
Tissue-Specific RNA-Seq Analysis and Identification of Receptor-Like Proteins Related to Plant Growth in Capsicum annuum


Abstract
:1. Introduction
2. Results
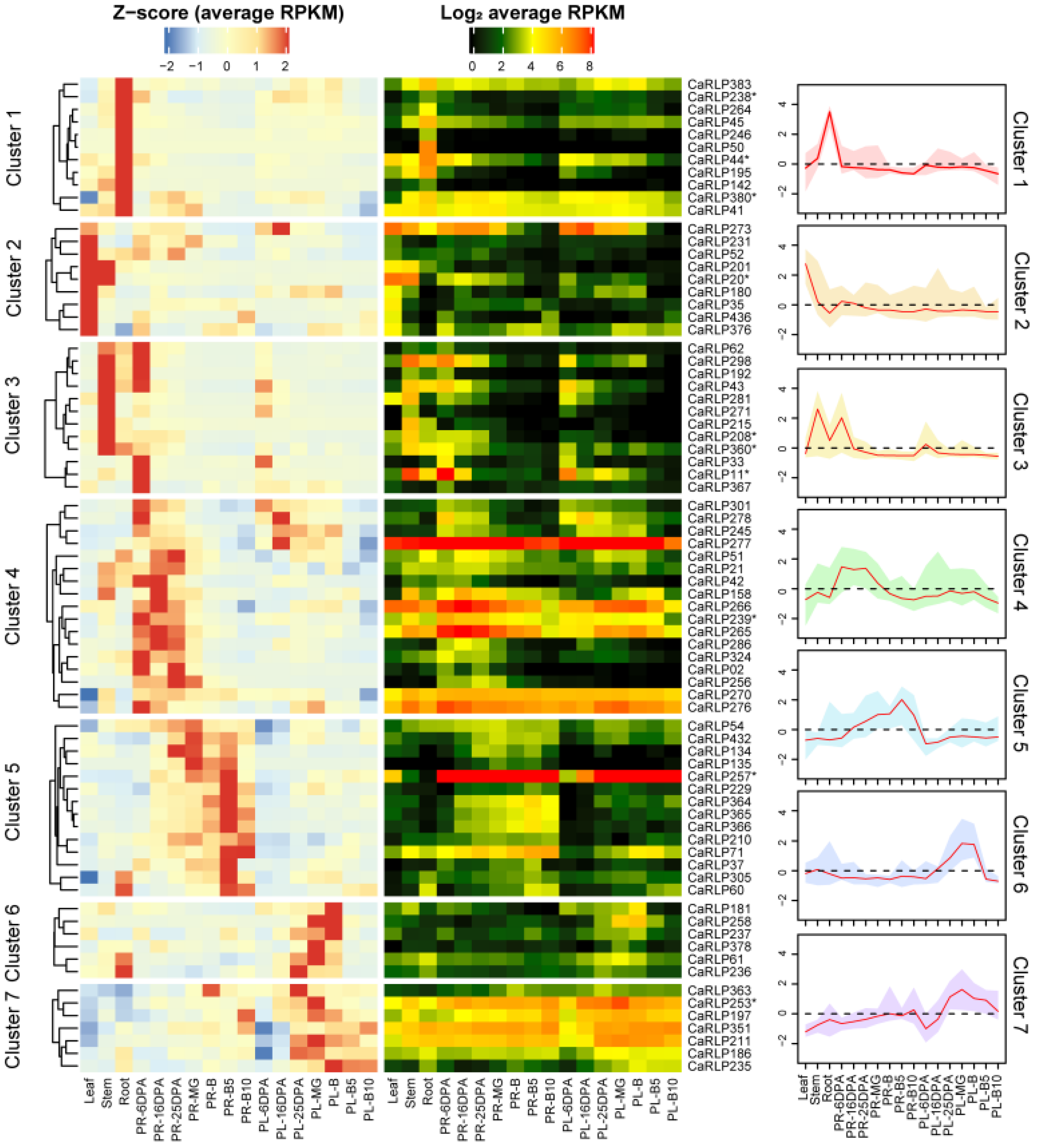
2.1. CaRLP Gene Expression Analysis in Various Organs and across Fruit Developmental Stages
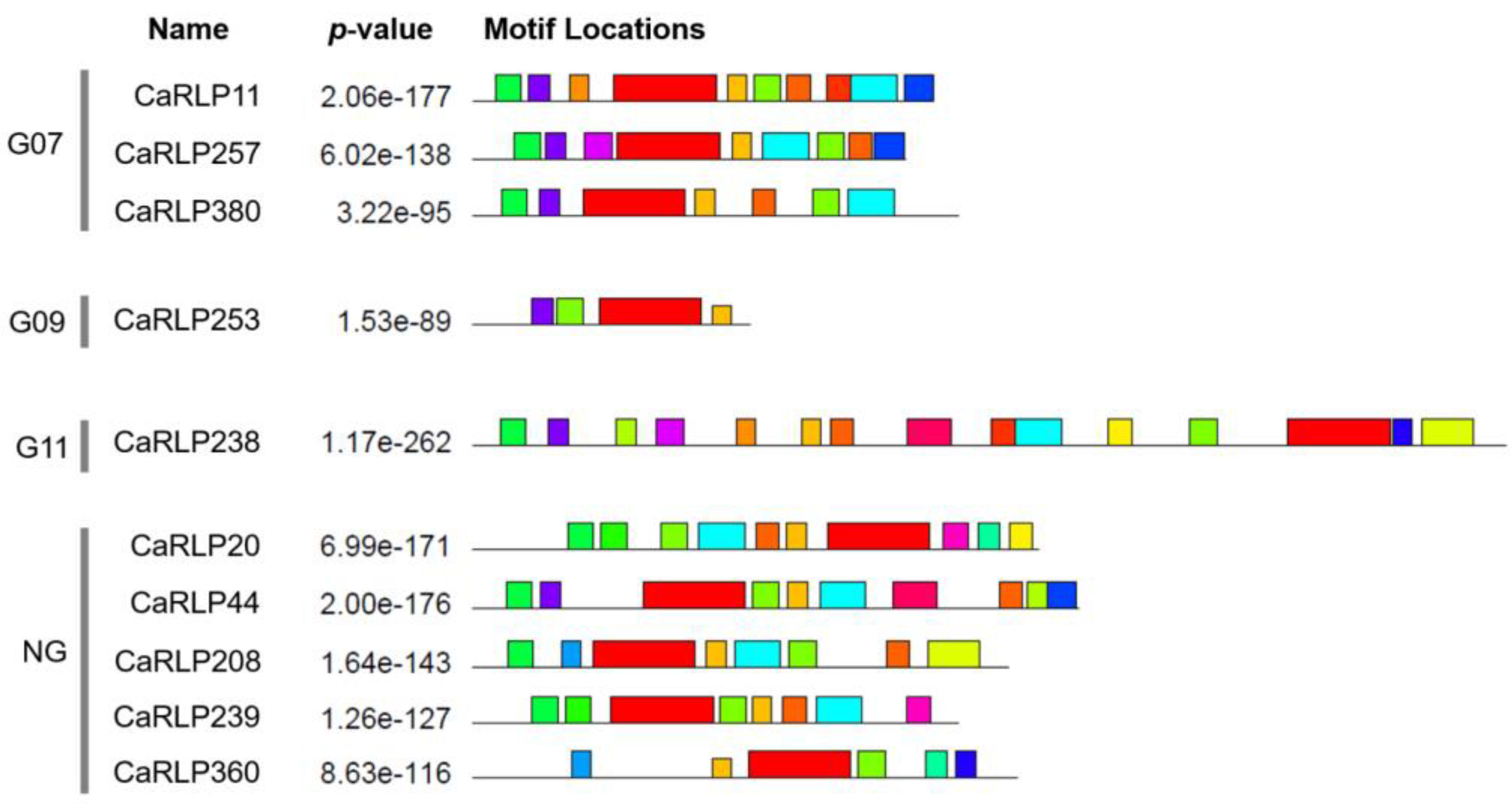
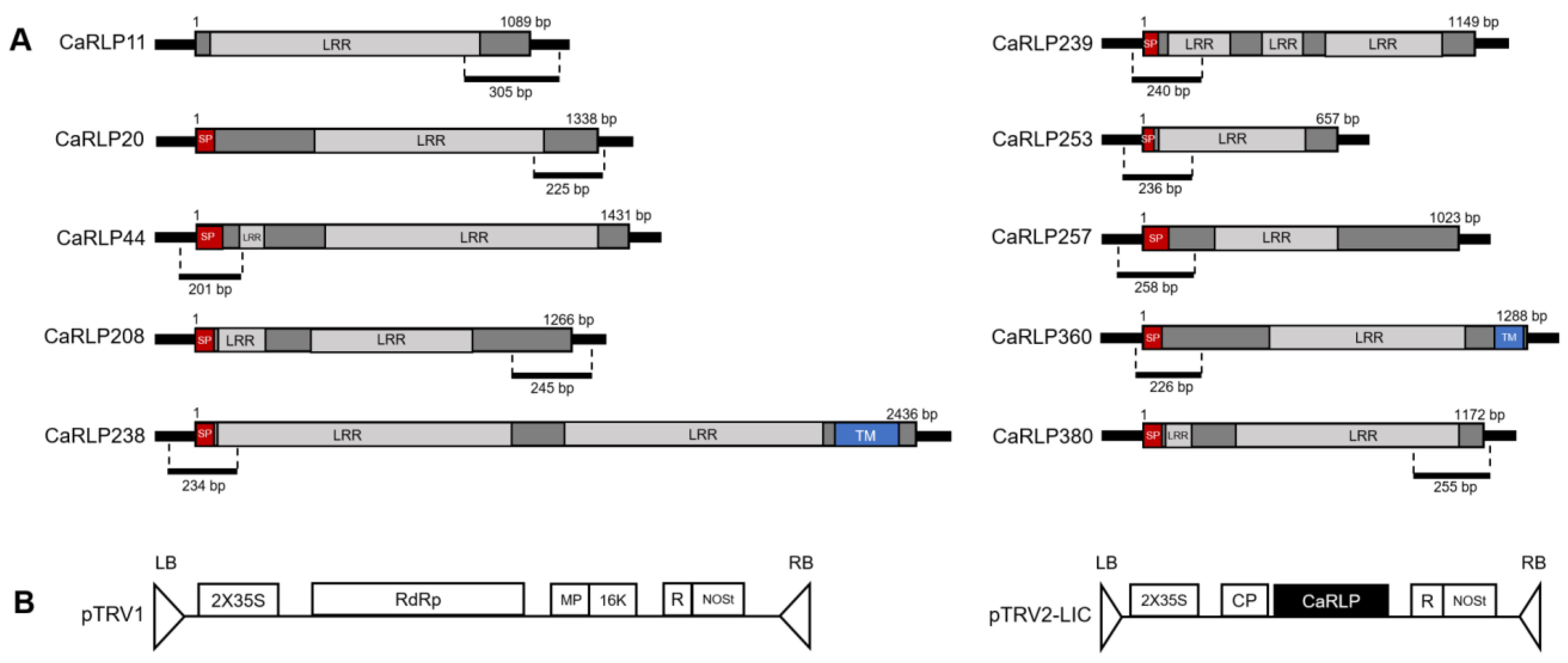
2.2. Structure Analysis of Selected CaRLPs
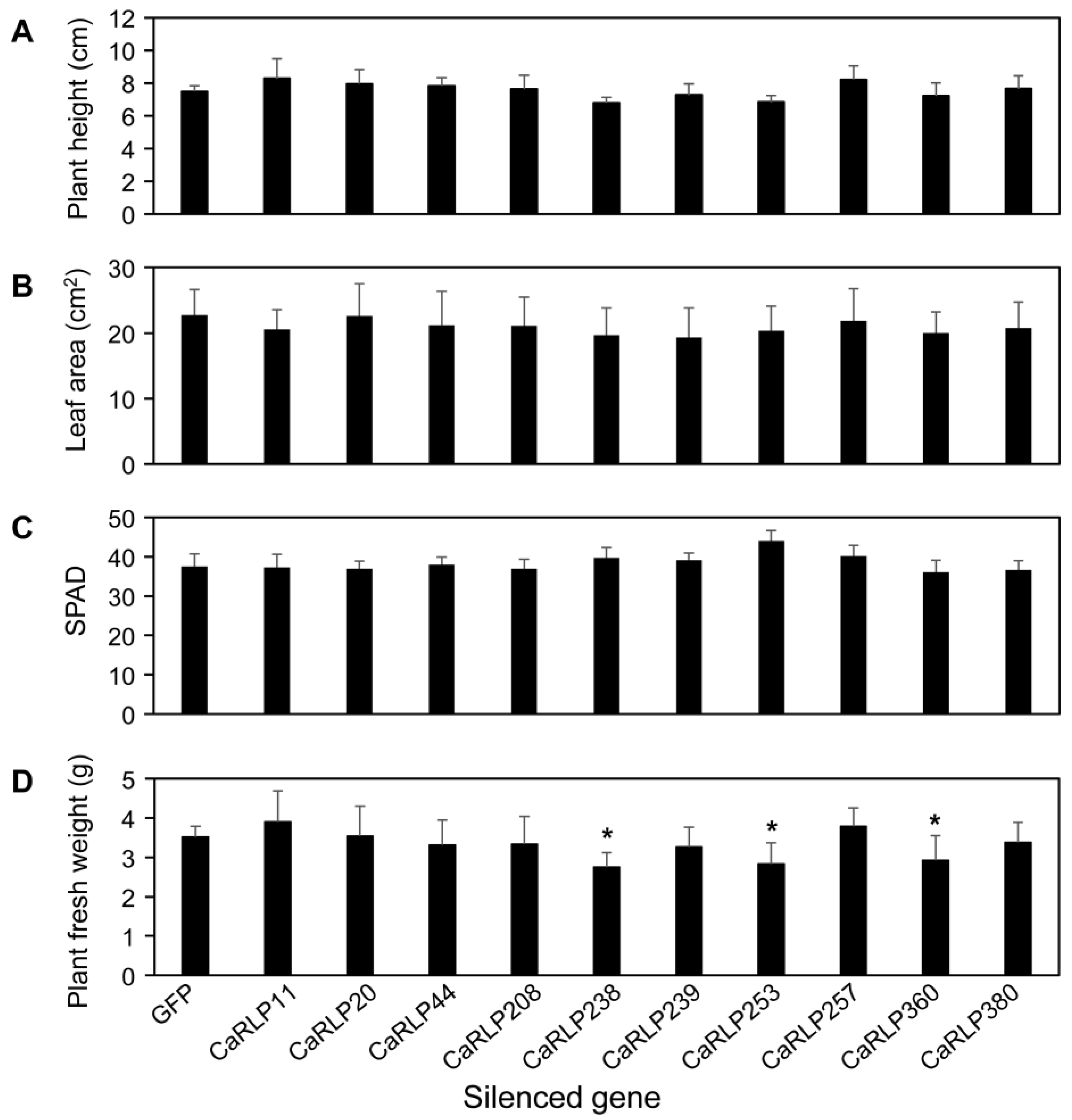
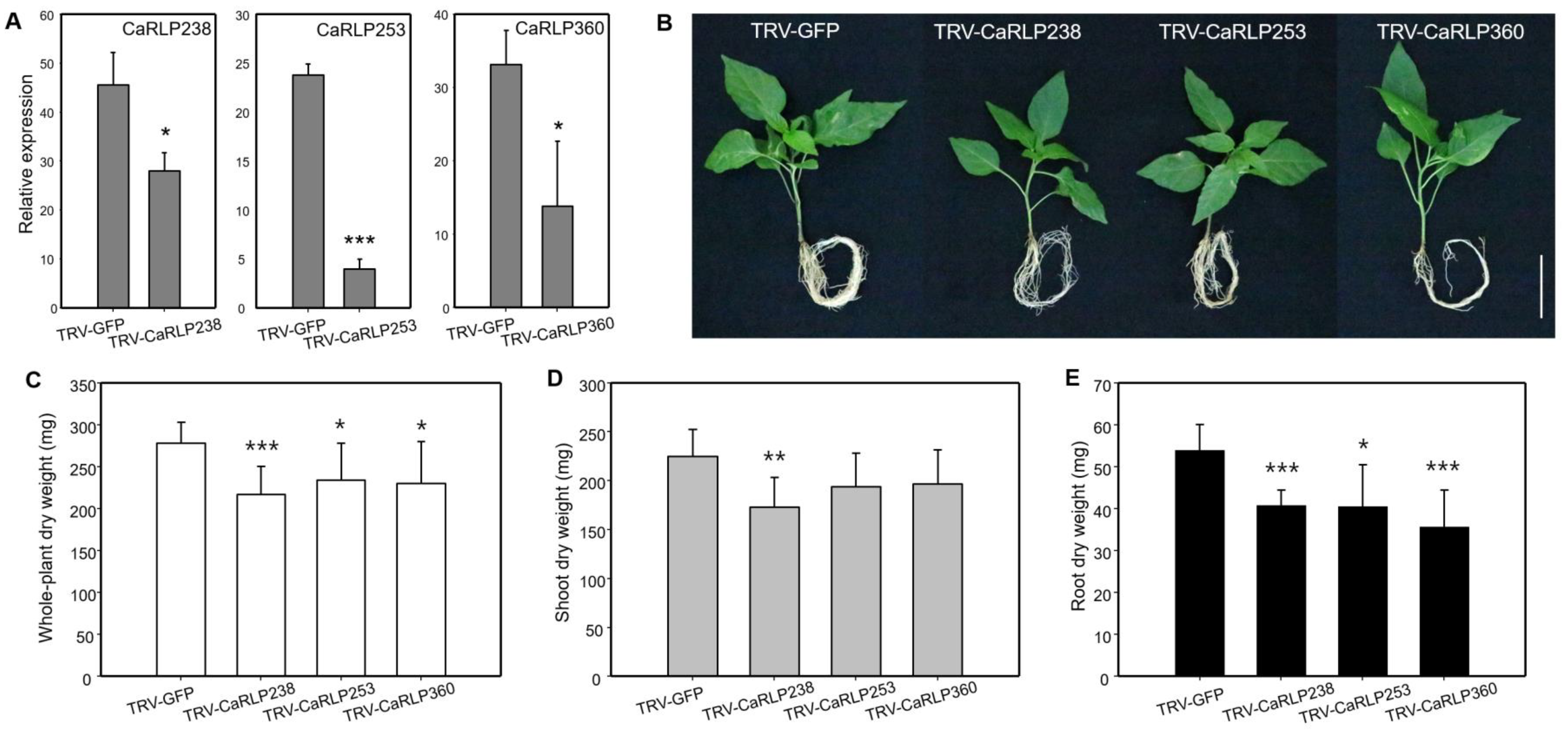
2.3. Phenotypic Alterations in Root Growth of CaRLP-Silenced Plants
3. Discussion
4. Materials and Methods
4.1. CaRLP Information, RNA-Seq Data, and Expression Profiling
4.2. Motif Analysis and Structure Analysis of CaRLP Genes
4.3. Gene Cloning for Virus-Induced Gene Silencing (VIGS) Assay
4.4. Semi-Quantitative RT-PCR Analysis
4.5. Phenotypic Analysis of CaRLP-Silenced Pepper Plants
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- He, Y.; Zhou, J.; Shan, L.; Meng, X. Plant cell surface receptor-mediated signaling—A common theme amid diversity. J. Cell Sci. 2018, 131, jcs209353. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.; Lv, S.; Yang, N.; Lv, Y.; Liu, Z.; Wu, J.; Wang, G. The multifunction of CLAVATA2 in plant development and immunity. Front. Plant Sci. 2016, 7, 1573. [Google Scholar] [CrossRef] [Green Version]
- Bohm, H.; Albert, I.; Fan, L.; Reinhard, A.; Nurnberger, T. Immune receptor complexes at the plant cell surface. Curr. Opin. Plant Biol. 2014, 20, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.A.; Thomas, C.M.; Hammond-Kosack, K.E.; Balint-Kurti, P.J.; Jones, J.D. Isolation of the tomato Cf-9 gene for resistance to Cladosporium fulvum by transposon tagging. Science 1994, 266, 789–793. [Google Scholar] [CrossRef]
- Dixon, M.S.; Jones, D.A.; Keddie, J.S.; Thomas, C.M.; Harrison, K.; Jones, J.D. The tomato Cf-2 disease resistance locus comprises two functional genes encoding leucine-rich repeat proteins. Cell 1996, 84, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Dixon, M.S.; Hatzixanthis, K.; Jones, D.A.; Harrison, K.; Jones, J.D. The tomato Cf-5 disease resistance gene and Six homologs show pronounced allelic variation in leucine-rich repeat copy number. Plant Cell 1998, 10, 1915–1925. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.M.; Jones, D.A.; Parniske, M.; Harrison, K.; Balint-Kurti, P.J.; Hatzixanthis, K.; Jones, J.D. Characterization of the tomato Cf-4 gene for resistance to Cladosporium fulvum identifies sequences that determine recognitional specificity in Cf-4 and Cf-9. Plant Cell 1997, 9, 2209–2224. [Google Scholar] [CrossRef] [Green Version]
- Takken, F.L.; Thomas, C.M.; Joosten, M.H.; Golstein, C.; Westerink, N.; Hille, J.; Nijkamp, H.J.; De Wit, P.J.; Jones, J.D. A second gene at the tomato Cf-4 locus confers resistance to cladosporium fulvum through recognition of a novel avirulence determinant. Plant J. 1999, 20, 279–288. [Google Scholar] [CrossRef]
- Ron, M.; Avni, A. The receptor for the fungal elicitor ethylene-inducing xylanase is a member of a resistance-like gene family in tomato. Plant Cell 2004, 16, 1604–1615. [Google Scholar] [CrossRef] [Green Version]
- Kawchuk, L.M.; Hachey, J.; Lynch, D.R.; Kulcsar, F.; van Rooijen, G.; Waterer, D.R.; Robertson, A.; Kokko, E.; Byers, R.; Howard, R.J.; et al. Tomato Ve disease resistance genes encode cell surface-like receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 6511–6515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegenauer, V.; Furst, U.; Kaiser, B.; Smoker, M.; Zipfel, C.; Felix, G.; Stahl, M.; Albert, M. Detection of the plant parasite Cuscuta reflexa by a tomato cell surface receptor. Science 2016, 353, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Jehle, A.K.; Lipschis, M.; Albert, M.; Fallahzadeh-Mamaghani, V.; Furst, U.; Mueller, K.; Felix, G. The receptor-like protein ReMAX of Arabidopsis detects the microbe-associated molecular pattern eMax from Xanthomonas. Plant Cell 2013, 25, 2330–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Diener, A.C. Arabidopsis thaliana resistance to fusarium oxysporum 2 implicates tyrosine-sulfated peptide signaling in susceptibility and resistance to root infection. PLoS Genet. 2013, 9, e1003525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albert, I.; Bohm, H.; Albert, M.; Feiler, C.E.; Imkampe, J.; Wallmeroth, N.; Brancato, C.; Raaymakers, T.M.; Oome, S.; Zhang, H.Q.; et al. An RLP23-SOBIR1-BAK1 complex mediates NLP-triggered immunity. Nat. Plants 2015, 1, 15140. [Google Scholar] [CrossRef] [PubMed]
- Larkan, N.J.; Lydiate, D.J.; Parkin, I.A.; Nelson, M.N.; Epp, D.J.; Cowling, W.A.; Rimmer, S.R.; Borhan, M.H. The Brassica napus blackleg resistance gene LepR3 encodes a receptor-like protein triggered by the Leptosphaeria maculans effector AVRLM1. New Phytol. 2013, 197, 595–605. [Google Scholar] [CrossRef]
- Vinatzer, B.A.; Patocchi, A.; Gianfranceschi, L.; Tartarini, S.; Zhang, H.B.; Gessler, C.; Sansavini, S. Apple contains receptor-like genes homologous to the Cladosporium fulvum resistance gene family of tomato with a cluster of genes cosegregating with Vf apple scab resistance. Mol. Plant Microbe Interact. 2001, 14, 508–515. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Verzaux, E.; Chaparro-Garcia, A.; Bijsterbosch, G.; Keizer, L.C.P.; Zhou, J.; Liebrand, T.W.H.; Xie, C.H.; Govers, F.; Robatzek, S.; et al. Elicitin recognition confers enhanced resistance to Phytophthora infestans in potato. Nat. Plants 2015, 1, 15034. [Google Scholar] [CrossRef] [PubMed]
- Nadeau, J.A.; Sack, F.D. Control of stomatal distribution on the Arabidopsis leaf surface. Science 2002, 296, 1697–1700. [Google Scholar] [CrossRef]
- Jeong, S.; Trotochaud, A.E.; Clark, S.E. The Arabidopsis CLAVATA2 gene encodes a receptor-like protein required for the stability of the CLAVATA1 receptor-like kinase. Plant Cell 1999, 11, 1925–1934. [Google Scholar] [CrossRef] [Green Version]
- Fiers, M.; Golemiec, E.; Xu, J.; van der Geest, L.; Heidstra, R.; Stiekema, W.; Liu, C.M. The 14-amino acid CLV3, CLE19, and CLE40 peptides trigger consumption of the root meristem in Arabidopsis through a CLAVATA2-dependent pathway. Plant Cell 2005, 17, 2542–2553. [Google Scholar] [CrossRef] [Green Version]
- Fiers, M.; Hause, G.; Boutilier, K.; Casamitjana-Martinez, E.; Weijers, D.; Offringa, R.; van der Geest, L.; van Lookeren Campagne, M.; Liu, C.M. Mis-expression of the CLV3/ESR-like gene CLE19 in Arabidopsis leads to a consumption of root meristem. Gene 2004, 327, 37–49. [Google Scholar] [CrossRef]
- Strabala, T.J.; O’Donnell, P.J.; Smit, A.M.; Ampomah-Dwamena, C.; Martin, E.J.; Netzler, N.; Nieuwenhuizen, N.J.; Quinn, B.D.; Foote, H.C.; Hudson, K.R. Gain-of-function phenotypes of many CLAVATA3/ESR genes, including four new family members, correlate with tandem variations in the conserved CLAVATA3/ESR domain. Plant Physiol. 2006, 140, 1331–1344. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Feldman, L.J. CLE14/CLE20 peptides may interact with CLAVATA2/CORYNE receptor-like kinases to irreversibly inhibit cell division in the root meristem of Arabidopsis. Planta 2010, 232, 1061–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.D.; Ellendorff, U.; Kemp, B.; Mansfield, J.W.; Forsyth, A.; Mitchell, K.; Bastas, K.; Liu, C.M.; Woods-Tor, A.; Zipfel, C.; et al. A genome-wide functional investigation into the roles of receptor-like proteins in Arabidopsis. Plant Physiol. 2008, 147, 503–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taguchi-Shiobara, F.; Yuan, Z.; Hake, S.; Jackson, D. The fasciated ear2 gene encodes a leucine-rich repeat receptor-like protein that regulates shoot meristem proliferation in maize. Genes Dev. 2001, 15, 2755–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, W.H.; Lee, J.; Koo, N.; Kwon, J.S.; Park, B.; Kim, Y.M.; Yeom, S.I. Universal gene co-expression network reveals receptor-like protein genes conferring broad-spectrum resistance in pepper (Capsicum annuum L.). bioRxiv 2021. [Google Scholar] [CrossRef]
- Kang, W.H.; Kim, S.; Lee, H.A.; Choi, D.; Yeom, S.I. Genome-wide analysis of Dof transcription factors reveals functional characteristics during development and response to biotic stresses in pepper. Sci. Rep. 2016, 6, 33332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamieson, P.A.; Shan, L.B.; He, P. Plant cell surface molecular cypher: Receptor-like proteins and their roles in immunity and development. Plant Sci. 2018, 274, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Fritz-Laylin, L.K.; Krishnamurthy, N.; Tor, M.; Sjolander, K.V.; Jones, J.D. Phylogenomic analysis of the receptor-like proteins of rice and Arabidopsis. Plant Physiol. 2005, 138, 611–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, W.H.; Yeom, S.I. Genome-wide identification, classification, and expression analysis of the receptor-like protein family in tomato. Plant Pathol. J. 2018, 34, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Petre, B.; Hacquard, S.; Duplessis, S.; Rouhier, N. Genome analysis of poplar LRR-RLP gene clusters reveals RISP, a defense-related gene coding a candidate endogenous peptide elicitor. Front. Plant Sci. 2014, 5, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restrepo-Montoya, D.; Brueggeman, R.; McClean, P.E.; Osorno, J.M. Computational identification of receptor-like kinases “RLK” and receptor-like proteins “RLP” in legumes. BMC Genom. 2020, 21, 459. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Bayer, P.E.; Tirnaz, S.; Edwards, D.; Batley, J. Genome-wide identification and evolution of receptor-like kinases (RLKs) and receptor like proteins (RLPs) in Brassica juncea. Biology 2020, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Starr, T.N.; Picton, L.K.; Thornton, J.W. Alternative evolutionary histories in the sequence space of an ancient protein. Nature 2017, 549, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Gabaldon, T.; Koonin, E.V. Functional and evolutionary implications of gene orthology. Nat. Rev. Genet. 2013, 14, 360–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, B.Q.; Xue, J.; Zhang, N.N.; Xu, L.H.; Yao, X.R.; Yang, Q.J.; Yu, Y.; Wang, H.B.; Zhang, D.D.; Li, J.F. Rice chitin receptor OsCEBiP is not a transmembrane protein but targets the plasma membrane via a GPI Anchor. Mol. Plant 2017, 10, 767–770. [Google Scholar] [CrossRef]
- Kellogg, E.A. Evolution of developmental traits. Curr. Opin. Plant Biol. 2004, 7, 92–98. [Google Scholar] [CrossRef]
- Burch-Smith, T.M.; Anderson, J.C.; Martin, G.B.; Dinesh-Kumar, S.P. Applications and advantages of virus-induced gene silencing for gene function studies in plants. Plant J. 2004, 39, 734–746. [Google Scholar] [CrossRef]
- Lange, M.; Yellina, A.L.; Orashakova, S.; Becker, A. Virus-induced gene silencing (VIGS) in plants: An overview of target species and the virus-derived vector systems. Methods Mol. Biol. 2013, 975, 1–14. [Google Scholar] [CrossRef]
- Wege, S.; Scholz, A.; Gleissberg, S.; Becker, A. Highly efficient virus-induced gene silencing (VIGS) in California poppy (Eschscholzia californica): An evaluation of VIGS as a strategy to obtain functional data from non-model plants. Ann. Bot. 2007, 100, 641–649. [Google Scholar] [CrossRef] [Green Version]
- Raz, A.; Dahan-Meir, T.; Melamed-Bessudo, C.; Leshkowitz, D.; Levy, A.A. Redistribution of meiotic crossovers along wheat chromosomes by virus-induced gene silencing. Front. Plant Sci. 2020, 11, 635139. [Google Scholar] [CrossRef]
- Kim, S.; Park, M.; Yeom, S.I.; Kim, Y.M.; Lee, J.M.; Lee, H.A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.T.; et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Nam, J.Y.; Jang, H.; Kim, N.; Kim, Y.M.; Kang, W.H.; Yeom, S.I. Comprehensive transcriptome resource for response to phytohormone-induced signaling in Capsicum annuum L. BMC Res. Notes 2020, 13, 40. [Google Scholar] [CrossRef]
- Kang, W.H.; Sim, Y.M.; Koo, N.; Nam, J.Y.; Lee, J.; Kim, N.; Jang, H.; Kim, Y.M.; Yeom, S.I. Transcriptome profiling of abiotic responses to heat, cold, salt, and osmotic stress of Capsicum annuum L. Sci. Data 2020, 7, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, W.H.; Lee, J.; Nam, J.Y.; Yeom, S.I. Identification of CaLOP regulating development and growth through virus-induced gene silencing in pepper. Hortic. Sci. Technol. 2018, 36, 292–301. [Google Scholar] [CrossRef]
- Dong, Y.; Burch-Smith, T.M.; Liu, Y.; Mamillapalli, P.; Dinesh-Kumar, S.P. A ligation-independent cloning tobacco rattle virus vector for high-throughput virus-induced gene silencing identifies Roles for NbMADS4-1 and -2 in floral development. Plant Physiol. 2007, 145, 1161–1170. [Google Scholar] [CrossRef] [Green Version]
- Kwon, J.S.; Nam, J.Y.; Yeom, S.I.; Kang, W.H. Leaf-to-whole plant spread bioassay for pepper and Ralstonia solanacearum interaction determines inheritance of resistance to bacterial wilt for further breeding. Int. J. Mol. Sci. 2021, 22, 2279. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CaRLP ID | Location | CDS Size (bp) | Protein Size (aa) | Group |
|---|---|---|---|---|
| CaRLP11 | Ch1 | 1089 | 362 | G07 |
| CaRLP20 | Ch2 | 1338 | 445 | NG |
| CaRLP44 | Ch2 | 1431 | 476 | NG |
| CaRLP208 | Ch7 | 1266 | 421 | NG |
| CaRLP238 | Ch8 | 2436 | 811 | G11 |
| CaRLP239 | Ch8 | 1149 | 382 | NG |
| CaRLP253 | Ch8 | 657 | 218 | G09 |
| CaRLP257 | Ch9 | 1023 | 340 | G07 |
| CaRLP360 | Ch0 1 | 1288 | 428 | NG |
| CaRLP380 | Ch0 | 1172 | 382 | G07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, W.-H.; Park, B.; Lee, J.; Yeom, S.-I. Tissue-Specific RNA-Seq Analysis and Identification of Receptor-Like Proteins Related to Plant Growth in Capsicum annuum. Plants 2021, 10, 972. https://doi.org/10.3390/plants10050972
Kang W-H, Park B, Lee J, Yeom S-I. Tissue-Specific RNA-Seq Analysis and Identification of Receptor-Like Proteins Related to Plant Growth in Capsicum annuum. Plants. 2021; 10(5):972. https://doi.org/10.3390/plants10050972
Chicago/Turabian StyleKang, Won-Hee, Boseul Park, Junesung Lee, and Seon-In Yeom. 2021. "Tissue-Specific RNA-Seq Analysis and Identification of Receptor-Like Proteins Related to Plant Growth in Capsicum annuum" Plants 10, no. 5: 972. https://doi.org/10.3390/plants10050972
APA StyleKang, W. -H., Park, B., Lee, J., & Yeom, S. -I. (2021). Tissue-Specific RNA-Seq Analysis and Identification of Receptor-Like Proteins Related to Plant Growth in Capsicum annuum. Plants, 10(5), 972. https://doi.org/10.3390/plants10050972