
Identifying SSR Markers Related to Seed Fatty Acid Content in Perilla Crop (Perilla frutescens L.)


Abstract
:1. Introduction
2. Results
2.1. Fatty Acid Composition in Perilla Accessions
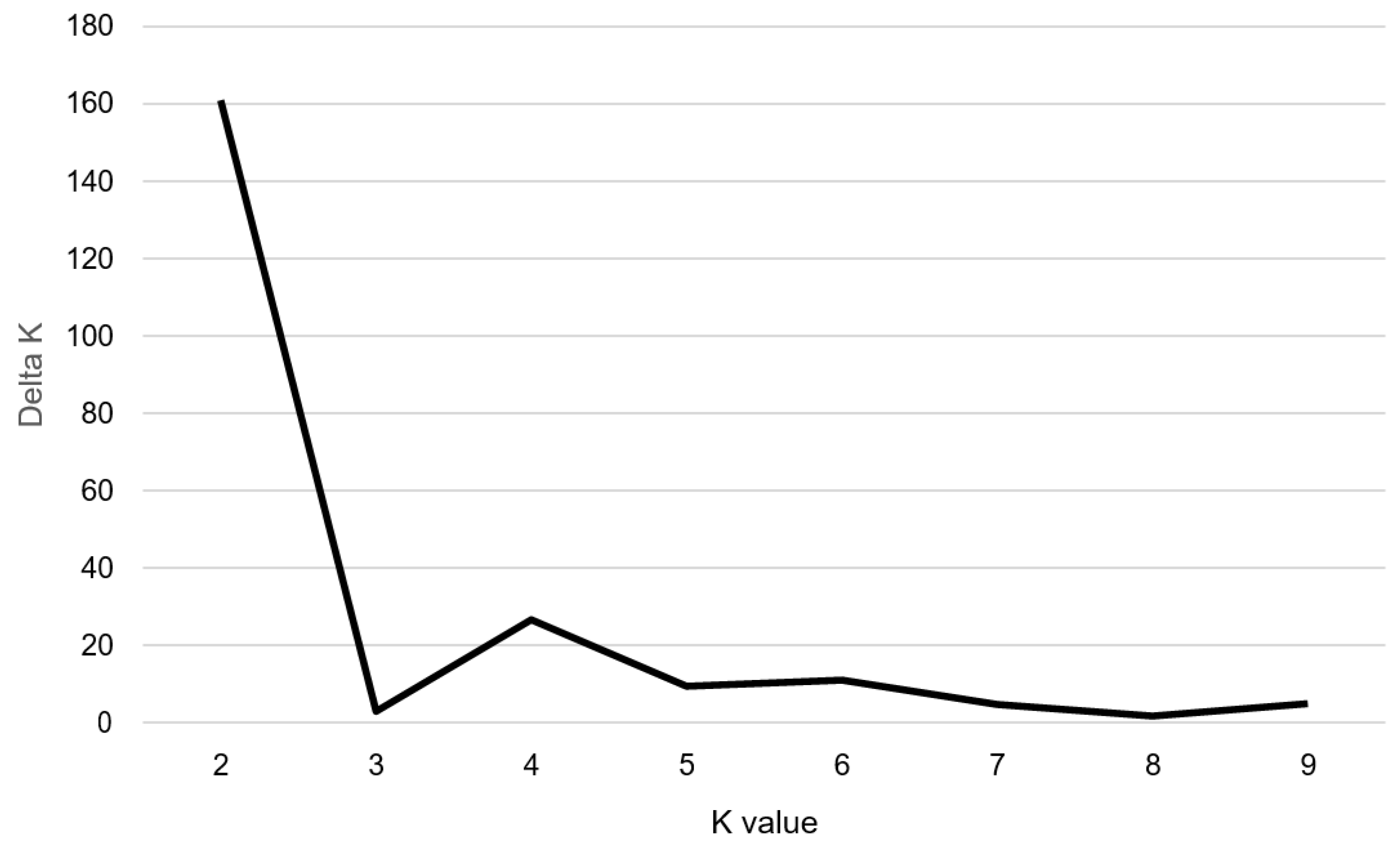
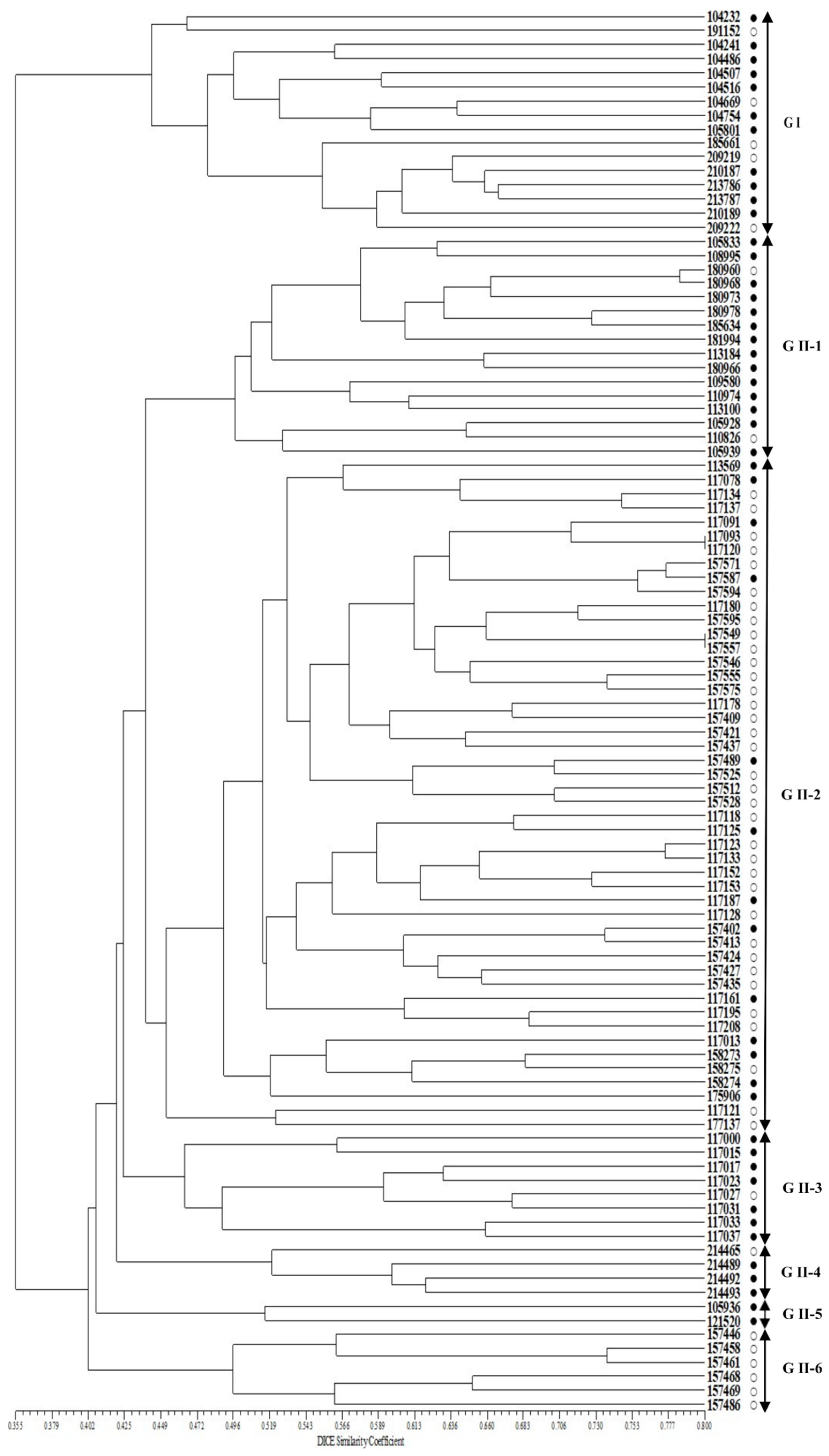
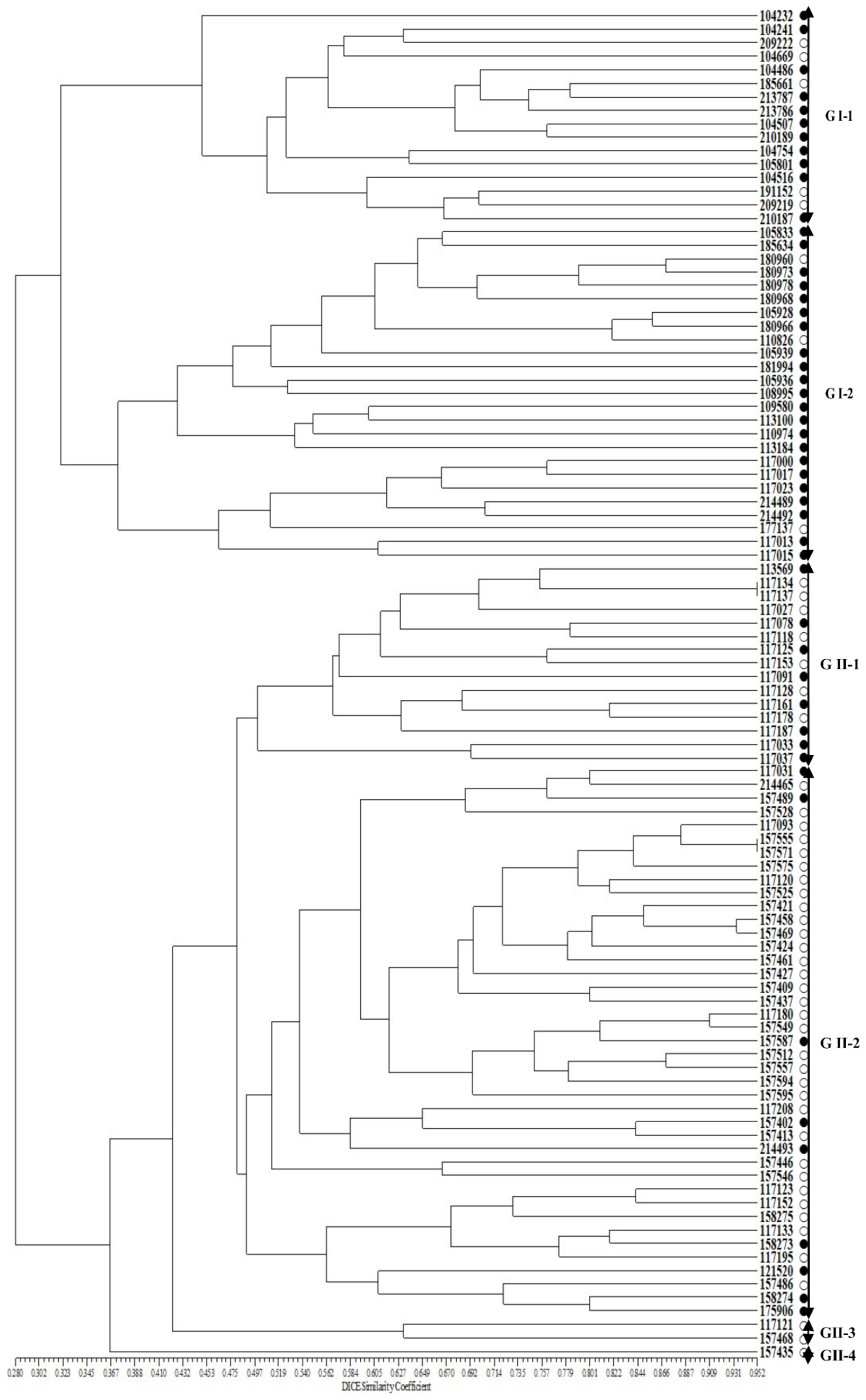
2.2. Genetic Variation in the Perilla Accessions Using SSR Markers
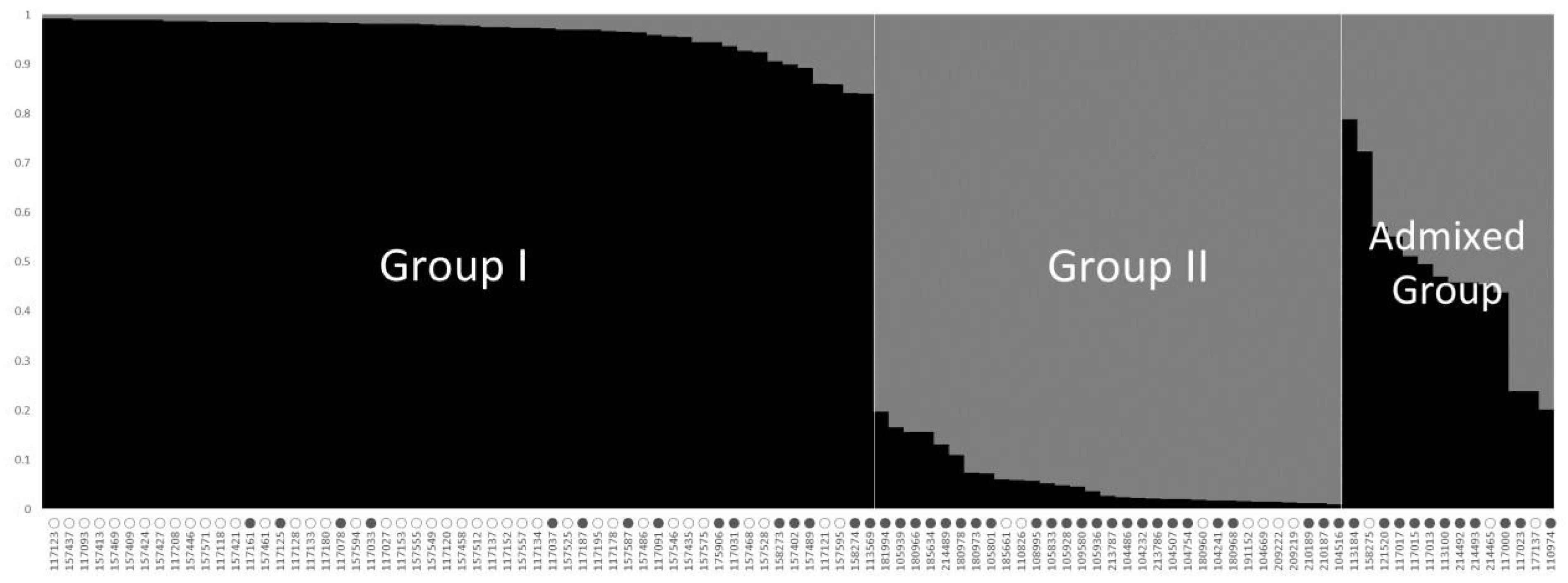
2.3. Population Structure and Association Analysis among 100 Perilla Accessions Using SSR Markers and Fatty Acid Content
2.4. Genetic Verification of SSR Markers among 100 Perilla Accessions
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Fatty Acid Composition
4.2. SSR Analysis and DNA Electrophoresis
4.3. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, J.K.; Ohnishi, O. Geographical differentiation of morphological characters among Perilla crops and their weedy types in East Asia. Breed Sci. 2001, 51, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.K.; Ohnishi, O. Genetic relationships among cultivated types of Perilla Frutescens and their weedy types in East Asia revealed by AFLP markers. Genet. Resour. Crop Evol. 2003, 50, 65–74. [Google Scholar] [CrossRef]
- Nitta, M.; Lee, J.K.; Ohnishi, O. Asian Perilla crops and their weedy forms: Their cultivation, utilization and genetic relationships. Econ. Bot. 2003, 57, 245–253. [Google Scholar] [CrossRef]
- Sa, K.J.; Choi, S.H.; Ueno, M.; Park, K.C.; Park, Y.J.; Ma, K.H.; Lee, J.K. Identification of genetic variations of cultivated and weedy types of Perilla species in Korea and Japan using morphological and SSR markers. Genes Genom. 2013, 35, 649–659. [Google Scholar] [CrossRef]
- Ma, S.J.; Sa, K.J.; Hong, T.K.; Lee, J.K. Genetic diversity and population structure analysis in Perilla crop and their weedy types from northern and southern areas of China based on simple sequence repeat (SSRs). Genes Genom. 2019, 41, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Makino, T. Makino’s New Illustrated Flora of Japan; Hokuryukan Co.: Tokyo, Japan, 1961. (In Japanese) [Google Scholar]
- Nitta, M. Origin Perilla Crops and Their Weedy Type. Ph.D. Thesis, Kyoto University, Kyoto, Japan, 2001; p. 78. [Google Scholar]
- Lee, J.I.; Han, E.D.; See, S.T.; Park, H.W. Study on the evaluation of oil quality and the differences of fatty acid composition between varieties in Perilla (Perilla frutescens Britton var. japonica Hara). Kor. J. Breed Sci. 1986, 18, 228–233. [Google Scholar]
- Lee, J.I.; Bang, J.K.; Lee, B.H.; Kim, K.H. Quality improvement in Perilla. I. Varietal differences of oil content and fatty acid composition. Kor. J. Crop Sci. 1991, 36, 48–61. [Google Scholar]
- Lee, J.; Lee, M.H.; Cho, E.J.; Lee, S. High-yield methods for purification of a-linolenic acid from Perilla frutescens var. japonica oil. Appl. Biol. Chem. 2016, 59, 89–94. [Google Scholar] [CrossRef]
- Shin, H.S.; Kim, S.W. Lipid composition of perilla seed. J. Am. Oil Chem. Soc. 1994, 71, 619–622. [Google Scholar] [CrossRef]
- Asif, M. Health effects of omega-3,6,9 fatty acids: Perilla frutescens is a good example of plant oils. Orient Pharm. Exp. Med. 2011, 11, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Ciftci, O.N.; Przybylski, R.; Rudzińska, M. Lipid components of flax, perilla, and chia seeds. Eur. J. Lipid Sci. Technol. 2012, 114, 794–800. [Google Scholar] [CrossRef]
- Okamoto, M.; Mitsunobu, F.; Ashida, K.; Mifune, T.; Hosaki, Y.; Tsugeno, H.; Harada, S.; Tanizaki, Y. Effects of dietary supplementation with n-3 fatty Acids compared with n-6 fatty acids on bronchial asthma. Intern Med. 2000, 39, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Talbott, S.M.; Hughes, K. Perilla seed oil (Perilla frutescens). In The Health Professional’s Guide to Dietary Supplements; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2006; pp. 258–260. [Google Scholar]
- Kopecky, J.; Rossmeisl, M.; Flachs, P.; Kuda, O.; Brauner, P.; Jilkova, Z.; Stankova, B.; Tvrzicka, E.; Bryhn, M. n-3 PUFA: Bioavailability and modulation of adipose tissue function. Proc. Nutr. Soc. 2009, 68, 361–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, G.L. Dietary n-6 and n-3 polyunsaturated fatty acids: From biochemistry to clinical implications in cardiovascular prevention. Biochem. Pharmacol. 2009, 77, 937–946. [Google Scholar] [CrossRef]
- Hashimoto, M.; Matsuzaki, K.; Kato, S.; Hossain, S.; Ohno, M.; Shido, O. Twelve-Month Studies on Perilla Oil Intake in Japanese Adults-Possible Supplement for Mental Health. Foods 2020, 9, 530. [Google Scholar] [CrossRef] [Green Version]
- Debener, T.; Salamini, E.; Gebhardt, C. Phylogeny of wild and cultivated Solanum species based on nuclear restriction fragment length polymorphisms (RFLPs). Theor. Appl. Genet. 1990, 79, 360–368. [Google Scholar] [CrossRef]
- Baird, E.; Coper-Bland, S.; Waugh, R.; DeMaine, M.; Powell, W. Molecular characterisation of interand intra-specific somatic hybrids of potato using randomly amplified polymorphic DNA (RAPD) markers. Mol. Gen. Genet. 1992, 233, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; Van De Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.J.; Lee, J.K.; Kim, N.S. Simple sequence repeat polymorphisms (SSRPs) for evaluation of molecular diversity and germplasm classification of minor crops. Molecules 2009, 14, 4546–4569. [Google Scholar] [CrossRef] [Green Version]
- Nasu, S.; Suzuki, J.; Ohta, R.; Hasegawa, K.; Yui, R.; Kitazawa, N.; Monna, L.; Minobe, Y. Search for and analysis of single nucleotide polymorphisms (SNPs) in rice (Oryza sativa, Oryza rufipogon) and establishment of SNP markers. DNA Res. 2002, 9, 163–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akagi, H.; Yokozaki, Y.; Inagaki, A.; Fujimura, T. Highly polymorphic microsatellites of rice consist of AT repeats, and a classification of closely related cultivars with these microsatellite loci. Theor. Appl. Genet. 1997, 94, 61–67. [Google Scholar] [CrossRef]
- Lee, J.K.; Nitta, M.; Kim, N.S.; Park, C.H.; Yoon, K.M.; Shin, Y.B.; Ohnishi, O. Genetic diversity of Perilla and related weedy types in Korea determined by AFLP analyses. Crop Sci. 2002, 42, 2161–2166. [Google Scholar] [CrossRef]
- Oh, J.K.; Sa, K.J.; Hyun, D.Y.; Cho, G.T.; Lee, J.K. Assessment of genetic diversity and population structure among a collection of Korean Perilla germplasms based on SSR markers. Genes Genom. 2020, 42, 1419–1430. [Google Scholar] [CrossRef]
- Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogelet, J.; Tingey, S.; Rafalski, A. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- Ball, R.D. Statistical analysis and experimental design. In Association Mapping in Plants; Oraguzie, N.C., Rikkerink, E.H.A., Gardiner, S.E., Eds.; Springer Science + Business Media LLC: New York, NY, USA, 2007; pp. 133–196. [Google Scholar]
- Gupta, P.K.; Kulwal, P.L.; Jaiswal, V. Association mapping in crop plants: Opportunities and challenges. In Advances in Genetics; Friedmann, T., Dunlap, J., Goodwin, S., Eds.; Academic Elsevier: Amsterdam, The Netherlands, 2014; Volume 85, pp. 109–148. [Google Scholar]
- Sa, K.J.; Park, J.Y.; Woo, S.Y.; Ramekar, R.V.; Jang, C.S.; Lee, J.K. Mapping of QTL traits in corn using a RIL population derived from a cross of dent corn x waxy corn. Genes Genom. 2015, 37, 1–14. [Google Scholar] [CrossRef]
- Choi, J.K.; Sa, K.J.; Park, D.H.; Lim, S.E.; Ryu, S.H.; Park, J.Y.; Park, K.J.; Rhee, H.I.; Lee, M.; Lee, J.K. Construction of genetic linkage map and identification of QTLs related to agronomic traits in DH. population of maize (Zea mays L.) using SSR markers. Genes Genom. 2019, 41, 667–678. [Google Scholar] [CrossRef]
- Kwon, S.J.; Lee, J.K.; Kim, N.S.; Yu, J.W.; Dixit, A.; Cho, E.G.; Park, Y.J. Isolation and characterization of SSR. markers in Perilla frutescens Britt. Mol. Eco. Notes 2005, 5, 454–456. [Google Scholar] [CrossRef]
- Park, Y.J.; Dixit, A.; Ma, K.H.; Lee, J.K.; Lee, M.H.; Chung, C.S.; Nitta, M.; Okuno, K.; Kim, T.S.; Cho, E.G.; et al. Evaluation of genetic diversity and relationships within an on-farm collection of Perilla frutescens (L.) Britt. using microsatellite markers. Genet. Resour. Crop Evol. 2008, 55, 523–535. [Google Scholar] [CrossRef]
- Sa, K.J.; Choi, I.K.; Park, K.C.; Lee, J.K. Genetic diversity and population structure among accessions of Perilla frutescens (L.) Britton in East Asia using new developed microsatellite markers. Genes Genom. 2018, 40, 1319–1329. [Google Scholar] [CrossRef] [PubMed]
- Sa, K.J.; Lim, S.E.; Choi, I.K.; Park, K.C.; Lee, J.K. Development and Characterization of New Microsatellite Markers for Perilla frutescens (L.) Britton. Amer. J. Plant Sci. 2019, 10, 1623–1630. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Sa, K.Y.; Ha, Y.J.; Lee, J.K. Genetic variation and association mapping in F2 population of Perilla crop (Perilla frutescens L.) using new developed Perilla SSR markers. Euphytica 2021, 217, 135. [Google Scholar] [CrossRef]
- Park, D.H.; SA, K.J.; Lim, S.E.; Ma, S.J.; Lee, J.K. Genetic diversity and population structure of Perilla frutescens collected from Korea and China based on simple sequence repeats (SSRs). Genes Genom. 2019, 41, 1329–1340. [Google Scholar] [CrossRef] [PubMed]
- Ha, Y.J.; Sa, K.J.; Lee, J.K. Identifying SSR markers associated with seed characteristics in Perilla (Perilla frutescens L.). Physiol. Mol. Biol. Plants 2021, 27, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.E.; Sa, K.J.; Lee, J.K. Bulk segregant analysis identifies SSR markers associated with leaf-and seed related traits in-Perilla crop (Perilla frutescens L.). Genes Genom. 2021, 43, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Li, H.L. The vegetables of ancient china. Econ. Bot. 1969, 23, 235–260. [Google Scholar] [CrossRef]
- Song, J.Y.; Lee, J.R.; Kim, C.Y.; Bae, C.H.; Lee, G.A.; Ma, K.H.; Choi, Y.M.; Park, H.J.; Lee, M.C. Assessment of Genetic Diversity and Fatty acid Composition of Perilla (Perilla frutescens var. frutescens) Germplasm. Kor. J. Plant Res. 2012, 25, 762–772. [Google Scholar] [CrossRef] [Green Version]
- Gunstone, F.D.; Harwood, J.; Padley, F.B. The Lipid Handbook, 2nd ed.; Chapman & Hall: London, UK, 1994. [Google Scholar]
- Ding, Y.; Neo, C.M.; Hu, Y.; Shi, L.; Ma, C.; Liu, Y.J. Characterization of fatty acid composition from five perilla seed oils in China and its relationship to annual growth temperature. J. Med. Plants Res. 2012, 6, 1645–1651. [Google Scholar]
- Gwari, G.; Lohani, H.; Haider, S.Z.; Bhandari, U.; Chauhan, N.; Rawat, D.S. Fatty acid and nutrient composition of perilla (Perilla frutescens L.) accessions collected from Uttarakhand. Inter. J. Phytop. 2014, 5, 379–382. [Google Scholar]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [Green Version]
- Dice, L.R. Measures of the amount of ecologic association between species. Ecology 1945, 26, 297–302. [Google Scholar] [CrossRef]
- Rohlf, F. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System; Version Exter Software: Setauket, NY, USA, 2000. [Google Scholar]
- Pritchard, J.K.; Wen, W. Documentation for STRUCTURE Software: Version. 2003. Available online: http://www.pritc.h.bsd.uchicago.Edu (accessed on 10 January 2021).
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software. STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession No. | Fatty Acid Content | |||||
|---|---|---|---|---|---|---|
| Total (%) | ||||||
| Palmitic Acid (%) | Stearic Acid (%) | Oleic Acid (%) | Linoleic Acid (%) | Linolenic Acid (%) | ||
| 104232 | 7.39 | 3.47 | 17.9 | 15.4 | 74.4 | 118.6 |
| 104241 | 7.38 | 3.19 | 19.0 | 15.4 | 71.7 | 116.7 |
| 104486 | 7.29 | 3.36 | 17.7 | 16.6 | 70.5 | 115.4 |
| 104507 | 7.28 | 3.13 | 19.6 | 20.9 | 65.9 | 116.8 |
| 104516 | 8.18 | 3.26 | 18.4 | 20.3 | 66.4 | 116.6 |
| 104669 | 7.57 | 3.10 | 14.3 | 14.8 | 57.9 | 97.6 |
| 104754 | 8.23 | 3.28 | 19.1 | 22.1 | 65.9 | 118.6 |
| 105801 | 8.91 | 3.33 | 18.5 | 21.0 | 69.7 | 121.5 |
| 105833 | 7.09 | 3.41 | 20.9 | 15.2 | 70.5 | 117.1 |
| 105928 | 8.76 | 3.24 | 18.7 | 18.3 | 69.8 | 118.8 |
| 105936 | 7.84 | 3.20 | 18.8 | 15.7 | 75.0 | 120.5 |
| 105939 | 7.41 | 2.56 | 17.1 | 20.2 | 72.2 | 119.4 |
| 108995 | 7.07 | 3.14 | 20.4 | 19.3 | 68.1 | 118.0 |
| 109580 | 7.43 | 3.34 | 18.2 | 17.0 | 72.4 | 118.4 |
| 110826 | 7.39 | 2.87 | 14.2 | 15.4 | 58.1 | 97.9 |
| 110974 | 7.79 | 3.30 | 18.5 | 14.2 | 75.1 | 118.9 |
| 113100 | 7.54 | 3.21 | 19.1 | 20.1 | 66.3 | 116.2 |
| 113184 | 8.53 | 3.29 | 17.8 | 21.0 | 68.6 | 119.2 |
| 113569 | 7.45 | 3.06 | 18.6 | 20.0 | 68.8 | 117.9 |
| 117000 | 7.02 | 3.77 | 21.2 | 13.5 | 70.8 | 116.3 |
| 117013 | 7.08 | 3.47 | 20.4 | 15.3 | 69.5 | 115.8 |
| 117015 | 7.05 | 3.88 | 19.0 | 10.2 | 75.4 | 115.5 |
| 117017 | 6.87 | 3.78 | 20.3 | 19.7 | 67.3 | 117.9 |
| 117023 | 7.28 | 3.70 | 19.6 | 12.6 | 72.9 | 116.0 |
| 117027 | 7.26 | 3.37 | 14.2 | 15.2 | 57.9 | 97.9 |
| 117031 | 7.49 | 3.95 | 21.3 | 12.0 | 71.0 | 115.7 |
| 117033 | 6.85 | 3.97 | 21.9 | 15.1 | 68.8 | 116.6 |
| 117037 | 7.37 | 3.79 | 21.3 | 14.5 | 69.1 | 116.0 |
| 117078 | 6.91 | 3.62 | 19.8 | 13.3 | 71.7 | 115.4 |
| 117091 | 7.19 | 3.28 | 18.1 | 20.6 | 70.3 | 119.5 |
| 117093 | 7.74 | 3.53 | 13.5 | 15.9 | 56.5 | 97.3 |
| 117118 | 7.18 | 3.06 | 15.3 | 15.8 | 56.9 | 98.2 |
| 117120 | 6.95 | 2.91 | 13.6 | 14.4 | 60.3 | 98.2 |
| 117121 | 7.33 | 2.96 | 11.9 | 14.5 | 61.0 | 97.7 |
| 117123 | 7.54 | 3.27 | 11.9 | 17.1 | 57.7 | 97.6 |
| 117125 | 7.53 | 3.71 | 19.2 | 16.7 | 68.0 | 115.2 |
| 117128 | 7.54 | 3.24 | 14.2 | 15.1 | 57.7 | 97.8 |
| 117133 | 7.76 | 3.38 | 11.3 | 15.6 | 59.1 | 97.1 |
| 117134 | 7.42 | 2.67 | 13.8 | 16.0 | 57.7 | 97.6 |
| 117137 | 7.67 | 2.95 | 12.5 | 17.7 | 56.6 | 97.5 |
| 117152 | 7.84 | 3.22 | 12.6 | 16.9 | 57.1 | 97.7 |
| 117153 | 7.67 | 3.28 | 14.8 | 17.7 | 54.3 | 97.7 |
| 117161 | 8.92 | 3.36 | 17.2 | 16.4 | 70.5 | 116.4 |
| 117178 | 7.85 | 3.33 | 16.5 | 12.6 | 57.6 | 98.0 |
| 117180 | 7.63 | 2.86 | 12.8 | 15.5 | 59.3 | 98.0 |
| 117187 | 7.28 | 3.25 | 18.7 | 13.6 | 72.5 | 115.4 |
| 117195 | 7.72 | 3.02 | 17.1 | 14.3 | 55.8 | 97.9 |
| 117208 | 7.59 | 3.03 | 16.8 | 14.2 | 56.1 | 97.7 |
| 121520 | 6.52 | 3.72 | 18.5 | 13.8 | 73.8 | 116.3 |
| 157402 | 7.26 | 2.95 | 17.3 | 20.0 | 67.9 | 115.4 |
| 157409 | 7.74 | 3.14 | 13.5 | 14.0 | 59.4 | 97.8 |
| 157413 | 7.47 | 3.21 | 15.6 | 15.2 | 56.3 | 97.8 |
| 157421 | 7.70 | 3.02 | 15.8 | 15.2 | 56.4 | 98.1 |
| 157424 | 7.61 | 3.22 | 14.7 | 17.1 | 55.2 | 97.8 |
| 157427 | 7.76 | 3.16 | 13.4 | 13.8 | 59.5 | 97.6 |
| 157435 | 7.78 | 3.18 | 13.1 | 14.4 | 59.2 | 97.6 |
| 157437 | 7.55 | 3.09 | 13.9 | 14.5 | 59.0 | 98.0 |
| 157446 | 7.46 | 3.11 | 13.1 | 16.5 | 57.4 | 97.6 |
| 157458 | 7.12 | 2.70 | 14.4 | 14.0 | 59.9 | 98.1 |
| 157461 | 7.12 | 2.67 | 13.8 | 14.3 | 60.3 | 98.3 |
| 157468 | 7.38 | 2.71 | 13.7 | 14.3 | 59.7 | 97.8 |
| 157469 | 6.00 | 1.90 | 12.0 | 21.2 | 58.8 | 99.9 |
| 157486 | 6.84 | 2.95 | 18.8 | 13.7 | 57.0 | 99.2 |
| 157489 | 7.10 | 3.99 | 19.2 | 12.6 | 74.4 | 117.3 |
| 157512 | 7.60 | 3.17 | 13.8 | 15.8 | 57.3 | 97.6 |
| 157525 | 7.19 | 3.11 | 14.8 | 14.3 | 58.5 | 97.9 |
| 157528 | 7.41 | 2.93 | 12.8 | 14.8 | 59.8 | 97.7 |
| 157546 | 7.06 | 2.64 | 13.1 | 14.4 | 60.4 | 97.6 |
| 157549 | 6.98 | 3.19 | 13.7 | 12.7 | 60.7 | 97.3 |
| 157555 | 7.18 | 2.82 | 14.4 | 13.8 | 59.3 | 97.5 |
| 157557 | 7.10 | 2.84 | 13.5 | 13.3 | 60.7 | 97.4 |
| 157571 | 7.09 | 2.71 | 12.6 | 15.1 | 60.6 | 98.1 |
| 157575 | 7.06 | 2.49 | 13.2 | 13.5 | 61.8 | 98.1 |
| 157587 | 9.13 | 3.29 | 17.7 | 23.4 | 71.3 | 124.8 |
| 157594 | 6.70 | 2.59 | 14.3 | 12.5 | 62.3 | 98.4 |
| 157595 | 6.90 | 2.55 | 12.4 | 14.8 | 61.5 | 98.2 |
| 158273 | 7.01 | 3.01 | 19.7 | 21.0 | 66.0 | 116.7 |
| 158274 | 8.52 | 3.12 | 15.6 | 20.2 | 68.7 | 116.1 |
| 158275 | 7.07 | 3.14 | 14.8 | 15.0 | 57.7 | 97.7 |
| 175906 | 7.52 | 3.06 | 16.8 | 17.7 | 72.5 | 117.6 |
| 177137 | 5.10 | 2.10 | 11.1 | 22.7 | 58.9 | 99.9 |
| 180960 | 6.89 | 2.81 | 16.5 | 14.4 | 57.2 | 97.8 |
| 180966 | 7.73 | 3.10 | 16.7 | 16.1 | 73.8 | 117.4 |
| 180968 | 6.79 | 3.69 | 19.2 | 17.5 | 68.6 | 115.8 |
| 180973 | 7.78 | 3.14 | 16.2 | 17.9 | 71.1 | 116.1 |
| 180978 | 8.73 | 3.12 | 16.0 | 19.2 | 70.9 | 117.9 |
| 181994 | 7.08 | 3.18 | 18.5 | 17.1 | 73.1 | 119.0 |
| 185634 | 8.03 | 3.24 | 17.7 | 21.8 | 66.3 | 117.1 |
| 185661 | 7.05 | 2.95 | 13.8 | 14.2 | 59.9 | 97.9 |
| 191152 | 7.08 | 3.03 | 14.8 | 14.3 | 58.2 | 97.4 |
| 209219 | 7.08 | 2.93 | 12.7 | 14.9 | 60.3 | 97.9 |
| 209222 | 6.20 | 1.70 | 15.3 | 17.0 | 59.7 | 99.9 |
| 210187 | 7.33 | 3.34 | 19.0 | 20.6 | 66.6 | 116.8 |
| 210189 | 7.62 | 3.35 | 16.1 | 14.0 | 74.7 | 115.8 |
| 213786 | 8.33 | 3.33 | 17.5 | 16.3 | 71.7 | 117.1 |
| 213787 | 7.57 | 3.01 | 18.5 | 12.7 | 74.8 | 116.6 |
| 214465 | 7.22 | 2.87 | 13.5 | 14.4 | 59.9 | 97.9 |
| 214489 | 7.58 | 3.13 | 17.3 | 18.0 | 71.1 | 117.1 |
| 214492 | 8.58 | 3.28 | 16.5 | 17.8 | 68.9 | 115.1 |
| 214493 | 7.14 | 3.67 | 17.9 | 13.7 | 73.3 | 115.7 |
| Mean | 7.43 ± 0.592 | 3.15 ± 0.39 | 16.3 ± 2.75 | 16.2 ± 2.76 | 64.6 ± 6.44 | 107.6 ± 9.79 |
| Max | 9.13 | 3.99 | 21.9 | 23.4 | 75.4 | 124.8 |
| Min | 5.10 | 1.70 | 11.1 | 10.2 | 54.3 | 97.1 |
| SA | OA | LA | LNA | TFAC | |
|---|---|---|---|---|---|
| PA | 0.315 ** | 0.123 | 0.246 * | 0.192 | 0.303 ** |
| SA | 0.619 ** | −0.157 | 0.477 ** | 0.503 ** | |
| OA | 0.141 | 0.724 ** | 0.829 ** | ||
| LA | 0.132 | 0.417 ** | |||
| LNA | 0.929 ** |
| SSR Loci | Allele No | GD | PIC | MAF |
|---|---|---|---|---|
| KNUPF1 | 5 | 0.332 | 0.315 | 0.810 |
| KNUPF2 | 9 | 0.757 | 0.717 | 0.450 |
| KNUPF3 | 6 | 0.625 | 0.585 | 0.630 |
| KNUPF4 | 5 | 0.709 | 0.655 | 0.820 |
| KNUPF5 | 6 | 0.687 | 0.633 | 0.620 |
| KNUPF10 | 9 | 0.743 | 0.718 | 0.280 |
| KNUPF11 | 4 | 0.313 | 0.293 | 0.550 |
| KNUPF14 | 5 | 0.594 | 0.544 | 0.700 |
| KNUPF16 | 9 | 0.798 | 0.769 | 0.710 |
| KNUPF19 | 4 | 0.329 | 0.310 | 0.650 |
| KNUPF23 | 4 | 0.502 | 0.457 | 0.570 |
| KNUPF25 | 6 | 0.637 | 0.575 | 0.280 |
| KNUPF26 | 3 | 0.398 | 0.353 | 0.810 |
| KNUPF28 | 5 | 0.486 | 0.450 | 0.320 |
| KNUPF35 | 4 | 0.635 | 0.579 | 0.670 |
| KNUPF36 | 4 | 0.665 | 0.618 | 0.490 |
| KNUPF37 | 4 | 0.661 | 0.602 | 0.750 |
| KNUPF39 | 7 | 0.695 | 0.662 | 0.690 |
| KNUPF43 | 5 | 0.645 | 0.574 | 0.560 |
| KNUPF53 | 11 | 0.807 | 0.783 | 0.520 |
| KNUPF55 | 9 | 0.759 | 0.720 | 0.500 |
| KNUPF58 | 3 | 0.462 | 0.414 | 0.470 |
| KNUPF59 | 5 | 0.543 | 0.457 | 0.490 |
| KNUPF62 | 6 | 0.612 | 0.570 | 0.370 |
| KNUPF71 | 4 | 0.639 | 0.568 | 0.430 |
| KNUPF72 | 9 | 0.771 | 0.740 | 0.430 |
| KNUPF74 | 3 | 0.516 | 0.453 | 0.310 |
| KNUPF77 | 3 | 0.663 | 0.589 | 0.320 |
| KNUPF78 | 4 | 0.665 | 0.603 | 0.700 |
| KNUPF80 | 7 | 0.794 | 0.764 | 0.570 |
| KNUPF82 | 6 | 0.697 | 0.643 | 0.570 |
| KNUPF85 | 4 | 0.565 | 0.516 | 0.460 |
| KNUPF89 | 12 | 0.824 | 0.802 | 0.370 |
| KNUPF107 | 6 | 0.568 | 0.540 | 0.640 |
| KNUPF112 | 8 | 0.588 | 0.567 | 0.370 |
| KNUPF113 | 10 | 0.847 | 0.830 | 0.440 |
| KNUPF117 | 4 | 0.617 | 0.566 | 0.270 |
| KNUPF118 | 4 | 0.446 | 0.383 | 0.400 |
| KNUPF121 | 4 | 0.461 | 0.424 | 0.610 |
| KNUPF125 | 5 | 0.516 | 0.463 | 0.270 |
| Mean | 5.8 | 0.614 | 0.570 | 0.522 |
| Total | 231 |
| Trait | SSR Marker | GLM |
|---|---|---|
| TFAC | KNUPF14 | *** |
| KNUPF62 | *** | |
| KNUPF72 | ** | |
| KNUPF85 | *** | |
| LNA | KNUPF14 | *** |
| KNUPF53 | ** | |
| KNUPF62 | *** | |
| KNUPF72 | ** | |
| KNUPF85 | *** | |
| LA | KNUPF125 | ** |
| OA | KNUPF62 | ** |
| KNUPF85 | ** | |
| PA | KNUPF10 | ** |
| KNUPF89 | *** | |
| SA | KNUPF10 | *** |
| KNUPF16 | ** | |
| KNUPF71 | *** | |
| KNUPF118 | ** |
| SSR Loci | Forward Primer | Reverse Primer | Repeat Motif |
|---|---|---|---|
| KNUPF1 | CTTGCAGCTGATCATTAAGCTA | TTTCTTGTGTGCTCTAACAACG | (AG)11 |
| KNUPF2 | GAAACCAAATTTCTTGTTCTTACA | CAAACGCAGACTCTTATCAATG | (AG)16 |
| KNUPF3 | TTCCTTGTAGTCATCTGATCCC | TGGAAATTAATTAAAGGGCTGA | (AG)16 |
| KNUPF4 | TTTCAAAAATCTTACCAACGCT | TTCGTTTTTGCATCTAATTATTCA | (AT)10 |
| KNUPF5 | TCCATCTCATCTCATTCAAACA | ATGGATCGGAAATCTAAAAACA | (AT)10 |
| KNUPF10 | GCTGATGGGACTACCCATAATA | AGGATCGGAACAATTATTAGCC | (AT)11 |
| KNUPF11 | TTGCAAGGTAAGATGATGATGA | TTGAGGATTGACAATGTTCGT | (CA)10 |
| KNUPF14 | AAATTCTCCCTCCACTCTTCAC | TGTTGGCTTTTTCAAATCTTTT | (CT)12 |
| KNUPF16 | CCTGTATCTCTCCCCGATAAAT | TGGATTTAATGCAGTTGAGTTG | (CT)22 |
| KNUPF19 | TCGAGGTTGAACAGATACAATG | TGATTAATTTCTTACGTACACTCCA | (TG)10 |
| KNUPF23 | TTGCAAGTTCTTGAATTGTGAC | CACTCCTTCCCTCCTCTTTAAT | (TG)11 |
| KNUPF25 | GCTTAGTGTGAGGAATTATGTAGGA | ACTCAGCATGCTTGAATTCTC | (AAG)12 |
| KNUPF26 | ATTTGAAATCGAAAAAGCAAAA | TGCAACCCTATTAGCAGTTTCT | (ACA)8 |
| KNUPF28 | CAACCTCTTAAGCCTTTGAACA | AATGTGACGGGTTCTGTAAATC | (AGA)8 |
| KNUPF35 | CTCTTTCCTTCTCATTCACCAC | CCCTTTTTCTTACCCACTCTCT | (CTT)10 |
| KNUPF36 | GGGAGACGAGATAACACATGAT | TGCATACTCGATTGAAAGAAGA | (GCT)8 |
| KNUPF37 | GGTGTGAAAAAGAGAGTGGAGA | TTGAATTGCCTGTTGATAGTGA | (GGT)10 |
| KNUPF39 | TCACCTTCCCCTTCATTTATTA | AGGATCGAACAGAACAAACTGT | (TCT)13 |
| KNUPF43 | GTCAAATGAAATTCACACATTTTA | GTAAATGGGAATTTTTGAGGAG | (AT)7 |
| KNUPF53 | GATTCATCATTCAGCTCTCTCC | ATGACCAATGGATTAAACAAGG | (CT)17 |
| KNUPF55 | TGCTGTTGATGACTTGTATGGT | ATGAGATTTGGCTTCACAGAGT | (AGC)7 |
| KNUPF58 | GTATATGTGTGGGAAGGTTGCT | TCAATTTCCTCATCAAATCAAA | (ATG)7 |
| KNUPF59 | AATCTCGATGCCTAACAACAGT | TTCCTTGTAAATCCAGCTAAGG | (CAG)7 |
| KNUPF62 | CCATCCTTCTTGTTCAACTCAT | AATGTTGATGAGGAGACGTTTT | (CAT)7 |
| KNUPF71 | GAAGAATGCATCAGTAACACGA | ATGCTGGCCAAGTAATAAGAGA | (AGCT)4 |
| KNUPF72 | TAATTTGAGGGATTCCTTTCCT | CGCCACCCTTACTACTTCATAC | (TCGA)4 |
| KNUPF74 | TTGACTGTACCAGAGCATCAAG | GGGTACACTCACAACTCTACCAA | (AAAT)6 |
| KNUPF77 | TTTTTGGTTGCTTTTTCTTGAT | AGCAGATAAAATGTGCTGGATT | (TATG)10 |
| KNUPF78 | GCGTTATTATTTTTCAAGATCG | TCAATGATTTTACAGAAGATGCT | (AAC)7 |
| KNUPF80 | GATTCATCATTCAGCTCTCTCC | ATGACCAATGGATTAAACAAGG | (CT)17 |
| KNUPF82 | AAACCAAGGAACTCGTCAACTA | CGCTTCGTCTTTATTGTGTGTA | (AGA)7 |
| KNUPF85 | GATGACGATGAAGCTTTTCAG | CTCCTCAGCAGTTTCACCTAAC | (GAT)7 |
| KNUPF89 | ATATCCCATTTTCTGATGCAAG | CCTTTTTCTTGCCTACTCATCA | (AGTG)6 |
| KNUPF107 | TAAGGTTGCCGATTGTACTTTT | ACAGAATGGCTCACAAGGATAG | (AG)25 |
| KNUPF112 | AGTTGGAGTGGTTAAACTTGGA | CACGCACACTCCAATACTACAA | (AG)17 |
| KNUPF113 | GTAGGCATCCCTTTTTCTTCC | TTCAAGTTTACTCTTCACCGCT | (CT)17 |
| KNUPF117 | GTTGACCGATTTAGTTGGTTGT | TTTACACTGGTGACCGTCATT | (ATA)7 |
| KNUPF118 | CTAAAGATCATTGTGGAGGGAA | ACTCCATGAAATCCAACTCATC | (ATG)7 |
| KNUPF121 | ACATACAAATGTCTTTCCTGGG | AAGTAAGACATGGGAATTGGTG | (TCT)7 |
| KNUPF125 | TGTCCTTATTTTTCGTCGTCTT | TTCATAGAAACCTATGCCCTTG | (AAAG)4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.; Sa, K.J.; Hyun, D.Y.; Lee, S.; Lee, J.K. Identifying SSR Markers Related to Seed Fatty Acid Content in Perilla Crop (Perilla frutescens L.). Plants 2021, 10, 1404. https://doi.org/10.3390/plants10071404
Park H, Sa KJ, Hyun DY, Lee S, Lee JK. Identifying SSR Markers Related to Seed Fatty Acid Content in Perilla Crop (Perilla frutescens L.). Plants. 2021; 10(7):1404. https://doi.org/10.3390/plants10071404
Chicago/Turabian StylePark, Hyeon, Kyu Jin Sa, Do Yoon Hyun, Sookyeong Lee, and Ju Kyong Lee. 2021. "Identifying SSR Markers Related to Seed Fatty Acid Content in Perilla Crop (Perilla frutescens L.)" Plants 10, no. 7: 1404. https://doi.org/10.3390/plants10071404
APA StylePark, H., Sa, K. J., Hyun, D. Y., Lee, S., & Lee, J. K. (2021). Identifying SSR Markers Related to Seed Fatty Acid Content in Perilla Crop (Perilla frutescens L.). Plants, 10(7), 1404. https://doi.org/10.3390/plants10071404