
A Review of Chenopodium quinoa (Willd.) Diseases—An Updated Perspective


Abstract
:
1. Introduction
2. Downy Mildew of Quinoa
2.1. Nomenclature and Distribution

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
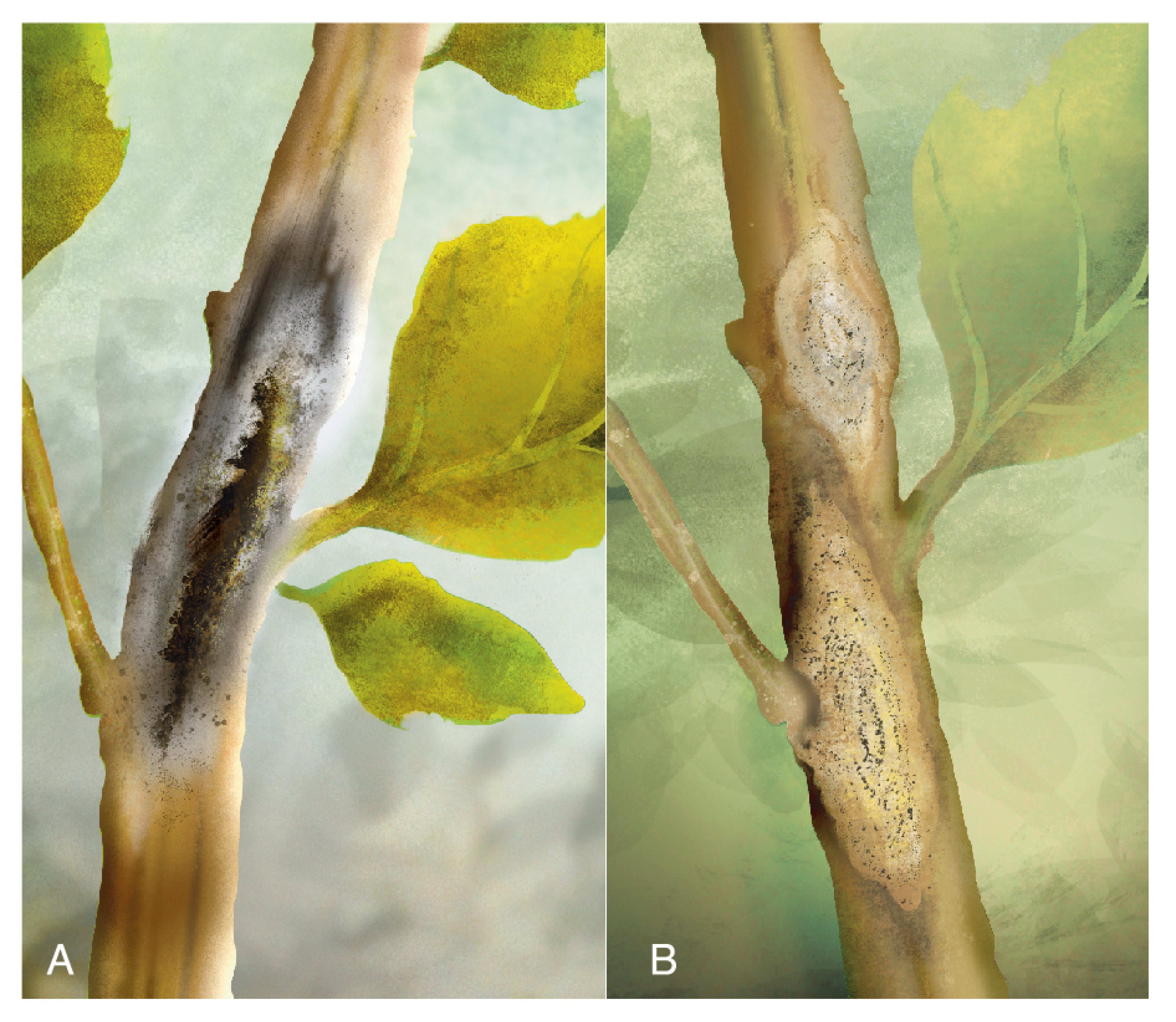{kind=link}
{kind=link}
{kind=link}
| Host (Genus/Species) | Pathogen Current Identity | Byford Classification (f. spp.) P. farinosa formae speciales |
|---|---|---|
| Beta spp. | P. schachtii [26] | P. farinosa f. sp. betae |
| C. álbum + C. quinoa | P. variabilis [29,30] | P. farinosa f. sp. chenopodii |
| Spinacia oleracea | P. effusa [33,34] | P. farinosa f. sp. spinaciae |
2.2. Infection Biology and Disease Symptoms
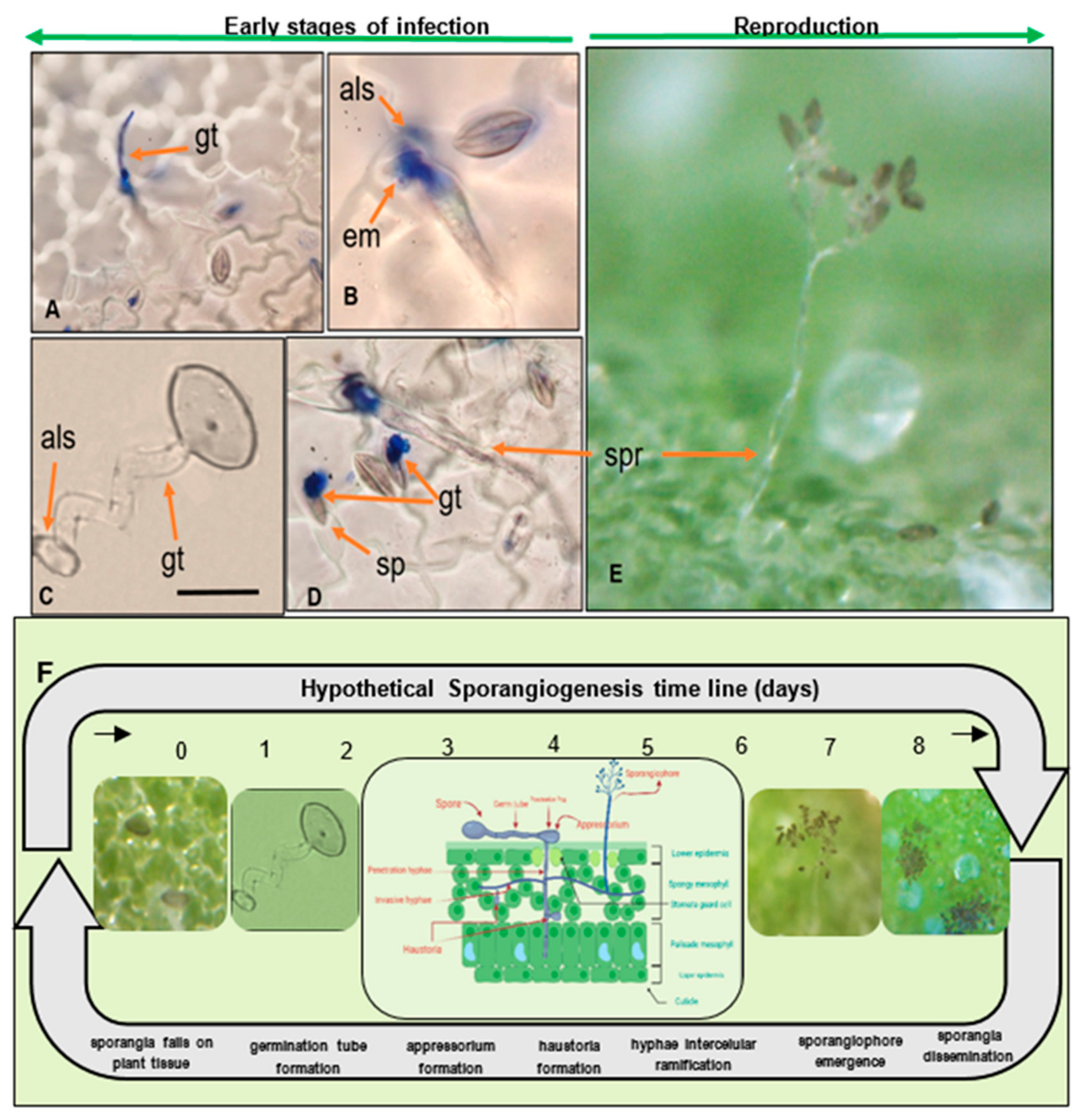
2.3. Morphology and Reproduction

2.4. Peronospora variabilis Genotypic Diversity and Virulence Profiling
- An area where quinoa cultivation was not regularly practised. The least virulent strains were present here and were identified as virulence group 2 (V2).
- A region where landraces and newly released cultivars were introduced. Only the most virulent strains belonging to group 4 (V4) were present here.
- Fields located where landraces and newly released cultivars have been cultivated for many years. Here, all four virulence groups were present.
2.5. Disease Assessment under Controlled Conditions and in the Field
- The phenological stage of the plants. Age-related resistance becomes relevant for biotrophic pathogens, which require healthy plant tissue to complete their cycle. [89,90]. The observation of symptoms should reflect the progression of the disease through periodical records, rather than observing its percentage of occurrence or incidence. For the quinoa/downy mildew interaction, it has been demonstrated that disease incidence has a low heritability H2 = 0.4 and a low correlation with severity and sporulation (0.67 and 0.65, respectively) [52]. Therefore, incidence or whole plant scores are unsuitable for this type of trial. To measure the area under the disease curve progression (AUDPC), a minimum record of three to four observations of disease severity is essential. A similar study has highlighted the importance of measuring the disease severity over time for other interactions, such as Phytophthora infestans infecting potatoes. The objective is to capture low, medium, and high infection levels in all the genotypes, including the susceptible ones [91].
- 2.
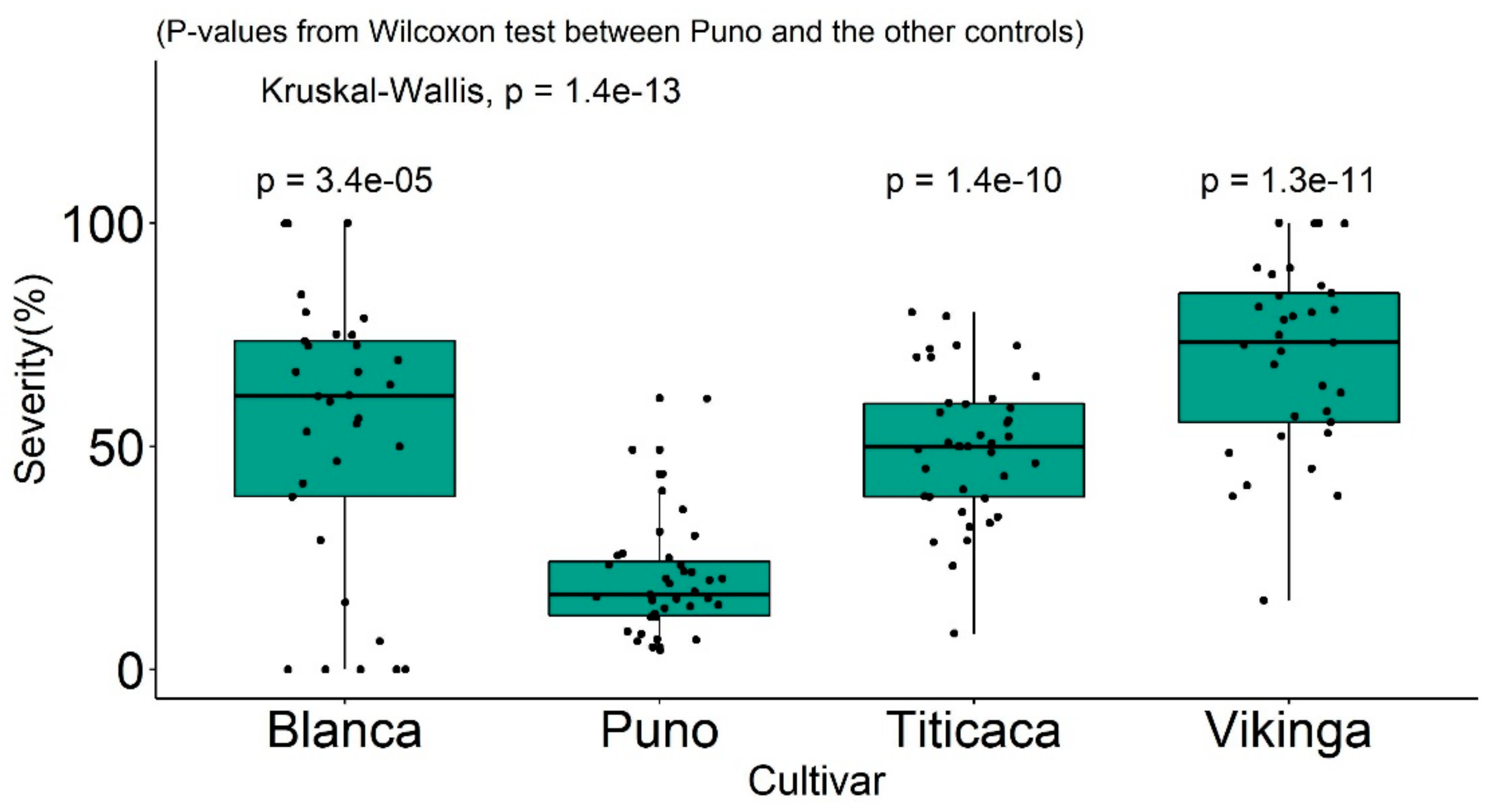
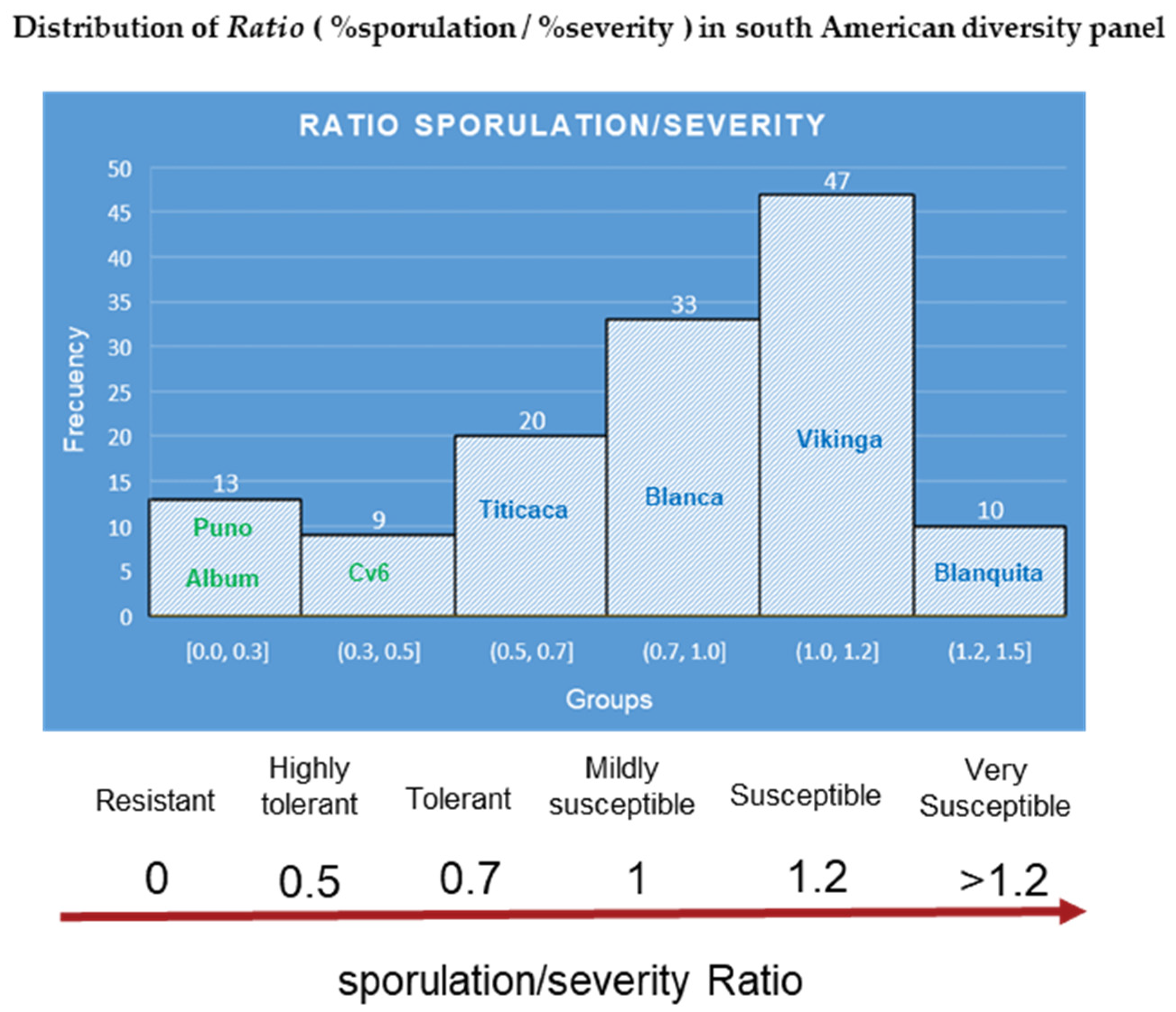
- The vegetative cycle of the plants. Late-maturing quinoa genotypes will display some degree of resistance [93] by increasing the latent period of the pathogen. Thus, readings for severity were taken ten days after infection instead of five in a recent study, in which a late cultivar Blanca was compared with the Danish cultivars Puno, Titicaca, and Vikinga [52] (Figure 7). Puno matures ten days later than Titicaca [94]. Cultivar Blanca is considered susceptible when additional time is given [52] (Figure 7). Vegetative cycle effects were also shown in another study that analyzed the mean-based cluster of inter-ecotype F2:6 population crosses and identified the following three clusters [48,95,96]:
- (a)
- Cluster one: consisting of late, mildew-resistant, high-yielding lines;
- (b)
- Cluster two: consisting of semi-late lines with intermediate yield and mildew susceptibility;
- (c)
- Cluster three: consisting of early to semi-late accessions with low yield and mildew susceptibility.
- 3.
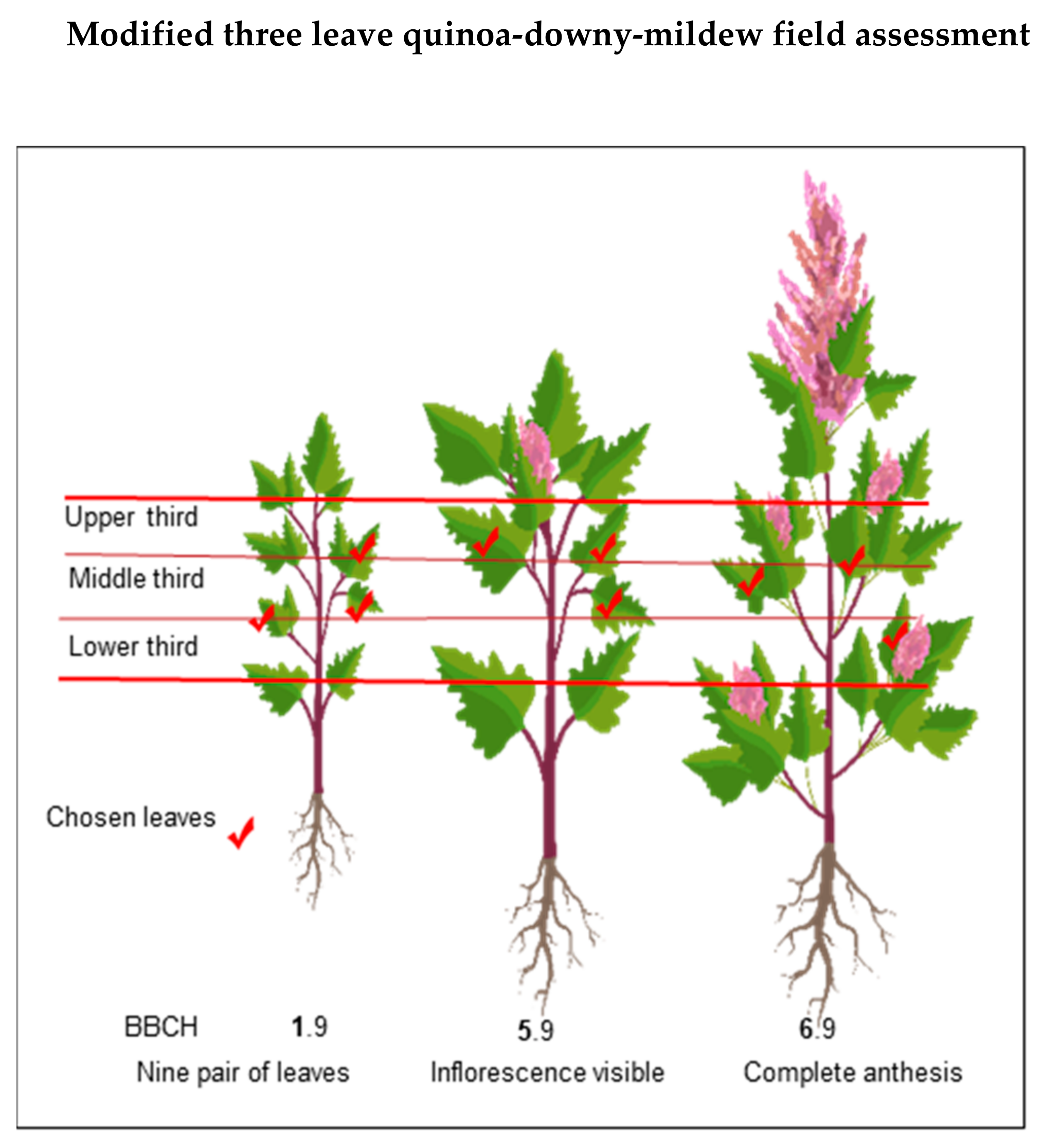
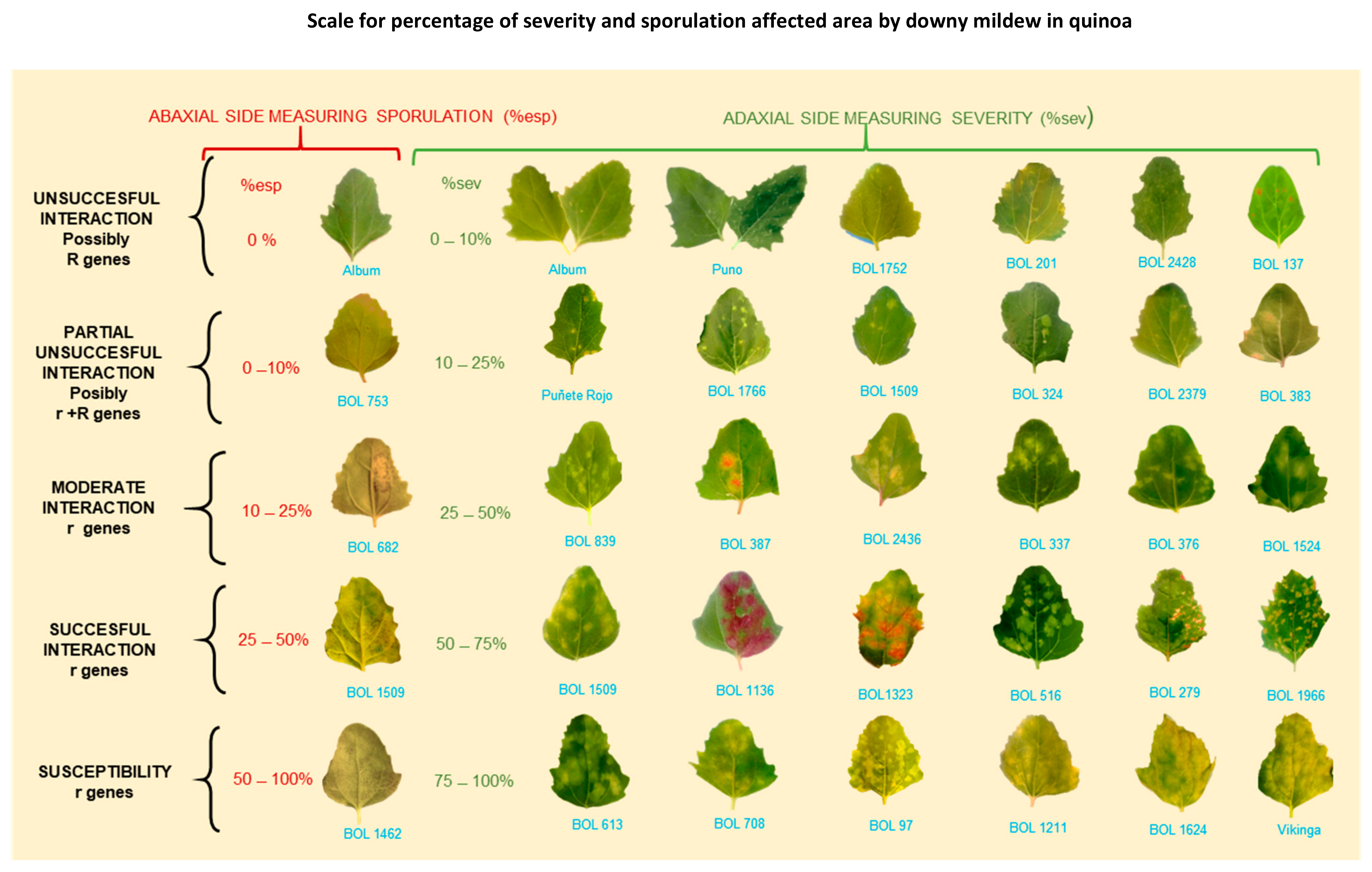
- Sampling method and sample size. Depending on the size of the experiment, there is no need to take severity readings in all the quinoa plants. Instead, consider the plot level and take readings on representative samples. Normally, 6–10 plants per plot are sufficient [54,91]. Next, an estimation of the percentage of affected foliage is required. Given the size of the plants and abundant foliage, it is not feasible to analyze the entire foliage; thus, it is recommended to perform scoring on individual leaves of the chosen plants [54]. Danielsen and Munk [97] evaluated various field assessment methods to predict yield losses due to downy mildew. The three-leaf method resulted in the highest negative correlation to yield (r = −0.736). Furthermore, disease progression relies on the successful infection of the host. It is often assumed that the susceptibility of host tissue is constant. However, in reality, it is a function of plant age and leaf position [14,98,99]. These responses might result from inducible plant defense responses, which occurs at the starting interaction site but also in distal, uninfected parts [99,100,101]. For these reasons, we suggest randomly choosing three leaves from the middle part (lower third, middle third, and upper third), as illustrated in Figure 8. Avoid the lower and upper extremities of the plant because they are prone to senescence/defoliation [97] and plant defense responses, respectively. Next, estimate the percentage of affected leaf area using the attached scale [79] (Figure 9). The average value from the score of the three leaves becomes the percentage of severity for each plant.

2.6. Yield Losses and Management
- tolerant crop varieties; and
- cultural practices (options on the list below).
- (a)
- Policymakers, smallholder farmers, and other stakeholders need resources for collective action for the establishment of a seed supply chain with quality standards (low levels of key seed-borne diseases). Experiences with complementary intervention such as capacity building and technical assistance have shown this influence in an appropriate conceptual model of sustainable production [104].
- (b)
- The detection of P. variabilis on the seed is achieved using a simple method [32,54]. In the case of the presence of an oospore, treat the seed with a systemic fungicide [105]. For small samples, alternative treatments such as a hot water bath (50 °C–60 °C) could be considered for 10 to 30 min, as this method has been applied successfully to eradicate seed-borne pathogens of spinach [106]. After or without treatment, the addition of beneficial microbes by priming the seed with products such as commercially available Trichoderma can enhance the growth of the plants [107].
- (c)
- Adjusting the space between rows and individuals, making the area less dense and increasing space between plants. In areas where the RH is as high as 80%, the minimum should be a 0.5-m space between rows and 0.15 m between plants [5].
- (d)
- Avoiding excess water in the field;
- (e)
- Implementing effective weed control, especially of alternate host C. album;
- (f)
- Practicing crop rotation;
- (g)
- Spraying the plants around 45 days after planting in areas with endemic infection as a preventive measure [69]. Use oomycete sensitive chemical control measures (e.g., Alietti) at principal growth stages, e.g., leaf development, inflorescence emergence, flowering, and fruit development [14,54,92]. Fungicides could be applied, alternating between systemic and contact products, starting with systemic products. Bio-pesticide or plant extracts could replace fungicides with a uniform and preventive application [5]. Inducers of resistance are an alternative [108].


2.7. Genetics of Resistance to Downy Mildew
3. Ascomycete Fungi
3.1. Fungi Identified by Molecular and Morphological Approaches
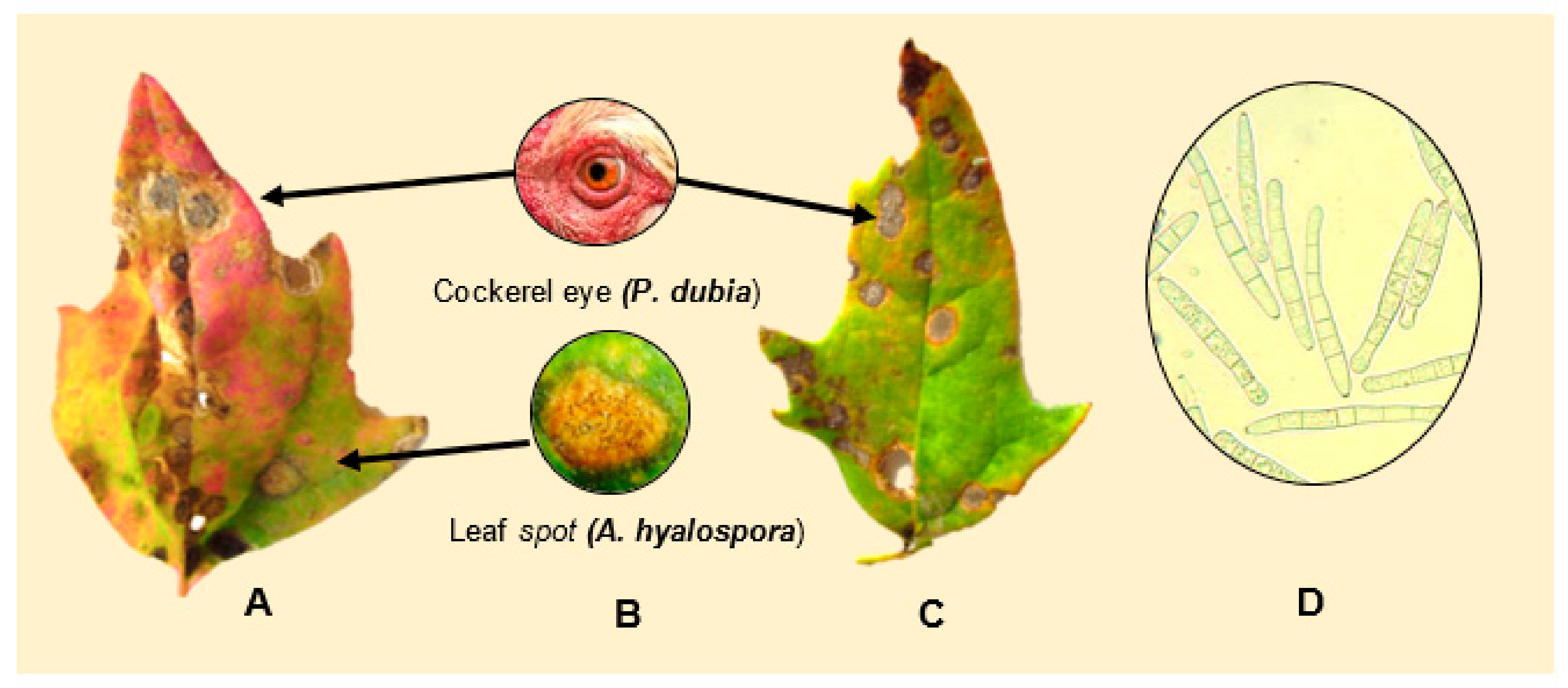
3.1.1. Ascochyta Leaf Spot and Black Stem (Ascochyta hyalospora and A. chenopodii)
3.1.2. Quinoa Black Stem (Ascochyta caulina)
3.1.3. Cockerel Eye/Quinoa Cercorporoid Leaf Spot
3.1.4. Cercospora Leaf Spot Caused by Cercospora cf. chenopodii
3.1.5. Quinoa Anthracnose Caused by Colletotrichum nigrum and C. truncatum
3.2. Fungi Identified by Morphological Approaches
3.2.1. Brown Stalk Rot
3.2.2. Quinoa Diamond Black Stem/“Mancha Ojival del Allo”
3.2.3. Sclerotium in Quinoa
3.2.4. Damping-Off
- Sensitivity of Pythium zingiberum and P. butleri oospores:Soil inoculation of oospores of P. zingiberum and P.butleri on soil caused damping-off of susceptible C. quinoa seedlings after ten days of incubation at 30 °C [160].
- Seedling damping-off caused by Fusarium avenacearum and Pytium aphanidermatum:The fungi were isolated from infected stems of quinoa seedlings grown in a greenhouse. Microbes were morphologically described and the cultured fungi were inoculated on C. quinoa cv. Cochabamba. Pathogenicity tests confirmed that P. aphanidermatum and F. avenaceum were the causal agents of the damping-off of quinoa seedlings under greenhouse conditions. The seedling infection was significantly higher up to the first pair of leaves, showing that quinoa is most susceptible to the pathogens before emergence. However, the sum of post-emergence damping-off was significantly lower than that observed in sugar beets and higher than that observed in cabbage plants, except for F. avenacearum, which also produced marked susceptibility at the first true leaves stage. In addition to the two pathogens, Ascochyta caulina, Fusarium spp., and Alternaria spp were also isolated from infected tissue but could not infect quinoa seedlings during pathogenicity tests [139].
- Pathogenicity tests on seedlings infected by Rhizoctonia solani and Fusarium spp.Rhizoctonia solani was isolated from the field in Peru. Pathogenicity tests performed in a greenhouse showed that R. solani prevented seed germination. It also created sunken lesions on the stems of old plants at ground level. Fusarium spp. reproduced wilting in old plants [4,161]. Quinoa seedling damping-off (Figure 18A) was observed during field experiments conducted at the experimental station of Nihon (Japan). It occurred from emergence until the four-leaf stage and increased under high soil moisture conditions. Rhizoctonia spp. (Figure 18B) and Fusarium spp. (Figure 18C) were identified morphologically from the symptomatic lesions [162].
- Pathogenicity tests on seedlings caused by Sclerotium rolfsii SaccSclerotium rolfsii was isolated from diseased seedlings of C. quinoa in a field of Southern California. The susceptibility of C. quinoa to S. rolfsii was demonstrated in vitro and under greenhouse conditions [163].

4. Chemical Control for Oomycetes and Fungi
5. Bacteria
5.1. Bacterial Leaf Spot Caused by Pseudomonas spp.
5.2. Bacterial Leaf Spot Caused by Pseudomonas syringae
6. Viruses
- Chenopodium mosaic virus: Seedlings of C. quinoa were found to contain a highly infectious, seed-borne virus that may remain latent. The virus was restricted to the Chenopodiaceae and was similar to the soybean mosaic virus in morphology and physio-chemical properties [169].
- Amaranthus leaf mottle virus (ALMV): Successful infections were achieved on C. quinoa, which exhibited chlorotic local lesions and severe systemic mosaic, leaf deformation, wilting, stunning, and finally collapse of the plants. Transmission via Aphis gossypii was confirmed 2 to 3 weeks after the 1-day inoculation access period [170].
- Ullucus virus C: UVC is a comovirus prevalent in Ullucus tuberosus grown at high altitudes in the Bolivian and Peruvian Andes. It was transmitted mechanically to C. amaranticolor and C. quinoa. It caused a systemic infection. UVC was not transmitted by either aphid species (Aphis gossypii or Myzus persicae) or through seeds of C. quinoa. However, it was transmitted through leaf contact between infected and healthy plants, causing chlorosis [173].
- Cucumber mosaic virus (CMV): Partially purified extracts from leaves of Phytolacca americana caused marked inhibition of CMV infection on C. quinoa [177].
- Tobacco mosaic virus: TMV has successfully infected C. quinoa [178].
- Passiflora latent virus (PLV): Chenopodium quinoa plants presenting systemic symptoms after inoculation with PLV showed high concentrations of virus particles in their cytoplasm, mitochondria, and chloroplasts [179]
- Plantago asiatica mosaic virus (PIAMV): Mechanical inoculation with infected sap of Lilium leaves on C. quinoa yielded chlorotic or necrotic local lesions [180].
- Carnation latent virus: C. quinoa is an indicator species for the carnation latent virus [181].
- Chlorotic leaf spot virus: Sap inoculation on C. quinoa resulted in a satisfactory infection [182].
7. Conclusions and Future Directions
- -
- Morphological identification paired with molecular tools for accurate descriptions of causal agents, published in scientific journals, as well as the sharing of knowledge within quinoa networks and conferences.
- -
- The performance of inclusive pathogenicity tests and Koch’s postulates to clarify the type of interaction observed (e.g., pathogenic, endophytic/symbiotic, or saprophytic.).
- -
- Standardized protocols for disease propagation and assessment methods for severity after infection.
- -
- The development of strategies for seed sanitation.
- -
- There exist several research centers located in areas where quinoa is traditionally grown, and recently a pilot global collaborative network on quinoa (GCN-Quinoa) (www.gcn-quinoa.org, accessed on 10 June 2021) has been established [189]. These networks primarily share knowledge on cultivation and plant breeding. Knowledge sharing in relation to quinoa diseases should also be considered.
- -
- More research on methodologies for the rapid, high throughput screening of quinoa seeds and plants for the presence of economically important pathogens of quinoa is needed. This would be useful for detecting causal agents early in disease development and ensuring certified pathogen-free quinoa seeds. Moreover, phone apps with deep learning models for diagnosing various plant diseases and pest attacks are becoming interesting tools, which may be useful in the future.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Latijnhouwers, M.; de Wit, P.J.G.M.; Govers, F. Oomycetes and Fungi: Similar Weaponry to Attack Plants. Trends Microbiol. 2003, 11, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Salas, B.; Otazu, V. Enfermedades. In Quinua y Kañiwa; IICA: Bogota, Colombia, 1976; pp. 137–148. [Google Scholar]
- Alandia, S.; Otazu, V.; Salas, B. Enfermedades. In Quinua y Kañiwa, Cultivos Andinos; Tapia, M.E., Ed.; Libros educativos; Instituto Interamericano de ciencias Agricolas IICA: Bogota, Colombia, 1979; Volume 49, pp. 137–148. [Google Scholar]
- Danielsen, S.; Bonifacio, A.; Ames, T. Diseases of Quinoa (Chenopodium Quinoa). Food Rev. Int. 2003, 19, 43–59. [Google Scholar] [CrossRef]
- Saravia, R.; Plata, G.; Gandarillas, A. Plagas y Enfermedades Del Cultivo de Quinua; Fundación PROINPA: Bolivia, Colombia, 2014. [Google Scholar]
- Gandarillas, A.; Saravia, R.; Quispe, R.; Ortiz-Romero, R. Principle quinoa pests and diseases. In State of the Art. Report on Quinoa around the World in 2013; FAO Regional Office for Latin America and the Caribean: Rome, Italy, 2015; pp. 192–217. [Google Scholar]
- Peintner, U.; Kuhnert-Finkernagel, R.; Wille, V.; Biasioli, F.; Shiryaev, A.; Perini, C. How to Resolve Cryptic Species of Polypores: An Example in Fomes. IMA Fungus 2019, 10, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruns, T.D.; White, T.J.; Taylor, J.W. Fungal Molecular Systematics. Annu. Rev. Ecol. Syst. 1991, 22, 525–564. [Google Scholar] [CrossRef]
- Klein, J.; Neilen, M.; van Verk, M.; Dutilh, B.E.; Van den Ackerveken, G. Genome Reconstruction of the Non-Culturable Spinach Downy Mildew Peronospora Effusa by Metagenome Filtering. PLoS ONE 2020, 15, e0225808. [Google Scholar] [CrossRef]
- Yahr, R.; Schoch, C.L.; Dentinger, B.T. Scaling up Discovery of Hidden Diversity in Fungi: Impacts of Barcoding Approaches. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150336. [Google Scholar] [CrossRef]
- Tekpinar, A.D.; Kalmer, A. Utility of Various Molecular Markers in Fungal Identification and Phylogeny. Nova Hedwig. 2019, 109, 187–224. [Google Scholar] [CrossRef]
- Seifert, K.A. Progress towards DNA Barcoding of Fungi. Mol. Ecol. Resour. 2009, 9, 83–89. [Google Scholar] [CrossRef]
- Lee, J.S.; Shin, H.-D.; Choi, Y.-J. Rediscovery of Seven Long-Forgotten Species of Peronospora and Plasmopara (Oomycota). Mycobiology 2020, 48, 331–340. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology, 5th ed.; Elservier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Grogan, R.G. The Science and Art of Plant-Disease Diagnosis. Annu. Rev. Phytopathol. 1981, 19, 333–351. [Google Scholar] [CrossRef]
- Grimes, D.J. Koch’s Postulates-Then and Now. Microbe Am. Soc. Microbiol. 2006, 1, 223. [Google Scholar] [CrossRef] [Green Version]
- Gäumann, E. Zur Kenntnis Der Chenopodiaceen Bewohnenden Peronospora-Arten. JSTOR 1920, 155. [Google Scholar]
- Baldauf, S.L. The Deep Roots of Eukaryotes. Science 2003, 300, 1703–1706. [Google Scholar] [CrossRef]
- Kamoun, S. Plant Pathogens: Oomycetes (water mold). In Encyclopedia of Microbiology, 3rd ed.; Schaechter, M., Ed.; Academic Press: Oxford, UK, 2009; pp. 689–695. ISBN 978-0-12-373944-5. [Google Scholar]
- Lee, S.C.; Ristaino, J.B.; Heitman, J. Parallels in Intercellular Communication in Oomycete and Fungal Pathogens of Plants and Humans. PLoS Pathog. 2012, 8, e1003028. [Google Scholar] [CrossRef] [Green Version]
- Griffith, J.M.; Davis, A.J.; Grant, B.R. Target Sites of Fungicides to Control Oomycetes. Target Sites Fungic. Action 1992, 69–100. [Google Scholar]
- Madoui, M.-A.; Bertrand-Michel, J.; Gaulin, E.; Dumas, B. Sterol Metabolism in the Oomycete Aphanomyces Euteiches, a Legume Root Pathogen. New Phytol. 2009, 183, 291–300. [Google Scholar] [CrossRef]
- Warrilow, A.G.; Hull, C.M.; Rolley, N.J.; Parker, J.E.; Nes, W.D.; Smith, S.N.; Kelly, D.E.; Kelly, S.L. Clotrimazole as a Potent Agent for Treating the Oomycete Fish Pathogen Saprolegnia Parasitica through Inhibition of Sterol 14α-Demethylase (CYP51). Appl. Environ. Microbiol. 2014, 80, 6154–6166. [Google Scholar] [CrossRef] [Green Version]
- Desmazières, J.B.H.J. Notice Sur Quelques Plantes Cryptogames Nouvellement Découvertes En France: Et Qui Vont Paraître, En Nature, Dans La Collection Publiée Par l’auteur; Annales des Sciences Naturelles Botanique: Paris, France, 1837. [Google Scholar]
- Yerkes, W.D.; Shaw, C.G. Taxonomy of the Peronospora Species on Cruciferaceae and Chenopodiaceae. Phytopathology 1959, 49. [Google Scholar]
- Choi, Y.-J.; Denchev, C.M.; Shin, H.-D. Morphological and Molecular Analyses Support the Existence of Host-Specific Peronospora Species Infecting Chenopodium. Mycopathologia 2008, 165, 155. [Google Scholar] [CrossRef]
- Byford, W.J. Field Experiments on Sugar-Beet Downy Mildew (Peronospora Farinosa). Ann. Appl. Biol. 1967, 60, 97–107. [Google Scholar] [CrossRef]
- Byford, W.J. Host Specialization of Peronospora Farinosa on Beta, Spinacia and Chenopodium. Trans. Br. Mycol. Soc. 1967, 50, 603–607. [Google Scholar] [CrossRef]
- Choi, Y.-J.; Danielsen, S.; Lübeck, M.; Hong, S.-B.; Delhey, R.; Shin, H.-D. Morphological and Molecular Characterization of the Causal Agent of Downy Mildew on Quinoa (Chenopodium Quinoa). Mycopathologia 2010, 169, 403–412. [Google Scholar] [CrossRef]
- Danielsen, S.; Lübeck, M. Universally Primed-PCR Indicates Geographical Variation of Peronospora Farinosa Ex. Chenopodium Quinoa. J. Basic Microbiol. 2010, 50, 104–109. [Google Scholar] [CrossRef]
- Kara, M.; Soylu, E.M.; Uysal, A.; Kurt, Ş.; Choi, Y.-J.; Soylu, S. Morphological and Molecular Characterization of Downy Mildew Disease Caused by Peronospora Variabilis on Chenopodium Album in Turkey. Australas. Plant Dis. Notes 2020, 15, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Testen, A.L.; del Mar Jiménez-Gasco, M.; Ochoa, J.B.; Backman, P.A. Molecular Detection of Peronospora Variabilis in Quinoa Seed and Phylogeny of the Quinoa Downy Mildew Pathogen in South America and the United States. Phytopathology 2014, 104, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Kandel, S.L.; Mou, B.; Shishkoff, N.; Shi, A.; Subbarao, K.V.; Klosterman, S.J. Spinach Downy Mildew: Advances in Our Understanding of the Disease Cycle and Prospects for Disease Management. Plant Dis. 2019, 103, 791–803. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.-J.; Hong, S.-B.; Shin, H.-D. Re-Consideration of Peronospora Farinosa Infecting Spinacia Oleracea as Distinct Species, Peronospora Effusa. Mycol. Res. 2007, 111, 381–391. [Google Scholar] [CrossRef]
- Choi, Y.-J.; Thines, M. (2288) Proposal to Reject the Name Botrytis Farinosa (Peronospora Farinosa)(Peronosporaceae: Oomycetes). Taxon 2014, 63, 675–676. [Google Scholar] [CrossRef]
- Stevenson, J.A.; Cardenas, M. Lista Preliminar de Los Hongos de Bolivia. Lilloa 1949, 21, 130. [Google Scholar]
- García Rada, G. Fitopatología Agrícola del Perú; Una Completa Recopilación de las Enfermedades Fungosas, Bacterianas y de Virus de las Plantas, Constatadas en el país, a Través de más de 20 años de Investigaciones; Estación Experimental Agrícola de la Molina: Lima, Peru, 1947. [Google Scholar]
- Tewari, J.P.; Boyetchko, S.M. Occurrence of Peronospora Farinosa f. Sp. Chenopodii on Quinoa in Canada. Can. Plant Dis. Surv. 1990, 70, 127–128. [Google Scholar]
- Aragón, L.; Gutiérrez, W. Downy Mildew on Four Chenopodium Species. Fitopatología 1992, 27, 104–109. [Google Scholar]
- Jacobs, T. Postulation of Virulence Groups and Resistance Factors in the Quinoa/Downy Mildew Pathosystem Using Material from Ecuador. Plant Pathol. 1999, 48, 425–430. [Google Scholar] [CrossRef]
- Danielsen, S.; Jacobsen, S.-E.; Hockenhull, J. First Report of Downy Mildew of Quinoa Caused by Peronospora Farinosa f. Sp. Chenopodii in Denmark. Plant Dis. 2002, 86, 1175. [Google Scholar] [CrossRef]
- Pańka, D.; Lenc, L.; Gęsiński, K. Preliminary Observations on Quinoa (Chenopodium Quinoa) Health Status in Poland. Phytopathol. Pol. 2004, 31, 61–66. [Google Scholar]
- Kumar, A.; Bhargava, A.; Shukla, S.; Singh, H.B.; Ohri, D. Screening of Exotic Chenopodium Quinoa Accessions for Downy Mildew Resistance under Mid-Eastern Conditions of India. Crop Prot. 2006, 25, 879–889. [Google Scholar] [CrossRef]
- Swenson, E.M. Genetic Diversity of Bolivian Peronospora Farinosa f.Sp. Chenopodii (Downy Mildew) and Quinoa’s Resistance Response. Master’s Thesis, Brigham Young University, Provo, UT, USA, 2006. [Google Scholar]
- Baiswar, P.; Chandra, S.; Kumar, R.; Ngachan, S.V. Peronospora Variabilis on Chenopodium Murale in India. Australas. Plant Dis. Notes 2010, 5, 45–47. [Google Scholar] [CrossRef] [Green Version]
- Testen, A.L.; McKemy, J.M.; Backman, P.A. First Report of Quinoa Downy Mildew Caused by Peronospora Variabilis in the United States. Plant Dis. 2012, 96, 146. [Google Scholar] [CrossRef]
- Choi, Y.J.; Choi, I.Y.; Kim, J.S.; Shin, H.-D. First Report of Quinoa Downy Mildew Caused by Peronospora Variabilis in Republic of Korea. Plant Dis. 2014, 98, 1003. [Google Scholar] [CrossRef]
- Mhada, M.; Ezzahiri, B.; Benlhabib, O. Assessment of Downy Mildew Resistance (Peronospora Farinosa) in a Quinoa (Chenopodium Quinoa Willd.) Germplasm. Int. J. Biol. Med. Res. 2015, 6, 4748–4752. [Google Scholar]
- Khalifa, W.; Thabet, M. Variation in Downy Mildew (Peronospora Variabilis Gäum) Resistance of Some Quinoa (Chenopodium Quinoa Willd) Cultivars under Egyptian Conditions. Middle East J. Agric. 2018, 70, 671–682. [Google Scholar]
- Nolen, H.B. Assessing Disease Concerns on Quinoa and Evaluating Sources of Disease Resistance in Chenopodium Species in New England. Master’s Thesis, University of new Hampshire, Durham, NH, USA, 2019. [Google Scholar]
- Gül, E. First Report of Peronospora Variabilis on Chenopodium Quinoa in Turkey. J. Plant Pathol. 2021, 103, 389–390. [Google Scholar] [CrossRef]
- Colque-Little, C.; Abondano, M.C.; Lund, O.S.; Amby, D.B.; Piepho, H.-P.; Andreasen, C.; Schmöckel, S.; Schmid, K. Genetic Variation for Tolerance to the Downy Mildew Pathogen Peronospora Variabilis in Genetic Resources of Quinoa (Chenopodium Quinoa). BMC Plant Biol. 2021, 21, 41. [Google Scholar] [CrossRef]
- El-Assiuty, E.M.; Taha, E.M.; Fahmy, Z.M.; Fahmy, G.M. Histological and Molecular Detections of Peronospora Variabilis Gäum Oospores in Seeds of Quinoa (Chenopodium Quino L.). Egypt. J. Exp. Biol. (Bot.) 2019, 15, 197–203. [Google Scholar]
- Danielsen, S.; Ames, T. Mildew (Peronospora Farinosa) of Quinua (Chenopodium Quinoa) in the Andean Region: Practical Manual for the Study of the Disease and Pathogen; International Potato Center: Lima, Peru, 2004. [Google Scholar]
- CABI Invasive Species Compendium 2020. Available online: https://www.cabi.org/publishing-products/invasive-species-compendium/ (accessed on 1 April 2021).
- Thines, M.; Choi, Y.-J. Evolution, Diversity, and Taxonomy of the Peronosporaceae, with Focus on the Genus Peronospora. Phytopathol. 2016, 106, 6–18. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.C.; Chauhan, L.S.; Mathur, R.S. Peronospora Farinosa (Fr.) Fr. on Chenopodium Murale L.-A New Record for India. Curr. Sci. 1964, 33, 720–721. [Google Scholar]
- Judelson, H.S. Sexual Reproduction in Oomycetes: Biology, Diversity and Contributions to Fitness. In Oomycete Genetics and Genomics: Diversity, Interactions and Research Tools; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2009; pp. 121–138. [Google Scholar]
- Kitz, L. Evaluation of Downy Mildew (Peronospora Farinosa f.Sp. Chenopodii) Resistance among Quinoa Genotypes and Investigation of P. Farinosa Growth Using Scanning Electron Microscopy; Brigham Young University: Provo, UT, USA, 2008. [Google Scholar]
- Cahill, D.M.; Hardham, A.R. Exploitation of Zoospore Taxis in the Development of a Novel Dipstick Immunoassay for the Specific Detection of Phytophthora Cinnamomi. Phytopathology 1994, 84, 193–200. [Google Scholar] [CrossRef]
- Kiefer, B.; Riemann, M.; Büche, C.; Kassemeyer, H.-H.; Nick, P. The Host Guides Morphogenesis and Stomatal Targeting in the Grapevine Pathogen Plasmopara Viticola. Planta 2002, 215, 387–393. [Google Scholar] [CrossRef]
- Michelmore, R.W.; Ilott, T.; Hulbert, S.H.; Farrara, B. The downy mildews. In Advances in Plant Pathology; Elsevier: Amsterdam, The Netherlands, 1988; Volume 6, pp. 53–79. [Google Scholar]
- Frinking, H.D.; Linders, E.G.A. A Comparison of Two Pathosystems: Downy Mildew on Spinacia Oleracea and on Chenopodium Album. Neth. J. Plant Pathol. 1986, 92, 97. [Google Scholar] [CrossRef]
- Doshi, A.; Thakore, B.B.L. Effect of Certain Environmental Factors on Germination of Conidia of Peronospora Arborescens (Berk) de Bary. J. Phytol. Res. 1993, 6, 59–62. [Google Scholar]
- Imamura, T.; Takagi, H.; Miyazato, A.; Ohki, S.; Mizukoshi, H.; Mori, M. Isolation and Characterization of the Betalain Biosynthesis Gene Involved in Hypocotyl Pigmentation of the Allotetraploid Chenopodium Quinoa. Biochem. Biophys. Res. Commun. 2018, 496, 280–286. [Google Scholar] [CrossRef]
- Polturak, G.; Aharoni, A. “La Vie En Rose”: Biosynthesis, Sources, and Applications of Betalain Pigments. Mol. Plant 2018, 11, 7–22. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, C.; Hajek, A.E.; Humber, R.A.; Bresciani, J.; Eilenberg, J. Soil as an Environment for Winter Survival of Aphid-Pathogenic Entomophthorales. Biol. Control 2003, 28, 92–100. [Google Scholar] [CrossRef]
- Taha, E.M. Molecular Detectionand Phylogeny of Peronospora Variabilis Gäum., The Causal Agent of Downy Mildew Disease of Quinoa at Different Growth Stages. Plant Cell Biotechnol. Mol. Biol. 2019, 20, 1189–1200. [Google Scholar]
- El-Assiuty, E.M.; Fahmy, G.M.; Taha, E.M.; Fahmy, Z.M.; Ismael, A.S.M.; Abd-Elghany, W.R.; Kafsheer, D.A. Microscopic Visualization of Peronospora Variabilis Gäum., the Cause of Quinoa Downy Mildew in Plant Tissues at Different Stages of Plant Growth. Int. J. Sci. Eng. Res. 2019, 10. [Google Scholar]
- Chapela, I.H. Spore Size Revisited: Analysis of Spore Populations Using Automated Particle Size. Sydowia 1991, 43, 1–14. [Google Scholar]
- Hamilton, A.J.; Cunnington, J.H. Calculating Minimum Sample Sizes for Taxonomic Measurements: Examples Using Gaumann’s Peronospora Spore Data. Mycotaxon 2006, 95, 189–194. [Google Scholar]
- Yin, H.; Zhou, J.B.; Chang, F.J.; Lu, H.; Gong, L.; Zhao, X. Identification of the pathogen causing downy mildew on Chenopodium quinoa. Acta Phytopathol. Sin. 2018, 48, 413–417. [Google Scholar] [CrossRef]
- Danielsen, S. Heterothallism in Peronospora Farinosa f. Sp. Chenopodii, the Causal Agent of Downy Mildew of Quinoa (Chenopodium Quinoa). J. Basic Microbiol. Int. J. Biochem. Physiol. Genet. Morphol. Ecol. Microorg. 2001, 41, 305–309. [Google Scholar]
- Fabritius, A.-L.; Cvitanich, C.; Judelson, H.S. Stage-Specific Gene Expression during Sexual Development in Phytophthora Infestans. Mol. Microbiol. 2002, 45, 1057–1066. [Google Scholar] [CrossRef] [Green Version]
- Danielsen, S.; Mercado, V.H.; Ames, T.; Munk, L. Seed Transmission of Downy Mildew (Peronospora Farinosa f.Sp. Chenopodii) in Quinoa and Effect of Relative Humidity on Seedling Infection. Seed Sci. Technol. 2004, 32, 91–98. [Google Scholar] [CrossRef]
- Vercesi, A.; Tornaghi, R.; Sant, S.; Burruano, S.; Faoro, F. A Cytological and Ultrastructural Study on the Maturation and Germination of Oospores of Plasmopara Viticola from Overwintering Vine Leaves. Mycol. Res. 1999, 103, 193–202. [Google Scholar] [CrossRef]
- Testen, A. Microbial Approaches to Support Andean Quinoa Production; The Pennsylvania State University: State College, PA, USA, 2012. [Google Scholar]
- Inaba, T.; Takahashi, K.; Morinaka, T. Seed Transmission of Spinach Downy Mildew. Plant Dis. 1983, 67, 1139–1141. [Google Scholar] [CrossRef]
- Inaba, T.; Hino, T. Production of Oospore-Encrusted Seeds by Inoculation of Conidia of Downy Mildew Fungus, Peronospora Manshurica, to Soybean Flower and Pod. Jpn. J. Phytopathol. 1980, 46, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Risco Mendoza, A. Severidad de Peronospora variabilis Gäum en Chenopodium quinoa Willd. “Pasankalla” como Respuesta a Plicaciones de Fungicidas y Bioestimulantes. Master’s Thesis, Universidad Nacional Agraria La Molina, Lima, Peru, 2014. [Google Scholar]
- Risco, A.; Mattos, L. Severidad de Peronospora Variabilis GÄUM. En Chenopodium Quinoa WILLD.‘Pasankalla’Como Respuesta a Aplicaciones de Fungicidas Sintéticos y Bioestimulantes. An. Científicos 2015, 76, 383–392. [Google Scholar] [CrossRef] [Green Version]
- Calixtro, G. Respuesta de 100 Accesiones de Guinua a la Infección Natural de Mildiu (Peronospora variabilis Gäum) en el Valle del Mantaro. Bachelor’s Thesis, Universidad Nacional Agraria La Molina, Lima, Peru, 2017. [Google Scholar]
- Risi, J.C.; Galwey, N.W. The Pattern of Genetic Diversity in the Andean Grain Crop Quinoa (Chenopodium Quinoa Willd). I. Associations between Characteristics. Euphytica 1989, 41, 147–162. [Google Scholar] [CrossRef]
- Mandák, B.; Krak, K.; Vít, P.; Lomonosova, M.N.; Belyayev, A.; Habibi, F.; Wang, L.; Douda, J.; Štorchová, H. Hybridization and Polyploidization within the Chenopodium Album Aggregate Analysed by Means of Cytological and Molecular Markers. Mol. Phylogenetics Evol. 2018, 129, 189–201. [Google Scholar] [CrossRef]
- Voglmayr, H.; Greilhuber, J. Genome Size Determination in Peronosporales (Oomycota) by Feulgen Image Analysis. Fungal Genet. Biol. 1998, 25, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Saito, K.; Liu, B.; Manley, A.; Kammeijer, K.; Mauzey, S.J.; Koike, S.; Correll, J.C. New Races and Novel Strains of the Spinach Downy Mildew Pathogen Peronospora Effusa. Plant Dis. 2018, 102, 613–618. [Google Scholar] [CrossRef] [Green Version]
- Irish, B.M.; Correll, J.C.; Koike, S.T.; Morelock, T.E. Three New Races of the Spinach Downy Mildew Pathogen Identified by a Modified Set of Spinach Differentials. Plant Dis. 2007, 91, 1392–1396. [Google Scholar] [CrossRef] [Green Version]
- Correll, J.; Koike, S.; Smilde, D.; de Visser, J. Race 15: New Spinach Downy Mildew Race Confirmed; Agriculture and Natural Resources, University of California: Los Angeles, CA, USA, 2014. [Google Scholar]
- Farber, D.H.; Mundt, C.C. Effect of Plant Age and Leaf Position on Susceptibility to Wheat Stripe Rust. Phytopathology 2017, 107, 412–417. [Google Scholar] [CrossRef]
- Panter, S.N.; Jones, D.A. Age-Related Resistance to Plant Pathogens. Adv. Bot. Res. 2002, 38, 251–280. [Google Scholar]
- Forbes, G.; Perez, W.; Andrade Piedra, J. Field Assessment of Resistance in Potato to Phytophthora Infestans: International Cooperators Guide; International Potato Center: Lima, Peru, 2014. [Google Scholar]
- Sosa-Zuniga, V.; Brito, V.; Fuentes, F.; Steinfort, U. Phenological Growth Stages of Quinoa (Chenopodium Quinoa) Based on the BBCH Scale. Ann. Appl. Biol. 2017, 171, 117–124. [Google Scholar] [CrossRef]
- Gabriel, J.; Luna, N.; Vargas, A.; Magne, J.; Angulo, A.; La Torre, J.; Bonifacio, A. Quinua de Valle (Chenopodium Quinoa Willd.): Fuente Valiosa de Resistencia Genética al Mildiu (Peronospora Farinosa Willd.). J. Selva Andin. Res. Soc. 2012, 3, 27–44. [Google Scholar] [CrossRef]
- Jacobsen, S.-E. Adaptation and scope for quinoa in northern latitudes of Europe. In State of the Art Report on Quinoa Around the World in 2013; FAO Regional Office for Latin America and the Caribean: Rome, Italy, 2015; pp. 431–446. [Google Scholar]
- Benlhabib, O.; Boujartani, N.; Maughan, P.J.; Jacobsen, S.E.; Jellen, E.N. Elevated Genetic Diversity in an F2: 6 Population of Quinoa (Chenopodium Quinoa) Developed through an Inter-Ecotype Cross. Front. Plant Sci. 2016, 7, 1222. [Google Scholar] [CrossRef] [Green Version]
- Bonifacio, A. Chenopodium Sp.: Genetic Resources, Ethnobotany, and Geographic Distribution. Food Rev. Int. 2003, 19, 1–7. [Google Scholar] [CrossRef]
- Danielsen, S.; Munk, L. Evaluation of Disease Assessment Methods in Quinoa for Their Ability to Predict Yield Loss Caused by Downy Mildew. Crop Prot. 2004, 23, 219–228. [Google Scholar] [CrossRef]
- Staub, J.; Bacher, J.; Poetter, K. Sources of Potential Errors in the Application of Random Amplified Polymorphic DNAs in Cucumber. HortScience 1996, 31, 262–266. [Google Scholar] [CrossRef] [Green Version]
- Hammerschmidt, R.; Métraux, J.-P.; Van Loon, L.C. Inducing Resistance: A Summary of Papers Presented at the First International Symposium on Induced Resistance to Plant Diseases, Corfu, May 2000. Eur. J. Plant Pathol. 2000, 107, 1–6. [Google Scholar] [CrossRef]
- Hammerschmidt, R. Induced Resistance to Disease in Plants; Springer: Berlin/Heidelberg, Germany, 1995. [Google Scholar]
- Strobel, N.E.; Ji, C.; Gopalan, S.; Kuc, J.A.; He, S.Y. Induction of Systemic Acquired Resistance in Cucumber by Pseudomonas Syringae Pv. Syringae 61 HrpZPss Protein. Plant J. 1996, 9, 431–439. [Google Scholar] [CrossRef]
- Colque-Little, C.X. Characterization of New Emerging Diseases Infecting Quinoa (Chenopodium Quinoa Willd.) and Search for Sources of Resistance to Downy Mildew (Peronospora Variavilis Gäum). Ph.D. Thesis, University of Copenhagen, Copenhagen, Denmark, 2021. [Google Scholar]
- La Torre-Farfan, J.P. Is Quinoa Cultivation on the Coastal Desert of Peru Sustainable? A Case Study from Majes, Arequipa. Master’s Thesis, Aarhus University, Aarhus, Denmark, 2017. [Google Scholar]
- Colque-Hinojosa, A.O. Collective Action and Contract Farming for Enhancing Smallholder Access to Markets: Case Studies of Farmer Organizations Participating in Rural Alliances Project in Bolivia. Master’s Thesis, Institute of Food and Resource Economics, University of Copenhagen, Copenhagen, Denmark, 2011. [Google Scholar]
- Gisi, U.; Sierotzki, H. Oomycete fungicides: Phenylamides, quinone outside inhibitors, and carboxylic acid amides. In Fungicide Resistance in Plant Pathogens; Springer: Berlin/Heidelberg, Germany, 2015; pp. 145–174. [Google Scholar]
- Du Toit, L.J.; Derie, M.L.; Brissey, L.M.; Holmes, B.J.; Gatch, E. Efficacy of Organic and Conventional Seed Treatments for Management of Verticillium in Spinach Seed. Phytopathology 2009, 99, S30. [Google Scholar]
- Leon Ttacca, B.; Ortiz Calcina, N.; Condori Ticona, N.; Chura Yupanqui, E. Cepas de Trichoderma Con Capacidad Endofitica Sobre El Control Del Mildiu (Peronospora Variabilis Gäum.) y Mejora Del Rendimiento de Quinua. Rev. Investig. Altoandinas 2018, 20, 19–30. [Google Scholar] [CrossRef]
- Aguilar, R.; More-Yarleque, M.M.; Rafael-Rutte, R.; Maldonado, E. Inductores de Defensa En El Control Del Mildiu (Peronospora Variabilis Gaum.) En El Cultivo de Quinua: Detección, Epidemiología, Síntomas, Características y Control. Sci. Agropecu. 2020, 11, 555–563. [Google Scholar] [CrossRef]
- Kiraly, Z.; Ersek, T.; Barna, B.; Adam, A.; Gullner, G. Pathophysiological Aspects of Plant Disease Resistance. Acta Phytopathol. Entomol. Hung. 1991, 26, 233–250. [Google Scholar]
- Van der Plank, J.E. Disease Resistance in Plants; Elsevier: Amsternad, The Netherlands, 1984. [Google Scholar]
- Gamboa, C.; Van den Broeck, G.; Maertens, M. Adoption of Improved Quinoa Varieties among Smallholder Farmers in the Peruvian Andes; CIP: Lima, Peru, 2018. [Google Scholar]
- Gandarillas, A.; Rojas, W.; Bonifacio, A.; Ojeda, N. Quinoa in Bolivia: The PROINPA foundation’s perspective. Chapter 5.1.a. In State of the Art Report on Quinoa around the World in 2013; Bazile, D., Bertero, D., Nieto, C., Eds.; FAO: La Paz, Bolivia, 2015; pp. 344–361. [Google Scholar]
- Gandarillas, H. Genetica y Origen. In Quinua y Kañiwa, Cultivos Andinos; Tapia, E.M., Ed.; Libros educativos; Instituto Interamericano de Ciencias Agricolas—IICA: Bogota, Colombia, 1979; Volume 49, pp. 45–64. [Google Scholar]
- McElhinny, E.; Mazon, N.; Rivera, M.; Peralta, I. Lineas promisorias de quinua con resistencia cuantitativa al mildiu en Ecuador. In Agro Biodiversidad y Produccion de Semilla con el Sector Informal a Traves del Mejoramiento participativo en la Zona Andina; PREDUZA, INIAP: Lima, Peru, 2003; pp. 40–47. [Google Scholar]
- Curti, R.N.; De la Vega, A.J.; Andrade, A.J.; Bramardi, S.J.; Bertero, H.D. Multi-Environmental Evaluation for Grain Yield and Its Physiological Determinants of Quinoa Genotypes across Northwest Argentina. Field Crops Res. 2014, 166, 46–57. [Google Scholar] [CrossRef]
- Poland, J.A.; Balint-Kurti, P.J.; Wisser, R.J.; Pratt, R.C.; Nelson, R.J. Shades of Gray: The World of Quantitative Disease Resistance. Trends Plant Sci. 2009, 14, 21–29. [Google Scholar] [CrossRef]
- Mastebroek, H.D.; Limburg, H.; Gilles, T.; Marvin, H.J.P. Occurrence of Sapogenins in Leaves and Seeds of Quinoa (Chenopodium Quinoa Willd). J. Sci. Food Agric. 2000, 80, 152–156. [Google Scholar] [CrossRef]
- Piepho, H.-P. A Mixed-Model Approach to Mapping Quantitative Trait Loci in Barley on the Basis of Multiple Environment Data. Genetics 2000, 156, 2043–2050. [Google Scholar] [CrossRef]
- Rojas, W.; Risi, J.; Bonifacio, A.; Gandarillas, A. El cultivo de la quinua en Bolivia. Capitulo 2. In Produccion y Mercado de la Quinua en Bolivia; Risi, J., Rojas, W., Pacheco, M., Eds.; IICA, Insitituo Interamericano de Investigacion Agricola: La Paz, Bolivia, 2015; pp. 33–67. [Google Scholar]
- Seiler, C.; Hutjes, R.W.; Kabat, P. Climate Variability and Trends in Bolivia. J. Appl. Meteorol. Climatol. 2013, 52, 130–146. [Google Scholar] [CrossRef] [Green Version]
- Cereceda, P.; Larrain, H.; Osses, P.; Farías, M.; Egaña, I. The Climate of the Coast and Fog Zone in the Tarapacá Region, Atacama Desert, Chile. Atmos. Res. 2008, 87, 301–311. [Google Scholar] [CrossRef]
- Lozano-Isla, F. Inti: Tools and Statistical Procedures in Plant Science. R Package Version 0.1.3; R Foundation: Vienna, Austria, 2021. [Google Scholar]
- Jellen, R.; Maughan, P.; Fuentes, F.; Kolano, B. Botany, phylogeny and evolution. Chapter 1.1. In State of the Art Report on Quinoa Around the World in 2013; Bazile, D., Bertero, D., Nieto, C., Eds.; FAO Regional Office for Latin America and the Caribean: Katowice, Poland, 2015; pp. 12–23. [Google Scholar]
- López-Marqués, R.L.; Nørrevang, A.F.; Ache, P.; Moog, M.; Visintainer, D.; Wendt, T.; Østerberg, J.T.; Dockter, C.; Jørgensen, M.E.; Salvador, A.T. Prospects for the Accelerated Improvement of the Resilient Crop Quinoa. J. Exp. Bot. 2020, 71, 5333–5347. [Google Scholar] [CrossRef]
- Kim, Y.C.; Blee, K.A.; Robins, J.; Anderson, A.J. OxycomTM under Field and Laboratory Conditions Increases Resistance Responses in Plants. Eur. J. Plant Pathol. 2001, 107, 129–136. [Google Scholar] [CrossRef]
- Bokshi, A.I.; Jobling, J.; McConchie, R. A Single Application of Milsana® Followed by Bion® Assists in the Control of Powdery Mildew in Cucumber and Helps Overcome Yield Losses. J. Hortic. Sci. Biotechnol. 2008, 83, 701–706. [Google Scholar] [CrossRef]
- Mathur, S.B.; Kongsdal, O. Common Laboratory Seed Health Testing Methods for Detecting Fungi; International Seed Testing Association: East Lansing, MI, USA, 2003. [Google Scholar]
- Boerema, G.H.; Mathur, S.B.; Neergaard, P. Ascochyta Hyalospora (Cooke & Ell.) Comb. Nov. in Seeds of Chenopodium Quinoa. Neth. J. Plant Pathol. 1977, 83, 153–159. [Google Scholar]
- Testen, A.L.; McKemy, J.M.; Backman, P.A. First Report of Ascochyta Leaf Spot of Quinoa Caused by Ascochyta Sp. in the United States. Plant Dis. 2013, 97, 844. [Google Scholar] [CrossRef]
- Otazu, V.; Aguilar, P.C.; Canahua, A. Resistance of Chenopodium quinoa to mildew (Peronospora effusa). Fitopatologia 1976, 11, 47–49. [Google Scholar]
- Fries, A.M.; Tapia, M.E. Guía de Campo de Los Cultivos Andinos; FAO, ANPE-PERÚ: Rome, Italy, 2007. [Google Scholar]
- Xiu-shi, Y.; Pei-you, Q.; Hui-min, G.; Gui-xing, R. Quinoa Industry Development in China. Int. J. Agric. Nat. Resour. 2019, 46, 208–219. [Google Scholar] [CrossRef]
- Boerema, G.H.; Loerakker, W.M.; Hamers, M.E. Check-List for Scientific Names of Common Parasitic Fungi. Supplement Series 2a (Additions and Corrections): Fungi on Field Crops: Beet and Potato; Caraway, Flax and Oil-Seed Poppy. Neth. J. Plant Pathol. 1987, 93, 1–20. [Google Scholar] [CrossRef]
- De Gruyter, J.; Woudenberg, J.H.C.; Aveskamp, M.M.; Verkley, G.J.M.; Groenewald, J.Z.; Crous, P.W. Redisposition of Phoma-like Anamorphs in Pleosporales. Stud. Mycol. 2013, 75, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Shivas, R.G.; Van der Quaedvlieg, W.; Van der Bank, M.; Zhang, Y.; Summerell, B.A.; Guarro, J.; Wingfield, M.J.; Wood, A.R.; Alfenas, A.C. Fungal Planet Description Sheets: 214–280. Pers. Mol. Phylogeny Evol. Fungi 2014, 32, 184. [Google Scholar] [CrossRef]
- Papizadeh, M.; Wijayawardene, N.N.; Amoozegar, M.A.; Saba, F.; Fazeli, S.A.S.; Hyde, K.D. Neocamarosporium Jorjanensis, N. Persepolisi, and N. Solicola Spp. Nov.(Neocamarosporiaceae, Pleosporales) Isolated from Saline Lakes of Iran Indicate the Possible Halotolerant Nature for the Genus. Mycol. Prog. 2018, 17, 661–679. [Google Scholar] [CrossRef]
- Yin, H.; Zhou, J.; Lv, H.; Qin, N.; Chang, F.J.; Zhao, X. Identification, Pathogenicity, and Fungicide Sensitivity of Ascochyta Caulina (Teleomorph: Neocamarosporium Calvescens) Associated with Black Stem on Quinoa in China. Plant Dis. 2020, 104, 2585–2597. [Google Scholar] [CrossRef]
- Van der Aa, H.A.; Van Kesteren, H.A. Some Pycnidial Fungi Occurring on Atriplex and Chenopodium. Pers. Mol. Phylogeny Evol. Fungi 1979, 10, 267–276. [Google Scholar]
- Dřímalková, M.; Veverka, K. Seedlings Damping-off of Chenopodium Quinoa Wild. Plant Protect. Sci 2004, 40, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhou, X.; Huang, H.; Li, G. Diseases Characteristic and Control Measurements for Chenopodium Quinoa Willd. In Proceedings of the 2017 6th International Conference on Energy and Environmental Protection (ICEEP 2017); Zhubai, China, 29–30 June 2017, Atlantis Press: Paris, France, 2017. [Google Scholar]
- Evidente, A.; Capasso, R.; Cutignano, A.; Taglialatela-Scafati, O.; Vurro, M.; Zonno, M.C.; Motta, A. Ascaulitoxin, a Phytotoxic Bis-Amino Acid N-Glucoside from Ascochyta Caulina. Phytochemistry 1998, 48, 1131–1137. [Google Scholar] [CrossRef]
- Muller-Scharer, H.; Scheepens, P.C.; Greaves, M.P. Biological Control of Weeds in European Crops: Recent Achievements and Future Work. Weed Res. 2000, 40, 83–98. [Google Scholar] [CrossRef]
- Scheepens, P.C.; Kempenaar, C.; Andreasen, C.; Eggers, T.H.; Netland, J.; Vurro, M. Biological Control of the Annual Weed Chenopodium Album, with Emphasis on the Application of Ascochyta Caulina as a Microbial Herbicide. Integr. Pest Manag. Rev. 1997, 2, 71–76. [Google Scholar] [CrossRef]
- Kempenaar, C.; Wanningen, R.; Scheepens, P.C. Control of Chenopodium Album by Soil Application of Ascochyta Caulina under Greenhouse Conditions. Ann. Appl. Biol. 1996, 129, 343–354. [Google Scholar] [CrossRef]
- Kempenaar, C.; Horsten, P.J.; Scheepens, P.C. Growth and Competitiveness of Common Lambsquarters (Chenopodium Album) after Foliar Application of Ascochyta Caulina as a Mycoherbicide. Weed Sci. 1996, 609–614. [Google Scholar] [CrossRef]
- Kempenaar, C.; Horsten, P.; Scheepens, P.C. Effects of Ascochyta Caulina on Photosynthesis of Leaves of Chenopodium Album. New Phytol. 1996, 132, 453–457. [Google Scholar] [CrossRef]
- Peralta, I. La Quinua En Ecuador.” Estado Del Arte”; INIAP: Quito, Ecuador, 2009. [Google Scholar]
- Świderska-Burek, U. Preliminary List of Cercosporoid Fungi from Poland. Mycotaxon 2007, 102, 5–8. [Google Scholar]
- Testen, A.L.; McKemy, J.M.; Backman, P.A. First Report of Passalora Leaf Spot of Quinoa Caused by Passalora Dubia in the United States. Plant Dis. 2013, 97, 139. [Google Scholar] [CrossRef]
- Sigovini, M.; Keppel, E.; Tagliapietra, D. Open Nomenclature in the Biodiversity Era. Methods Ecol. Evol. 2016, 7, 1217–1225. [Google Scholar] [CrossRef]
- Yin, H.; Zhou, J.; Lü, H.; Chang, F.; Qin, N.; Zhai, S.; Xing, K.; Zhao, F.; Zhao, X. Identification of the pathogen causing Cercospora leaf spot on quinoa. Acta Phytopathol. Sin. 2019, 49, 408–414. [Google Scholar]
- Videira, S.I.R.; Groenewald, J.Z.; Nakashima, C.; Braun, U.; Barreto, R.W.; de Wit, P.J.; Crous, P.W. Mycosphaerellaceae–Chaos or Clarity? Stud. Mycol. 2017, 87, 257–421. [Google Scholar] [CrossRef]
- Videira, S.I.R.; Groenewald, J.Z.; Braun, U.; Shin, H.-D.; Crous, P.W. All That Glitters Is Not Ramularia. Stud. Mycol. 2016, 83, 49–163. [Google Scholar] [CrossRef] [Green Version]
- Pal, N.; Testen, A.L. First Report of Quinoa Anthracnose Caused by Colletotrichum Nigrum and C. Truncatum in the United States. Plant Dis. 2020, 105, 3. [Google Scholar] [CrossRef]
- Boerema, G.H.; Höweler, L.H. Phoma Exigua Desm. and Its Varieties. Pers. Mol. Phylogeny Evol. Fungi 1967, 5, 15–28. [Google Scholar]
- Logan, C.; Khan, A.A. Comparative Studies of Phoma Spp. Associated with Potato Gangrene in Northern Ireland. Trans. Br. Mycol. Soc. 1969, 52, 9–17. [Google Scholar] [CrossRef]
- Otazu, V.; Salas, B. Brown Stalk Rot of Chenopodium Quinoa Caused by Phoma Exigua Var. Foveata. Fitoatologia 1977, 12, 54–58. [Google Scholar]
- Otazú, V.; Boerema, G.H.; Mooi, J.C.; Salas, B. Possible Geographical Origin of Phoma Exigua Var. Foveata, the Principal Causal Organism of Potato Gangrene. Potato Res. 1979, 22, 333–338. [Google Scholar] [CrossRef]
- Daza Vera, E. Enfermedad Micotica En Quinua. SITUA 1995, 29–32. [Google Scholar]
- Ikeda, A.; Ichitani, T. Density of Pythium Zingiberum Oospores in Relation to Infection of Chenopodium Quinoa Seedlings. Bull. Univ. Osaka Prefect. 1985, 37, 13–18. [Google Scholar]
- Barbosa, J.; Ames, T.; Jacobsen, S. Reduccion de La Germinacion y Muerte de Plantas de Chenopodium Quinoa Causadas Por Rhizoctonia y Fusarium; Centro Internacional de la Papa, Universidad Agraria la Molina: Lima, Peru, 2000. [Google Scholar]
- Isobe, K.; Sugiyama, T.; Katagiri, M.; Ishizuka, C.; Tamura, Y.; Higo, M.; Fujita, Y. Study on the Cause Damping-off in Quinoa (Chenopodium Quinoa Willd.) and a Method for Suppressing Its Occurrence. Jpn. J. Crop Sci. 2019, 88, 117–124. [Google Scholar] [CrossRef]
- Beckman, P.M.; Finch, H.C. Seed Rot and Damping-off of Chenopodium Quinoa Caused by Sclerotium Rolfsii. Plant Dis. 1980, 64, 497–498. [Google Scholar] [CrossRef]
- Masiello, M.; Somma, S.; Ghionna, V.; Logrieco, A.F.; Moretti, A. In Vitro and in Field Response of Different Fungicides against Aspergillus Flavus and Fusarium Species Causing Ear Rot Disease of Maize. Toxins 2019, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- Lebeda, A.; Cohen, Y. Fungicide resistance in Pseudoperonospora cubensis, the causal pathogen of cucurbit downy mildew. In Fungicide Resistance in Crop Protection: Risk and Management; CABI: Wallingford, UK, 2012; pp. 44–63. [Google Scholar]
- Plimmer, J.R. Encyclopedia of Agrochemicals; Wiley-Interscience: Hoboken, NJ, USA, 2003. [Google Scholar]
- Fonseca-Guerra, I.; Chiquillo, C.; Padilla, M.J.; Benavides-Rozo, M. First Report of Bacterial Leaf Spot on Chenopodium Quinoa Caused by Pseudomonas Syringae in Colombia. J. Plant Dis. Prot. 2021, 128, 871–874. [Google Scholar] [CrossRef]
- Walkey, D.G. Applied Plant Virology; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar]
- Dias, H.F.; Waterworth, H.E. The Identity of a Seed-Borne Mosaic Virus of Chenopodium Amaranticolor and C. Quinoa. Can. J. Bot. 1967, 45, 1285–1295. [Google Scholar] [CrossRef]
- Segundo, E.; Lesemann, D.E.; Martín, G.; Carmona, M.P.; Ruiz, L.; Cuadrado, I.M.; Velasco, L.; Janssen, D. Amaranthus Leaf Mottle Virus: 3′-End RNA Sequence Proves Classification as Distinct Virus and Reveals Affinities within the Genus Potyvirus. Eur. J. Plant Pathol. 2007, 117, 81–87. [Google Scholar] [CrossRef]
- Kenten, R.H.; Jones, R.A.C. Arracacha Virus B, a Second Isometric Virus Infecting Arracacha (Arracacia Xanthorrhiza; Umbelliferae) in the Peruvian Andes. Ann. Appl. Biol. 1979, 93, 31–36. [Google Scholar] [CrossRef]
- Jones, R.A.C.; Kenten, R.H. A Strain of Arracacha Virus B Infecting Oca (Oxalis Tuberosa: Oxalidaceae) in the Peruvian Andes. Phytopathol. Z. 1981, 100, 88–95. [Google Scholar] [CrossRef]
- Brunt, A.A.; Barton, R.J.; Phillips, S.; Jones, R.A.C. Ullucus Virus C, a Newly Recognised Comovirus Infecting Ullucus Tuberosus (Basellaceae). Ann. Appl. Biol. 1982, 101, 73–78. [Google Scholar] [CrossRef]
- Hiruki, C.; Shukla, P. Intracellular Location of Potato Virus S in Leaf Tissue of Chenopodium Quinoa. Can. J. Bot. 1973, 51, 1699–1702. [Google Scholar] [CrossRef]
- Herrera, V.V.; Juárez, E.R.; Muñiz, R.B. Detección de Virus Por Serología y Plantas Indicadoras En El Tubérculo-Semilla y Plantas de Cultivo de Meristemos En Papa (Solanum Tuberosum L.) Var. Alfa. Rev. Mex. De Fitopatol. 2003, 21, 176–180. [Google Scholar]
- Gibbs, A.J.; Hecht-Poinar, E.; Woods, R.D.; McKee, R.K. Some Properties of Three Related Viruses: Andean Potato Latent, Dulcamara Mottle, and Onions Yellow Mosaic. Microbiology 1966, 44, 177–193. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, J.A.; Walker, V.M.; Flewett, T.H.; Barclay, G.R. The Inhibition of Infection by Cucumber Mosaic Virus and Influenza Virus by Extracts from Phytolacca Americana. J. Gen. Virol. 1974, 22, 225–232. [Google Scholar] [CrossRef]
- Murray, R.R.; Emblow, M.S.; Hetherington, A.M.; Foster, G.D. Plant Virus Infections Control Stomatal Development. Sci. Rep. 2016, 6, 34507. [Google Scholar] [CrossRef] [Green Version]
- Bos, L.; Rubio-Huertos, M. Intracellular Accumulation of Passiflora Latent Virus in Chenopodium Quinoa. Neth. J. Plant Pathol. 1971, 77, 145–153. [Google Scholar] [CrossRef]
- Hammond, J.; Bampi, D.; Reinsel, M.D. First Report of Plantago Asiatica Mosaic Virus in Imported Asiatic and Oriental Lilies (Lilium Hybrids) in the United States. Plant Dis. 2014, 99, 292. [Google Scholar] [CrossRef]
- Hollings, M.; Stone, O.M. Chenopodium Quinoa Willd. as an Indicator Plant for Carnation Latent Virus. Plant Pathol. 1965, 14, 66–68. [Google Scholar] [CrossRef]
- Pfaeltzer, H.J. Chenopodium Quinoa, a Herbaceous Test Plant for Chlorotic Leaf Spot Virus in Apple. Neth. J. Plant Pathol. 1968, 74, 12–16. [Google Scholar] [CrossRef]
- Jelinek, C.F.; Pohland, A.E.; Wood, G.E. Worldwide Occurrence of Mycotoxins in Foods and Feeds—an Update. J. Assoc. Off. Anal. Chem. 1989, 72, 223–230. [Google Scholar] [CrossRef]
- Miller, J.D. Mycotoxins in Small Grains and Maize: Old Problems, New Challenges. Food Addit. Contam. 2008, 25, 219–230. [Google Scholar] [CrossRef]
- Magan, N.; Aldred, D. Post-Harvest Control Strategies: Minimizing Mycotoxins in the Food Chain. Int. J. Food Microbiol. 2007, 119, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Diaz, J.M.; Sulyok, M.; Jacobsen, S.-E.; Jouppila, K.; Nathanail, A.V. Comparative Study of Mycotoxin Occurrence in Andean and Cereal Grains Cultivated in South America and North Europe. Food Control 2021, 108260. [Google Scholar] [CrossRef]
- Kolakowski, B.; O’ROURKE, S.M.; Bietlot, H.P.; Kurz, K.; Aweryn, B. Ochratoxin A Concentrations in a Variety of Grain-Based and Non–Grain-Based Foods on the Canadian Retail Market from 2009 to 2014. J. Food Prot. 2016, 79, 2143–2159. [Google Scholar] [CrossRef]
- Woldemichael, G.M.; Wink, M. Identification and Biological Activities of Triterpenoid Saponins from Chenopodium Quinoa. J. Agric. Food Chem. 2001, 49, 2327–2332. [Google Scholar] [CrossRef]
- Murphy, K.M.; Bazile, D.; Kellogg, J.; Rahmanian, M. Development of a Worldwide Consortium on Evolutionary Participatory Breeding in Quinoa. Front. Plant Sci. 2016, 7, 608. [Google Scholar] [CrossRef]














| Country | C. quinoa Leaves (√), Seed (x) | C.album Leaves | C. berlandieri var. Macrocalycium | C. murale Leaves | C. ambrosoides Leaves | C. ficifolium Leaves | Researcher | Year | [Ref] | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mor. | Mol. | Mor. | Mol. | Mor. | Mol. | Mor. | Mol. | Mor. | Mol. | Mor. | Mol. | ||||
| Bolivia | √ | Martin Cardenas | 1941 | [36] | |||||||||||
| Peru | √ | G. Garcia | 1947 | [37] | |||||||||||
| Canada | √ | JF.Tewari | 1990 | [38] | |||||||||||
| Peru | √ | √ | √ | √ | L.Aragon | 1992 | [39] | ||||||||
| Ecuador | √ | Jose Ochoa | 1999 | [40] | |||||||||||
| Denmark | √ | S. Danielsen | 2002 | [41] | |||||||||||
| Poland | √ | Panka | 2004 | [42] | |||||||||||
| India | √ | A. Kumar | 2006 | [43] | |||||||||||
| Bolivia | √ | √ | Erica Swenson | 2006 | [44] | ||||||||||
| Argentina | √ | √ | Y.J. Choi | 2008 | [26] | ||||||||||
| China | √ | √ | 2010 | [29] | |||||||||||
| Ireland | √ | √ | |||||||||||||
| South Korea | √ | √ | |||||||||||||
| Netherlands | √ | √ | |||||||||||||
| Germany | √ | √ | |||||||||||||
| Latvia | √ | √ | |||||||||||||
| Romania | √ | √ | |||||||||||||
| Italy | √ | √ | |||||||||||||
| Peru | √ | S. Danielsen | 2010 | [30] | |||||||||||
| Ecuador | √ | ||||||||||||||
| Denmark | √ | ||||||||||||||
| India | √ | P. Baisvar | 2010 | [45] | |||||||||||
| USA (Pennsylvania) | √ | √ | Ana Testen | 2012 | [46] | ||||||||||
| Bolivia | x | Ana Testen | 2014 | [32] | |||||||||||
| Ecuador | x | ||||||||||||||
| USA | x | ||||||||||||||
| Korea | √ | √ | Y.J. Choi | 2014 | [47] | ||||||||||
| Morocco | - | - | Manal Mhada | 2014 | [48] | ||||||||||
| Egypt | √ | √ | Walaa Khalifa | 2018 | [49] | ||||||||||
| USA (N. Hampshire) | √ | √ | √ | √ | x√* | x√* | √** | Helen Nolen | 2019 | [50] | |||||
| Turkey | √ | √ | M.Kara | 2020 | [31] | ||||||||||
| Turkey | √ | √ | Esra Gül | 2021 | [51] | ||||||||||
| Denmark | √ | √ | √ | √ | C. Colque-Little | 2021 | [52] | ||||||||
| P. variabilis Sporangium Isolation Origin | ||
|---|---|---|
| C. quinoa | C. album | |
| av. Length × Width (µm) | av. Length × Width (µm) | Reference |
| 25.5 × 17.5 | Khalifa and Thabet 2018 [49] | |
| 22 × 23.13 | Yin et al., 2018 [72] | |
| 27.5 × 20 | Gül, 2021b [51] | |
| 28.8 × 21.8 | Danielsen & Ames, 2004 [54] | |
| 30.7 × 23.8 | Choi et al., 2010 [29] | |
| 31 × 23 | Testen et al., 2012 [46] | |
| 28.5 × 23.5 | Choi et al., 2014 [47] | |
| 29.5 × 23 | Choi et al., 2008 [26] | |
| 30 × 25 | Kara et al., 2020 [31] | |
| 27.7 × 21.0 | 30.1 × 24 | av. size |
| 1.32 | 1.25 | av.ratio |
| Cultivar | Hypothesized Resistance Factors | Response to Downy Mildew | Origin | |
|---|---|---|---|---|
| % Severity | % Sporulation | |||
| C. album | R1, R2, R3, R4 | 5 | 0.04 | Denmark |
| Puno | R1, R2, R3, R4 | 11 | 0.2 | Denmark |
| Rosa Blanca | R1, R2, R3 | 32 | 17 | Bolivia |
| Blanca | R1, R2, R3 | 46 | 47 | Bolivia |
| Titicaca | R1, R2 | 52 | 40 | Denmark |
| Vikinga | R1 | 70 | 69 | Denmark |
| Name | % Severity | % Sporulation | Spo/Sev Ratio | % Incidence | Ratio Based Classification |
|---|---|---|---|---|---|
| C. album | 5 | 0.4 | 0.08 | 45 | Resistant |
| Puno | 11 | 0.2 | 0.02 | 42 | Resistant |
| Cv6 (Rosa Blanca) | 32 | 17 | 0.53 | 59 | Highly tolerant |
| Cv17 (Canchis) | 41 | 30 | 0.73 | 73 | Mildly resistant |
| Cv18 (Pandela Roja) | 45 | 29 | 0.64 | 74 | Mildly resistant |
| Cv16 (Kurmi) | 45 | 50 | 1.1 | 56 | Susceptible |
| Blanca | 46 | 47 | 1 | 79 | Mildly susceptible |
| Cv8 (Blanquita) | 50 | 69 | 1.4 | 67 | Very susceptible |
| Titicaca | 52 | 40 | 0.77 | 81 | Mildly resistant |
| Cv3 (Ayrampu) | 52 | 63 | 1.2 | 77 | Susceptible |
| Cv20 (Aynoka) | 58 | 63 | 1.1 | 83 | Susceptible |
| Cv21 (Mariqueña) | 71 | 84 | 1.2 | 82 | Susceptible |
| Temperature | ||||||
|---|---|---|---|---|---|---|
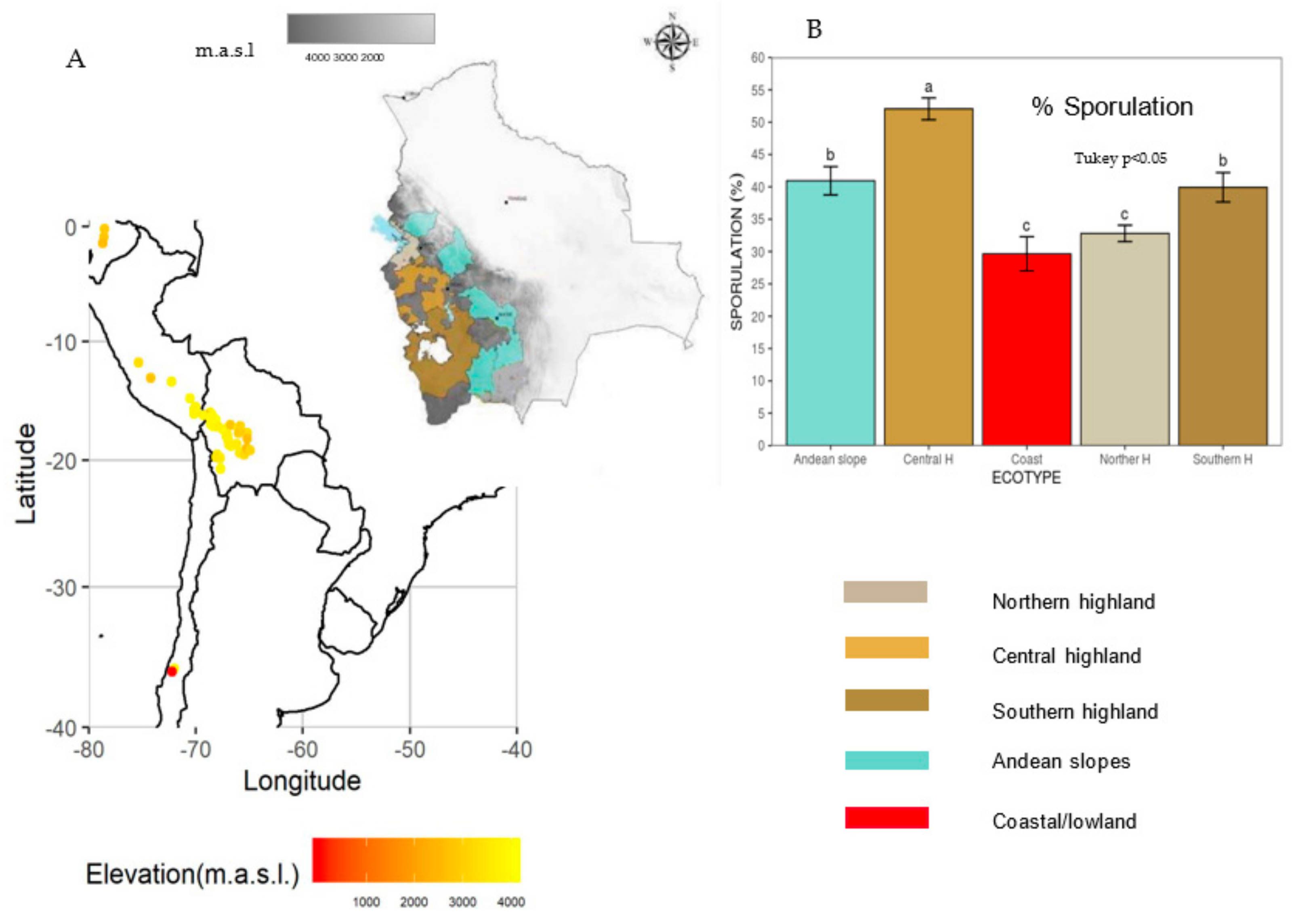
| Eco-Region | Soil | Altitude m.a.s.l | Rainfall (mm) | Max. | Min. | Av. |
| Northern Highland shores of Lake Titicaca | Rich in organic matter | 3500–4000 | 500 | 14 | 4 | 7 |
| Central Highland | Slightly acid | 3300–4100 | 350 | 17.7 | −2 | 8.7 |
| Southern Highland | Arid and poor soils | 3200–4000 | 50–200 | 18 | −11 | 5.7 |
| Andean Slopes | Variable | 800–3200 | 3500–700 | 12 | 3 | 7.6 |
| Coastal/Lowland Northern, Central, and Southern | Variable | Sea level to Mountain range | 40 > 2000 | 23 21 17 | −8 7 6 | 4.5 14 11 |
| Mode of Trans-location | Fungicide Group and Key Active Ingredients | Resistance Risk a | Foliar | Seed | Soil | Type of Activity | Translocation in Plants | Biochemical Mode of Action |
|---|---|---|---|---|---|---|---|---|
| Fully Systemic | Phenylamides: Metalaxyl, mefenoxam, oxadixyl, benalaxyl, kiralaxyl, ofurace | High | √ | √ | Preventive, curative, eradicative | Apoplastic, symplastic, translaminar | Inhibition of rRNA synthesis | |
| Partially Systemic | b Quinone outside inhibitors: Azoxystrobin, fenamidone, famox, adone, trifloxystrobin: kresoxin-methyl, Pyraclostrobin | √ | √ | Preventive | translaminar apoplastic | Inhibition of mitochondrial respiration at enzyme complex III | ||
| Non-Systemic | b Multisites: For example, mancozeb; chlorothalonil, copper, cu-oxychloride, cu-hydroxide; folpet; thiram, chlorothalonil | Low | √ | Preventive | Multi-site inhibition | |||
| Non-Systemic | Carboxylic acid amides: Dimethomorph, flumorph; iprovalicarb, benthiavalicarb; mandipropamid | Moderate | √ | Preventive | Translaminar | Cell wall synthesis, Ces3A cellulose synthase inhibition | ||
| Fully Systemic | Cyanoacetamide, oximes (cymoxanil) | Moderate | √ | √ | Preventive, curative | Apoplastic, symplastic, translaminar | Inhibition of mitochondrial respiration at the enzyme complex III | |
| Non-Systemic | Dinitroanilines (fluazinam) | Moderate | Preventive | Inhibition of ATP production | ||||
| Fully Systemic | Phosphonates (fosetyl-Al) | Moderate | √ | Preventive, curative | Apoplastic, symplastic, | Inhibition of spore germination, retardation of mycelia | ||
| Partially Systemic | Quinone inside respiration inhibitors: Cyazofamid, amisulbrom | Medium to hight | √ | √ | Preventive, curative, eradicative/ | Translaminar | ||
| Fully Systemic | Benzamides (fluopicolide) | Mod. | √ | √ | Preventive, curative | Apoplastic, symplastic, translaminar | Delocalization of spectrin-like proteins | |
| Benzamides, carboxamides Ethaboxam, zoxamide | Low | |||||||
| Systemic | Hymexaxol (heteroaromatics) | √ | √ | Fungal RNA and DNA syntheses | ||||
| Contact | b Thiadiazoles (Etridiazole) | √ | Preventive, curative | Lipid structure of Mitochondria | ||||
| Resistance inducer | Acibenzolar-S-methyl. | √ | ||||||
| b Demethylation inhibitor fungicides (DMIs): Imidazoles, triazolinthiones, triazoles prothioconazole, prochloraz, terbuconazole, difenoconazole | √ | √ | Preventive, curative | Sterole biosynthesis in membranes | ||||
| b PP fungicides (Phenylpyrroles) phenylpyrroles Fludioxonil | √ | √ | Preventive, curative | Signal transduction | ||||
| Fully Systemic | Carbamates: Propamocarb, prothiocarb | Preventive, eradicative | Apoplastic | Multi-site inhibition Affecting the membrane |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Colque-Little, C.; Amby, D.B.; Andreasen, C. A Review of Chenopodium quinoa (Willd.) Diseases—An Updated Perspective. Plants 2021, 10, 1228. https://doi.org/10.3390/plants10061228
Colque-Little C, Amby DB, Andreasen C. A Review of Chenopodium quinoa (Willd.) Diseases—An Updated Perspective. Plants. 2021; 10(6):1228. https://doi.org/10.3390/plants10061228
Chicago/Turabian StyleColque-Little, Carla, Daniel Buchvaldt Amby, and Christian Andreasen. 2021. "A Review of Chenopodium quinoa (Willd.) Diseases—An Updated Perspective" Plants 10, no. 6: 1228. https://doi.org/10.3390/plants10061228
APA StyleColque-Little, C., Amby, D. B., & Andreasen, C. (2021). A Review of Chenopodium quinoa (Willd.) Diseases—An Updated Perspective. Plants, 10(6), 1228. https://doi.org/10.3390/plants10061228