and Functional Food Applications" Plants 10, no. 5: 836. https://doi.org/10.3390/plants10050836
APA StyleKiran, B. R., & Venkata Mohan, S. (2021). Microalgal Cell Biofactory—Therapeutic, Nutraceutical and Functional Food Applications. Plants, 10(5), 836. https://doi.org/10.3390/plants10050836