
New Insight into Taxonomy of European Mountain Pines, Pinus mugo Complex, Based on Complete Chloroplast Genomes Sequencing


Abstract
:1. Introduction
2. Results and Discussion
2.1. General Features of P. mugo, P. rotundata and P. uncinata Chloroplast Genomes
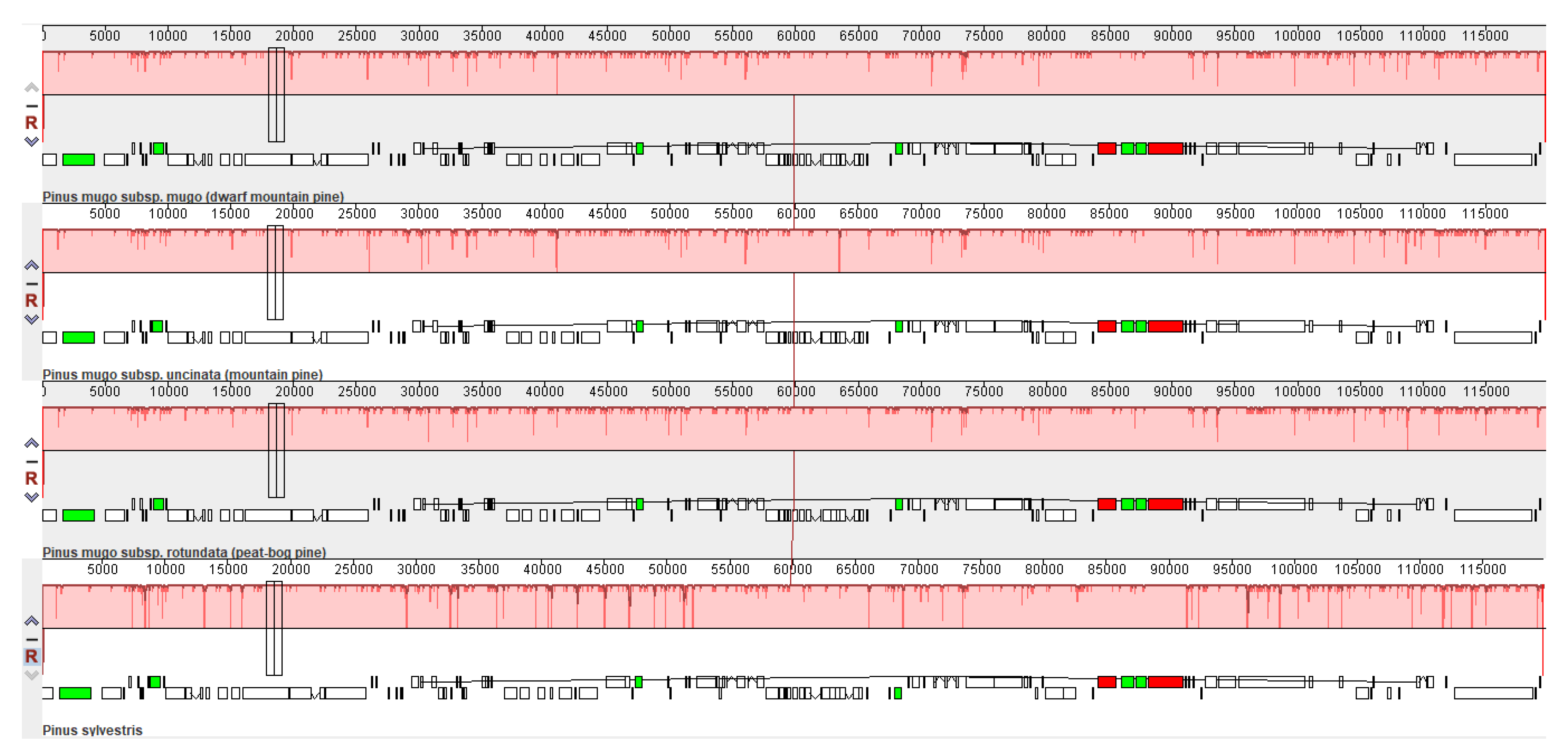
2.2. Genome Comparative Analysis and Identification of Divergent Hotspots
2.3. Simple Sequence Repeats Analysis
2.4. Phylogenetic Inference
3. Materials and Methods
3.1. Sampling, DNA Extraction and Genomic Library Preparation
3.2. Next Generation Sequencing
3.3. Chloroplast Genomes Assembly and Gene Annotation
3.4. Genome Comparative Analysis and Identification of Divergent Hotspots
3.5. Identification of Simple Sequence Repeats
3.6. Phylogenetic Inference
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Christensen, K.L. Taxonomic revision of the Pinus mugo complex and P. rhaetica (P. mugo sylvestris) (Pinaceae). Nord. J. Bot. 1987, 7, 383–408. [Google Scholar] [CrossRef]
- Hamernik, J.; Musil, I. The Pinus mugo complex−its structuring and general overview of the used nomenclature. J. For. Sci. 2008, 53, 253–266. [Google Scholar] [CrossRef] [Green Version]
- Critchfield, W.B.; Little, E.L. Geographic Distribution of the Pines of the World; Departament of Agriculture, Forest Service: Washington, DC, USA, 1966. [CrossRef] [Green Version]
- Ballian, D.; Ravazzi, C.; de Rigo, D.C. Pinus mugo in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., De Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016; pp. 124–125. [Google Scholar]
- Jalas, J.; Suominen, J. Atlas Florae Europaeae: Distribution of Vascular Plants in Europe. Gymnospermae, Volume 2; The Commitee for Mapping the Flora of Europe and Society Biology Fenn: Helsinki, Finland, 1973; p. 40. [Google Scholar]
- Businský, R.; Kirschner, J. Nomenclatural Notes on the Pinus mugo Complex in Central Europe. Phyt. Ann. Rei Bot. 2006, 46, 129–139. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2020-3. Available online: https://www.iucnredlist.org (accessed on 12 January 2021).
- Lewandowski, A.; Wiśniewska, M. Short Note: Crossability between Pinus Uliginosa and Its Putative Parental Species Pinus sylvestris and Pinus mugo. Silvae Genet. 2006, 55, 52–54. [Google Scholar] [CrossRef] [Green Version]
- Wachowiak, W.; Celiński, K.; Prus-Głowacki, W. Evidence of Natural Reciprocal Hybridisation between Pinus Uliginosa and P. sylvestris in the Sympatric Population of the Species. Flora Morphol. Distrib. Funct. Ecol. Plants 2005, 200, 563–568. [Google Scholar] [CrossRef]
- Wachowiak, W.; Prus-Głowacki, W. Hybridisation Processes in Sympatric Populations of Pines Pinus sylvestris L., P. mugo Turra and P. uliginosa Neumann. Plant Syst. Evol. 2008, 271, 29–40. [Google Scholar] [CrossRef]
- Wachowiak, W.; Żukowska, W.B.; Wójkiewicz, B.; Cavers, S.; Litkowiec, M. Hybridization in Contact Zone between Temperate European Pine Species. Tree Genet. Genomes 2016, 12. [Google Scholar] [CrossRef] [Green Version]
- Danielewicz, W.Z.J. Ochrona Sosny Błotnej Pinus Uliginosa A. Neumann Na Terenie Borów Dolnośląskich. Przegląd Przyr. 2000, 11, 113–124. [Google Scholar]
- Gołąb, Z. Sosna Błotna (Pinus Uliginosa Neumann) Na Wielkim Torfowisku Batorowskim w Górach Stołowych. Szczeliniec 1999, 3, 41–48. [Google Scholar]
- Boratyńska, K.; Boratyński, A. Taxonomic Differences among Closely Related Pines Pinus sylvestris, P. mugo, P. uncinata, P. rotundata and P. uliginosa as Revealed in Needle Sclerenchyma Cells. Flora Morphol. Distrib. Funct. Ecol. Plants 2007, 202, 555–569. [Google Scholar] [CrossRef]
- Boratyńska, K.M. Pinus Uncinata Ramond Taxonomy Basedon Needle Characters. Plant Syst. Evol. 2001, 227, 183–194. [Google Scholar]
- Boratyńska, K.; Boratyński, A.; Lewandowski, A. Morphology of Pinus Uliginosa (Pinaceae) Needles from Populations Exposed to and Isolated from the Direct Influence of Pinus sylvestris. Bot. J. Linn. Soc. 2003, 124, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Siedlewska, A.; Prus-Głowacki, W. Genetic Structure and Taxonomic Position of Pinus Uliginosa Neumann Population from Wielkie Torfowisko Batorowskie in Stołowe Mts. Locus Classicus. Acta Soc. Bot. Pol. 1995, 64, 51–58. [Google Scholar] [CrossRef]
- Lewandowski, A.; Burczyk, J.; Wachowiak, W.; Boratyński, A.; Prus-Głowacki, W. Genetic Evaluation of Seeds of Highly Endangered Pinus Uliginosa Neumann from Wȩgliniec Reserve for Ex-Situ Conservation Program. Acta Soc. Bot. Pol. 2005, 74, 237–242. [Google Scholar] [CrossRef] [Green Version]
- Celiński, K.; Pawlaczyk, E.M.; Wojnicka-Półtorak, A.; Chudzińska, E.; Prus-Głowacki, W. Cross-Species Amplification and Characterization of Microsatellite Loci in Pinus mugo Turra. Biology 2013, 68, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Heuertz, M.; Teufel, J.; González-Martínez, S.C.; Soto, A.; Fady, B.; Alía, R.; Vendramin, G.G. Geography Determines Genetic Relationships between Species of Mountain Pine (Pinus mugo Complex) in Western Europe. J. Biogeogr. 2010, 37, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Celiński, K.; Zbránková, V.; Wojnicka-Półtorak, A.; Chudzińska, E. Biogeography and evolutionary factors determine genetic differentiation of Pinus mugo (Turra) in the Tatra Mountains (Central Europe). J. Mt. Sci. 2015, 12, 549–557. [Google Scholar] [CrossRef]
- Danusevičius, D.; Marozas, V.; Brazaitis, G.; Petrokas, R.; Christensen, K.I. Spontaneous Hybridization between Pinus mugo and Pinus sylvestris at the Lithuanian Seaside: A Morphological Survey. Sci. World J. 2012, 2012, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kormutak, A.; Demankova, B.; Gömöry, D. Spontaneous Hybridization between Pinus sylvestris L. and P. mugo Turra in Slovakia. Silvae Genet. 2008, 57, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Kormutak, A.; Galgoci, M.; Bolecek, P.; Gőmőry, D.; Libantova, J. Reinforced evidence on partial compatibility between Pinus sylvestris and Pinus mugo and on maternal inheritance of chloroplast DNA in the Pinus mugo × Pinus sylvestris cross. Silvae Genet. 2020, 69, 108–115. [Google Scholar] [CrossRef]
- Bogunić, F.; Siljak-Yakovlev, S.; Muratovic, E.; Pustahija, F.; Medjedović, S. Molecular cytogenetics and flow cytometry reveal conserved genome organization in Pinus mugo and P. uncinata. Ann. For. Sci. 2011, 68, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Celiński, K.; Chudzińska, E.; Gmur, A.; Piosik, Ł.; Wojnicka-Półtorak, A. Cytological characterization of three closely related pines-Pinus mugo, P. uliginosa and P. × rhaetica from the Pinus mugo complex (Pinaceae). Biology 2019, 74, 751–756. [Google Scholar] [CrossRef] [Green Version]
- Lewandowski, A.; Boratyński, A.; Mejnartowicz, L. Allozyme Investigations on the Genetic Differentiation between Closely Related Pines—Pinus sylvestris, P. mugo, P. uncinata, and P. uliginosa (Pinaceae). Plant Syst. Evol. 2000, 221, 15–24. [Google Scholar] [CrossRef]
- Prus-Głowacki, W.; Bujas, E.; Ratyńska, H. Taxonomic Position of Pinus Uliginosa Neumann as Related to Other Taxa of Pinus mugo Complex. Acta Soc. Bot. Pol. 1998, 67, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Celiński, K.; Kijak, H.; Barylski, J.; Grabsztunowicz, M.; Wojnicka-Półtorak, A.; Chudzińska, E. Characterization of the complete chloroplast genome of Pinus uliginosa (Neumann) from the Pinus mugo complex. Conserv. Genet. Resour. 2016, 9, 209–212. [Google Scholar] [CrossRef] [Green Version]
- Celiński, K.; Kijak, H.; Wojnicka-Półtorak, A.; Buczkowska-Chmielewska, K.; Sokołowska, J.; Chudzińska, E. Effectiveness of the DNA barcoding approach for closely related conifers discrimination: A case study of the Pinus mugo complex. Comptes Rendus Biol. 2017, 340, 339–348. [Google Scholar] [CrossRef]
- Celiński, K.; Sokołowska, J.; Zemleduch-Barylska, A.; Kuna, R.; Kijak, H.; Staszak, A.M.; Wojnicka-Półtorak, A.; Chudzińska, E. Seed Total Protein Profiling in Discrimination of Closely Related Pines: Evidence from the Pinus mugo Complex. Plants 2020, 9, 872. [Google Scholar] [CrossRef]
- Celiński, K.; Bonikowski, R.; Wojnicka-Półtorak, A.; Chudzińska, E.; Maliński, T. Volatiles as Chemosystematic Markers for Distinguishing Closely Related Species within the Pinus mugo Complex. Chem. Biodivers. 2015, 12, 1208–1213. [Google Scholar] [CrossRef]
- Bonikowski, R.; Celinski, K.; Wojnicka-Półtorak, A.; Maliński, T. Composition of Essential Oils Isolated from the Needles of Pinus uncinata and P. uliginosa Grown in Poland. Nat. Prod. Commun. 2015, 10, 371–373. [Google Scholar] [CrossRef] [Green Version]
- Cavers, S.; Wachowiak, W.; Boratyńska, K. Geographical Patterns of Nucleotide Diversity and Population Differentiation in Three Closely Related European Pine Species in the Pinus mugo Complex. Bot. J. Linn. Soc. 2013, 172, 225–238. [Google Scholar]
- Li, X.; Yang, Y.; Henry, R.; Rossetto, M.; Wang, Y.; Chen, S. Plant DNA barcoding: From gene to genome. Biol. Rev. 2015, 90, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Sudianto, E.; Wu, C.-S.; Lin, C.-P.; Chaw, S.-M. Revisiting the Plastid Phylogenomics of Pinaceae with Two Complete Plastomes of Pseudolarix and Tsuga. Genome Biol. Evol. 2016, 8, 1804–1811. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Zhao, T.; Ma, Q.; Liang, L.; Wang, G. Comparative Genomics and Phylogenetic Analysis Revealed the Chloroplast Genome Variation and Interspecific Relationships of Corylus (Betulaceae) Species. Front. Plant Sci. 2018, 9, 927. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sylvester, S.P.; Li, M.; Zhang, C.; Li, X.; Duan, Y.; Wang, X. The Complete Plastid Genome of Magnolia zenii and Genetic Comparison to Magnoliaceae species. Molecules 2019, 24, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Li, Y.; Zang, M.; Li, M.; Fang, Y. Complete Chloroplast Genome Sequence and Phylogenetic Analysis of Quercus acutissima. Int. J. Mol. Sci. 2018, 19, 2443. [Google Scholar] [CrossRef] [Green Version]
- Jansen, R.K.; Ruhlman, T.A. Plastid Genomes of Seed Plants. In Genomics of Chloroplasts and Mitochondria. Advances in Photosynthesis and Respiration (Including Bioenergy and Related Processes); Bock, R., Knoop, V., Eds.; Springer: Dordrecht, The Netherlands, 2012; Volume 35, pp. 103–126. [Google Scholar]
- Lin, M.; Qi, X.; Chen, J.; Sun, L.; Zhong, Y.; Fang, J.; Hu, C. The complete chloroplast genome sequence of Actinidia arguta using the PacBio RS II platform. PLoS ONE 2018, 13, e0197393. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.-I.; Lee, H.O.; Lee, I.H.; Kim, I.S.; Lee, S.-W.; Yang, T.J.; Shim, D. Complete Chloroplast Genome of Pinus densiflora Siebold & Zucc. and Comparative Analysis with Five Pine Trees. Forests 2019, 10, 600. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Chen, L.; Yi, X.; Li, M. The complete chloroplast genome of Pinus yunnanensis Franchet (Pinaceae). Mitochondrial DNA Part B 2019, 4, 2600–2601. [Google Scholar] [CrossRef]
- Kim, K.-J. Complete Chloroplast Genome Sequences from Korean Ginseng (Panax schinseng Nees) and Comparative Analysis of Sequence Evolution among 17 Vascular Plants. DNA Res. 2004, 11, 247–261. [Google Scholar] [CrossRef]
- Perry, A.; Wolfe, K.H. Nucleotide Substitution Rates in Legume Chloroplast DNA Depend on the Presence of the Inverted Repeat. J. Mol. Evol. 2002, 55, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Chen, H.; He, S.; Wang, L.; Chen, A.J.; Liu, C. Sequencing, Characterization, and Comparative Analyses of the Plastome of Caragana rosea var. rosea. Int. J. Mol. Sci. 2018, 19, 1419. [Google Scholar] [CrossRef] [Green Version]
- Yi, X.; Gao, L.; Wang, B.; Su, Y.-J.; Wang, T. The Complete Chloroplast Genome Sequence of Cephalotaxus oliveri (Cephalotaxaceae): Evolutionary Comparison of Cephalotaxus Chloroplast DNAs and Insights into the Loss of Inverted Repeat Copies in Gymnosperms. Genome Biol. Evol. 2013, 5, 688–698. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Celiński, K.; Kijak, H.; Wiland-Szymańska, J. Complete Chloroplast Genome Sequence and Phylogenetic Inference of the Canary Islands Dragon Tree (Dracaena draco L.). Forests 2020, 11, 309. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Xu, C.; Li, C.; Sun, J.; Zuo, Y.; Shi, S.; Cheng, T.; Guo, J.; Zhou, S. ycf1, the most promising plastid DNA barcode of land plants. Sci. Rep. 2015, 5, 8348. [Google Scholar] [CrossRef] [Green Version]
- Olsson, S.; Grivet, D.; Cid-Vian, J. Species-diagnostic markers in the genus Pinus: Evaluation of the chloroplast regions matK and ycf. For. Syst. 2018, 27, e016. [Google Scholar] [CrossRef] [Green Version]
- Ellegren, H. Microsatellites: Simple sequences with complex evolution. Nat. Rev. Genet. 2004, 5, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak, L.; Wojnicka-Półtorak, A.; Celinski, K.; Lesiczka, P.; Pawlaczyk, E.; Aučina, A. Genetic resources of relict populations of Pinus sylvestris (L.) in Western Carpathians assessed by chloroplast microsatellites. Biologia 2019, 74, 1077–1086. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, E.; Pádua, J.G.; Zucchi, M.I.; Vencovsky, R.; Vieira, M.L.C. Origin, evolution and genome distribution of microsatellites. Genet. Mol. Biol. 2006, 29, 294–307. [Google Scholar] [CrossRef]
- Gómez, A.; González-Martínez, S.C.; Collada, C.; Climent, J.; Gil, L. Complex population genetic structure in the endemic Canary Island pine revealed using chloroplast microsatellite markers. Theor. Appl. Genet. 2003, 107, 1123–1131. [Google Scholar] [CrossRef] [PubMed]
- Asaf, S.; Khan, A.L.; Khan, M.A.; Shahzad, R.; Lubna; Kang, S.M.; Al-Harrasi, A.; Al-Rawahi, A.; Lee, I.-J. Complete chloroplast genome sequence and comparative analysis of loblolly pine (Pinus taeda L.) with related species. PLoS ONE 2018, 13, e0192966. [Google Scholar] [CrossRef] [Green Version]
- Dzialuk, A.; Boratyńska, K.; Romo, A.; Boratynski, A. Taxonomic and geographic variation of the Pinus mugo complex on chloroplast microsatellite markers. Syst. Biodivers. 2016, 15, 464–479. [Google Scholar] [CrossRef] [Green Version]
- Dzialuk, A.; Muchewicz, E.; Boratyński, A.; Montserrat, J.M.; Boratyńska, K.; Burczyk, J. Genetic variation of Pinus uncinata (Pinaceae) in the Pyrenees determined with cpSSR markers. Plant Syst. Evol. 2009, 277, 197–205. [Google Scholar] [CrossRef]
- Li, D.-M.; Zhao, C.-Y.; Liu, X.-F. Complete Chloroplast Genome Sequences of Kaempferia Galanga and Kaempferia Elegans: Molecular Structures and Comparative Analysis. Molecules 2019, 24, 474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, H.-T.; Tran, N.; Nguyen, T.-D.; Vu, Q.-L.; Bui, M.-H.; Le, M.-T.; Le, L. Complete Chloroplast Genome of Paphiopedilum delenatii and Phylogenetic Relationships among Orchidaceae. Plants 2020, 9, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Zhang, Y.; Song, M.; Guan, Y.; Ma, X. Species Identification of Dracaena Using the Complete Chloroplast Genome as a Super-Barcode. Front. Pharmacol. 2019, 10, 1441. [Google Scholar] [CrossRef]
- Ge, J.; Cai, L.; Bi, G.-Q.; Chen, G.; Sun, W. Characterization of the Complete Chloroplast Genomes of Buddleja colvilei and B. sessilifolia: Implications for the Taxonomy of Buddleja L. Molecules 2018, 23, 1248. [Google Scholar] [CrossRef] [Green Version]
- Hernández-León, S.; Gernandt, D.S.; De La Rosa, J.A.P.; Barbolla, L.J. Phylogenetic Relationships and Species Delimitation in Pinus Section Trifoliae Inferrred from Plastid DNA. PLoS ONE 2013, 8, e70501. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of Plants DNA from Fresh Tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Chen, H.; Jiang, M.; Wang, L.; Wu, X.; Huang, L.; Liu, C. CPGAVAS2, an integrated plastome sequence annotator and analyzer. Nucleic Acids Res. 2019, 47, W65–W73. [Google Scholar] [CrossRef] [PubMed]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq–versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA Genes in Genomic Sequences. Methods Mol. Biol. 2019, 1962, 1–14. [Google Scholar] [CrossRef]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [Green Version]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple Genome Alignment with Gene Gain, Loss and Rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Bi, Y.; Zhang, M.-F.; Xue, J.; Dong, R.; Du, Y.-P.; Zhang, X.-H. Chloroplast genomic resources for phylogeny and DNA barcoding: A case study on Fritillaria. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef] [Green Version]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genome Features | Pinus mugo | Pinus rotundata | Pinus uncinata | Pinus sylvestris | Pinus densiflora | Larix decidua | Abies alba |
|---|---|---|---|---|---|---|---|
| Genome size (bp) | 119,765 | 119,759 | 119,780 | 119,758 | 119,875 | 122,747 | 121,243 |
| Total coding length (bp) | 67,592 | 67,593 | 67,592 | 67,625 | 67,684 | 68,621 | 67,983 |
| Protein coding length (bp) | 60,339 | 60,339 | 60,339 | 60,384 | 60,444 | 61,524 | 60,810 |
| rRNA coding length (bp) | 4517 | 4518 | 4518 | 4518 | 4518 | 4520 | 4522 |
| tRNA coding length (bp) | 2736 | 2736 | 2735 | 2654 | 2723 | 2577 | 2651 |
| Total GC content (%) | 38.5 | 38.5 | 38.5 | 38.5 | 38.5 | 38.8 | 38.3 |
| Total number of genes | 121 | 121 | 121 | 116 | 118 | 110 | 113 |
| Number of protein-coding genes | 73 | 73 | 73 | 73 | 73 | 72 | 74 |
| Number of rRNA genes | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| Number of tRNA genes | 36 | 36 | 36 | 35 | 36 | 34 | 35 |
| GenBank Acc. No. | MZ333466 | MZ333465 | MZ333464 | KR476379 | MK285358 | AB501189 | NC_042410 |
| P. mugo | P. rotundata | P. uncinata | P. sylvestris | |
|---|---|---|---|---|
| P. mugo | - | 0.000044 | 0.000055 | 0.000158 |
| P. rotundata | 0.000259 | - | 0.000061 | 0.000158 |
| P. uncinata | 0.000351 | 0.000409 | - | 0.000158 |
| P. sylvestris | 0.003117 | 0.003126 | 0.003184 | - |
| Taxon | ID | Type | Repeat Motif | Length (bp) | Start | End | Location | ID | Type | Repeat Motif | Length (bp) | Start | End | Location |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P. mugo | 1 | p1 | (C)12 | 12 | 15142 | 15153 | IGS | 11 | p1 | (A)11 | 11 | 79749 | 79759 | IGS |
| 2 | p1 | (A)12 | 12 | 26050 | 26061 | IGS | 12 | p1 | (T)10 | 10 | 87077 | 87086 | IGS | |
| 3 | c | (A)10(G)10 | 20 | 30198 | 30217 | IGS | 13 | p1 | (A)10 | 10 | 100605 | 100614 | IGS | |
| 4 | p1 | (T)23 | 23 | 40994 | 41016 | IGS | 14 | p1 | (T)10 | 10 | 103575 | 103584 | IGS | |
| 5 | p1 | (T)13 | 13 | 44949 | 44961 | IGS | 15 | p1 | (G)11 | 11 | 104142 | 104152 | CDS (ndhD) | |
| 6 | p1 | (T)10 | 10 | 48132 | 48141 | IGS | 16 | p1 | (A)13 | 13 | 106928 | 106940 | IGS | |
| 7 | p1 | (A)10 | 10 | 54429 | 54438 | IGS | 17 | p1 | (T)11 | 11 | 107335 | 107345 | CDS (rpl32) | |
| 8 | p1 | (A)10 | 10 | 67826 | 67835 | IGS | 18 | p1 | (A)10 | 10 | 109379 | 109388 | IGS | |
| 9 | p1 | (T)11 | 11 | 71751 | 71761 | CDS (ycf3) | 19 | p1 | (A)10 | 10 | 109840 | 109849 | CDS (rps12) | |
| 10 | p2 | (AT)6 | 12 | 73254 | 73265 | IGS | 20 | p2 | (AT)6 | 12 | 111752 | 111763 | IGS | |
| 1 | p1 | (C)13 | 13 | 15141 | 15153 | IGS | 11 | p1 | (T)11 | 11 | 87069 | 87079 | IGS | |
| 2 | p1 | (A)12 | 12 | 26050 | 26061 | IGS | 12 | p1 | (A)10 | 10 | 100597 | 100606 | IGS | |
| P. rotundata | 3 | c | (A)11(G)10 | 21 | 30197 | 30217 | IGS | 13 | p1 | (T)10 | 10 | 100883 | 100892 | IGS |
| 4 | p1 | (T)15 | 15 | 40993 | 41007 | IGS | 14 | p1 | (T)10 | 10 | 103568 | 103577 | IGS | |
| 5 | p1 | (T)15 | 15 | 44940 | 44954 | IGS | 15 | p1 | (G)10 | 10 | 104135 | 104144 | CDS (ndhD) | |
| 6 | p1 | (T)11 | 11 | 48122 | 48132 | IGS | 16 | p1 | (A)13 | 13 | 106920 | 106932 | IGS | |
| 7 | p1 | (A)10 | 10 | 67816 | 67825 | IGS | 17 | p1 | (T)11 | 11 | 107327 | 107337 | CDS (rpl32) | |
| 8 | p1 | (T)12 | 12 | 71741 | 71752 | CDS (ycf3) | 18 | p1 | (A)10 | 10 | 109375 | 109384 | IGS | |
| 9 | p2 | (AT)6 | 12 | 73245 | 73256 | IGS | 19 | p1 | (A)10 | 10 | 109836 | 109845 | CDS (rps12) | |
| 10 | p1 | (A)11 | 11 | 79740 | 79750 | IGS | 20 | p2 | (AT)6 | 12 | 111746 | 111757 | IGS | |
| 1 | p1 | (C)13 | 13 | 15142 | 15154 | IGS | 11 | p1 | (T)11 | 11 | 87091 | 87101 | IGS | |
| 2 | p1 | (A)15 | 15 | 26051 | 26065 | IGS | 12 | p1 | (A)10 | 10 | 100620 | 100629 | IGS | |
| 3 | c | (A)11(G)10 | 21 | 30203 | 30223 | IGS | 13 | p1 | (T)10 | 10 | 103590 | 103599 | IGS | |
| 4 | p1 | (T)23 | 23 | 40998 | 41020 | IGS | 14 | p1 | (G)10 | 10 | 104157 | 104166 | CDS (ndhD) | |
| P. uncinata | 5 | p1 | (T)11 | 11 | 48131 | 48141 | IGS | 15 | p1 | (A)13 | 13 | 106942 | 106954 | IGS |
| 6 | p1 | (A)10 | 10 | 54428 | 54437 | IGS | 16 | p1 | (T)11 | 11 | 107349 | 107359 | CDS (rpl32) | |
| 7 | p1 | (A)10 | 10 | 67836 | 67845 | IGS | 17 | p1 | (A)11 | 11 | 109393 | 109403 | IGS | |
| 8 | p1 | (T)13 | 13 | 71760 | 71772 | CDS (ycf3) | 18 | p1 | (A)11 | 11 | 109855 | 109865 | CDS (rps12) | |
| 9 | p2 | (AT)6 | 12 | 73265 | 73276 | IGS | 19 | p2 | (AT)6 | 12 | 111767 | 111778 | IGS | |
| 10 | p1 | (A)12 | 12 | 79761 | 79772 | IGS | ||||||||
| 1 | p1 | (T)11 | 11 | 1376 | 1386 | IGS | 12 | p1 | (A)10 | 10 | 79947 | 79956 | IGS | |
| 2 | p1 | (A)10 | 10 | 9837 | 9846 | IGS | 13 | p1 | (T)10 | 10 | 87277 | 87286 | IGS | |
| 3 | c | (C)10(T)11 | 21 | 15195 | 15215 | IGS | 14 | p1 | (A)10 | 10 | 100844 | 100853 | IGS | |
| 4 | p1 | (A)12 | 12 | 26112 | 26123 | IGS | 15 | p1 | (T)11 | 11 | 101130 | 101140 | IGS | |
| P. sylvestris | 5 | c | (A)11(G)10 | 21 | 30269 | 30289 | IGS | 16 | p1 | (T)10 | 10 | 101833 | 101842 | CDS (ndhH) |
| 6 | p1 | (T)11 | 11 | 41059 | 41069 | IGS | 17 | p1 | (T)10 | 10 | 102658 | 102667 | IGS | |
| 7 | p1 | (T)19 | 19 | 45043 | 45061 | IGS | 18 | p1 | (G)11 | 11 | 104388 | 104398 | CDS (ndhD) | |
| 8 | p1 | (A)12 | 12 | 68030 | 68041 | IGS | 19 | p1 | (T)11 | 11 | 107567 | 107577 | CDS (rpl32) | |
| 9 | p1 | (T)14 | 14 | 71957 | 71970 | CDS (ycf3) | 20 | p1 | (A)10 | 10 | 109610 | 109619 | IGS | |
| 10 | p2 | (AT)6 | 12 | 73462 | 73473 | IGS | 21 | p1 | (A)12 | 12 | 110071 | 110082 | CDS (rps12) | |
| 11 | p2 | (AT)6 | 12 | 79134 | 79145 | IGS | 22 | p2 | (AT)7 | 14 | 111984 | 111997 | IGS |
| GenBank Accession | Taxon | Common Name | Family |
|---|---|---|---|
| NC_042410 | Abies alba | silver fir | Pinaceae |
| KP742350 | Abies koreana | Korean fir | Pinaceae |
| AB501189 | Larix decidua | common larch | Pinaceae |
| NC_036811 | Larix sibirica | Siberian larch | Pinaceae |
| NC_021456 | Picea abies | Norway spruce | Pinaceae |
| NC_032367 | Picea asperata | dragon spruce | Pinaceae |
| MN536531 | Pinus cembra | Swiss stone pine | Pinaceae |
| MK285358 | Pinus densiflora | Japanese red pine | Pinaceae |
| MZ333466 | Pinus mugo subsp. mugo | dwarf mountain pine | Pinaceae |
| MZ333465 | Pinus mugo subsp. rotundata | peat-bog pine | Pinaceae |
| MZ333464 | Pinus mugo subsp. uncinata | mountain pine | Pinaceae |
| NC_039585 | Pinus pinea | Italian stone pine | Pinaceae |
| NC_026302 | Pinus strobus | Eastern white pine | Pinaceae |
| KR476379 | Pinus sylvestris | Scots pine | Pinaceae |
| KY964286 | Pinus taeda | loblolly pine | Pinaceae |
| MH536745 | Podocarpus latifolius | broad-leaved yellowwood | Podocarpaceae |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokołowska, J.; Fuchs, H.; Celiński, K. New Insight into Taxonomy of European Mountain Pines, Pinus mugo Complex, Based on Complete Chloroplast Genomes Sequencing. Plants 2021, 10, 1331. https://doi.org/10.3390/plants10071331
Sokołowska J, Fuchs H, Celiński K. New Insight into Taxonomy of European Mountain Pines, Pinus mugo Complex, Based on Complete Chloroplast Genomes Sequencing. Plants. 2021; 10(7):1331. https://doi.org/10.3390/plants10071331
Chicago/Turabian StyleSokołowska, Joanna, Hanna Fuchs, and Konrad Celiński. 2021. "New Insight into Taxonomy of European Mountain Pines, Pinus mugo Complex, Based on Complete Chloroplast Genomes Sequencing" Plants 10, no. 7: 1331. https://doi.org/10.3390/plants10071331
APA StyleSokołowska, J., Fuchs, H., & Celiński, K. (2021). New Insight into Taxonomy of European Mountain Pines, Pinus mugo Complex, Based on Complete Chloroplast Genomes Sequencing. Plants, 10(7), 1331. https://doi.org/10.3390/plants10071331