
Water Use Efficiency in Popcorn (Zea mays L. var. everta): Which Physiological Traits Would Be Useful for Breeding?

 ,
,  ,
,  ,
,  , , ,
, , ,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Effect of Water Restriction, Comparison between Genotypic Groups and Heterosis Estimates
2.1.1. Morphological Characteristics
2.1.2. Physiological Characteristics
2.1.3. Root Traits
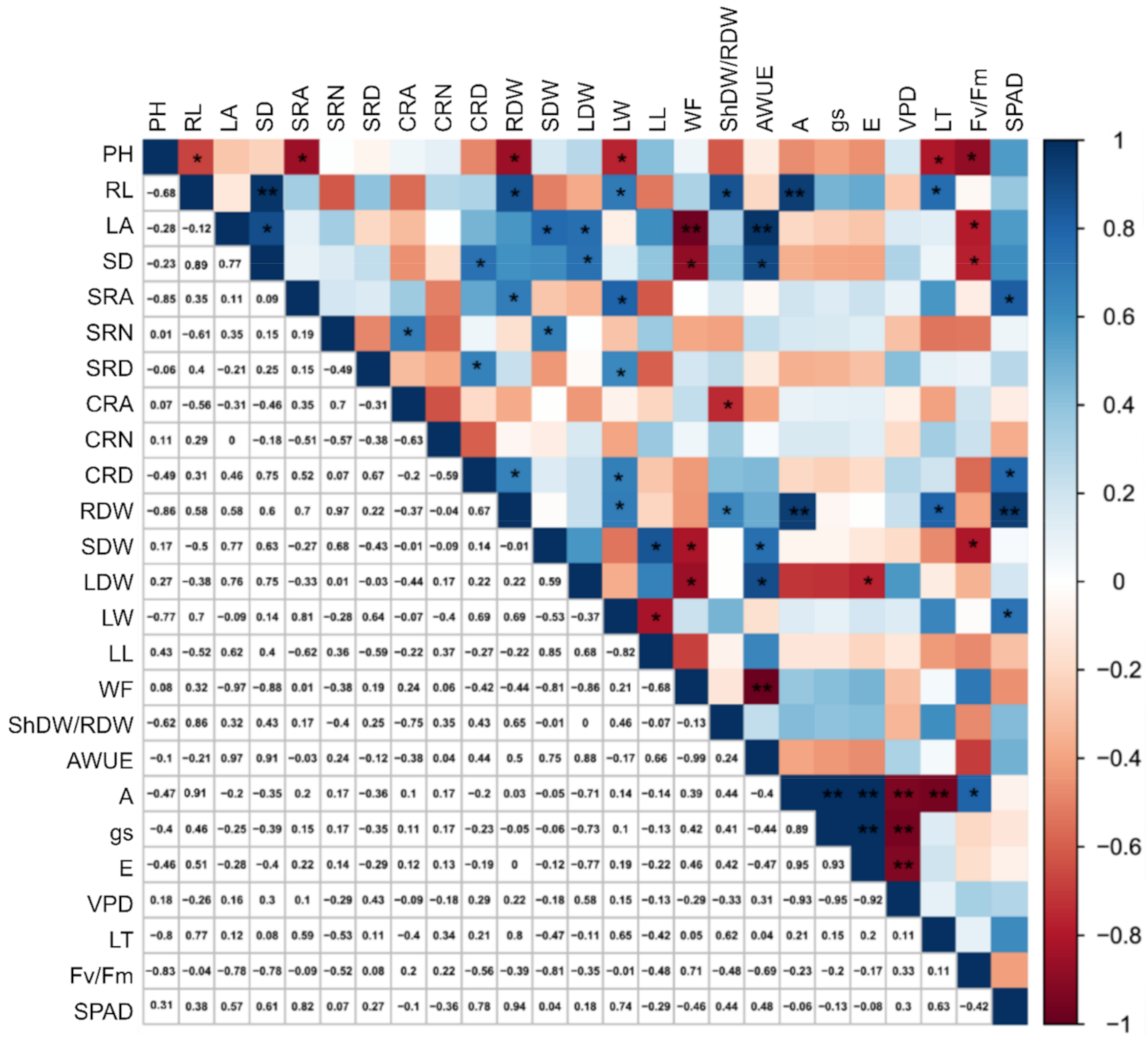
2.2. Genotypic Correlations
3. Discussion
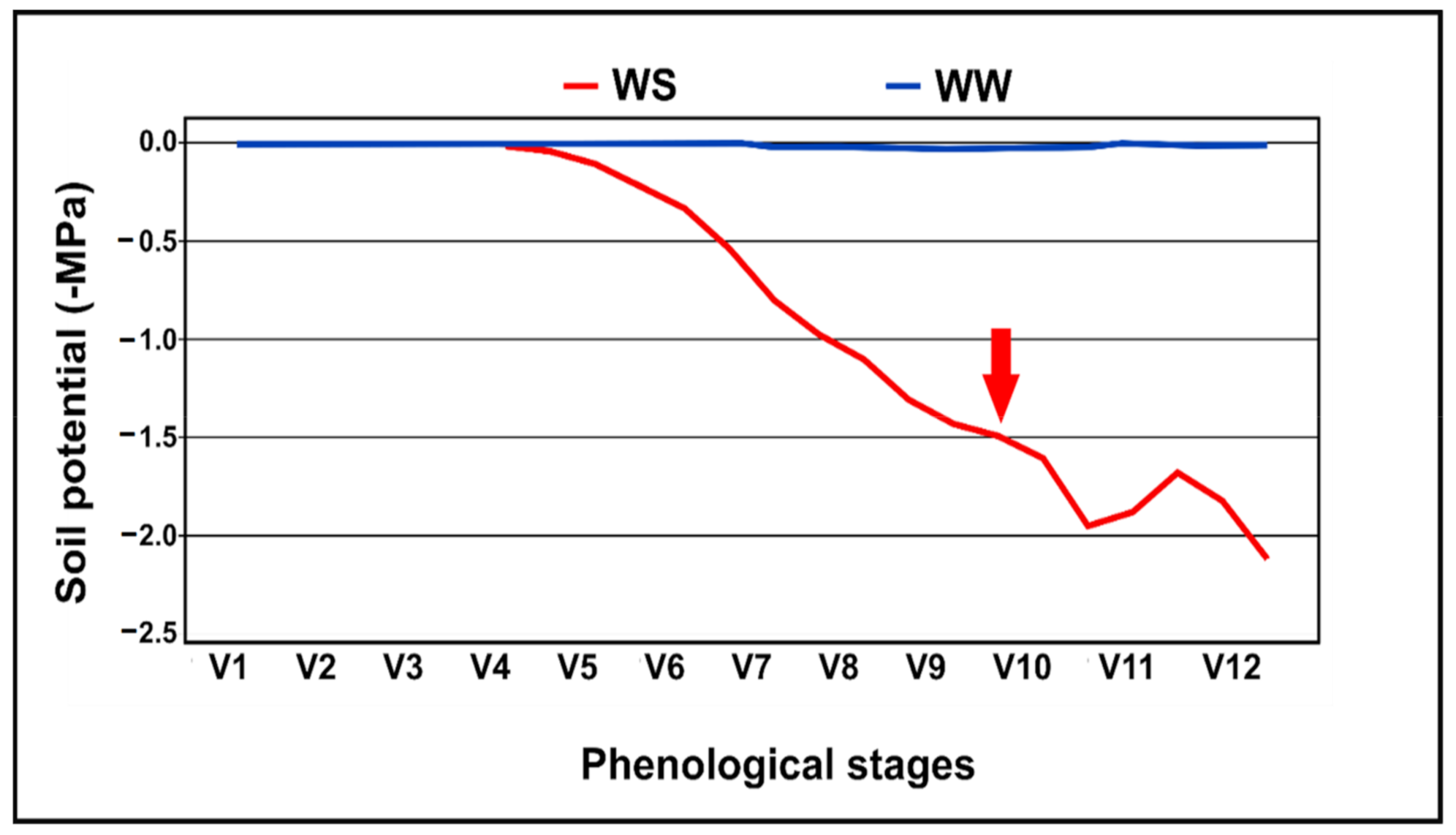
4. Materials and Methods
4.1. Study Traits
4.1.1. Morphological Traits
4.1.2. Physiological Traits
4.1.3. Root Traits
4.2. Statistical Analyses
4.3. Heterosis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014.
- Fisher, M.; Abate, T.; Lunduka, R.W.; Asnake, W.; Alemayehu, Y.; Madulu, R.B. Drought tolerant maize for farmer adaptation to drought in sub-Saharan Africa: Determinants of adoption in eastern and southern Africa. Clim. Chang. 2015, 133, 283–299. [Google Scholar] [CrossRef] [Green Version]
- Knutti, R.; Rogelj, J.; Sedlácek, J.; Fischer, E.M. A scientific critique of the two-degree climate change target. Nat. Geosci. 2016, 9, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Simova-Stoilova, L.; Vassileva, V.; Feller, U. Selection and Breeding of Suitable Crop Genotypes for Drought and Heat Periods in a Changing Climate: Which Morphological and Physiological Properties Should Be Considered? Agriculture 2016, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- Lunduka, R.W.; Mateva, K.I.; Magorokosho, C.; Manjeru, P. Impact of adoption of drought-tolerant maize varieties on total maize production in south Eastern Zimbabwe. Clim. Dev. 2019, 11, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Aslam, M.; Maqbool, M.A.; Cengiz, R. Drought Stress in Maize (Zea mays L.); Springer International Publishing: Faisalabad, Pakistan, 2015; pp. 5–17. [Google Scholar]
- Sah, R.P.; Chakraborty, M.; Prasad, K.; Pandit, M.; Tudu, V.K.; Chakravarty, M.K.; Narayan, S.C.; Rana, M.; Moharana, D. Impact of water deficit stress in maize: Phenology and yield components. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zia, S.; Romano, G.; Spreer, W.; Sanchez, C.; Cairns, J.; Araus, J.L.; Müller, J. Infrared Thermal Imaging as a Rapid Tool for Identifying Water-Stress Tolerant Maize Genotypes of Different Phenology. J. Agron. Crop Sci. 2013, 199, 75–84. [Google Scholar] [CrossRef]
- Song, Y.; Linderholm, H.W.; Luo, Y.; Xu, J.; Zhou, G. Climatic causes of maize production loss under global warming in Northeast China. Sustainability 2020, 12, 7829. [Google Scholar] [CrossRef]
- Kamphorst, S.H.; do Amaral Júnior, A.T.; de Lima, V.J.; Guimarães, L.J.M.; Schmitt, K.F.M.; Leite, J.T.; Santos, P.H.A.D.; Chaves, M.M.; Mafra, G.S.; dos Santos Junior, D.R.; et al. Can Genetic Progress for Drought Tolerance in Popcorn Be Achieved by Indirect Selection? Agronomy 2019, 9, 792. [Google Scholar] [CrossRef] [Green Version]
- Cairns, J.E.; Hellin, J.; Sonder, K.; Araus, J.L.; MacRobert, J.F.; Thierfelder, C.; Prasanna, B.M. Adapting maize production to climate change in sub-Saharan Africa. Food Secur. 2013, 5, 345–360. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Linderholm, H.W.; Song, Y.; Wang, F.; Liu, Y.; Tian, J.; Xu, J.; Song, Y.; Ren, G. Impacts of drought on maize and soybean production in northeast China during the past five decades. Int. J. Environ. Res. Public Health 2020, 17, 2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Qin, F. Genetic dissection of maize drought tolerance for trait improvement. Mol. Breed. 2021, 41, 8. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, R.; Chen, J.; Fan, Z. Using adaptive cross approximation for efficient calculation of monostatic scattering with multiple incident angles. Appl. Comput. Electromagn. Soc. J. 2011, 26, 325–333. [Google Scholar]
- Fathi, A.; Tari, D.B. Effect of Drought Stress and its Mechanism in Plants. Int. J. Life Sci. 2016, 10, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Binott, J.J.; Owuoche, J.O.; Bartels, D. Physiological and molecular characterization of Kenyan barley (Hordeum vulgare L.) seedlings for salinity and drought tolerance. Euphytica 2017, 213, 1–23. [Google Scholar] [CrossRef]
- Noor, J.J.; Vinayan, M.T.; Umar, S.; Devi, P.; Iqbal, M.; Seetharam, K.; Zaidi, P.H. Morpho-physiological traits associated with heat stress tolerance in tropical maize (Zea mays L.) at the reproductive stage. Aust. J. Crop Sci. 2019, 13, 536–545. [Google Scholar] [CrossRef]
- Kamphorst, S.H.; De Lima, V.J.; Leite, J.T.; Carvalho, C.M.; Xavier, K.B.; Campostrini, E. Popcorn breeding for water-stress tolerance or for agronomic water-use efficiency? Genet. Mol. Res. 2018, 17, 1–18. [Google Scholar] [CrossRef]
- Trachsel, S.; Kaeppler, S.M.; Brown, K.M.; Lynch, J.P. Shovelomics: High throughput phenotyping of maize (Zea mays L.) root architecture in the field. Plant Soil 2011, 341, 75–87. [Google Scholar] [CrossRef]
- Witcombe, J.R.; Hollington, P.A.; Howarth, C.J.; Reader, S.; Steele, K.A. Breeding for abiotic stresses for sustainable agriculture. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 703–716. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Lynch, J.P. Reduced crown root number improves water acquisition under water deficit stress in maize (Zea mays L.). J. Exp. Bot. 2016, 67, 4545–4557. [Google Scholar] [CrossRef] [Green Version]
- Qi, D.; Hu, T.; Liu, T. Biomass accumulation and distribution, yield formation and water use efficiency responses of maize (Zea mays L.) to nitrogen supply methods under partial root-zone irrigation. Agric. Water Manag. 2020, 230, 105981. [Google Scholar] [CrossRef]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamphorst, S.H.; de Lima, V.J.; Schimitt, K.F.M.; Leite, J.T.; Azeredo, V.C.; Pena, G.F.; Santos, P.H.A.D.; Santos Júnior, D.R.; da Silva Júnior, S.B.; Bispo, R.B.; et al. Water stress adaptation of popcorn roots and association with agronomic traits. Genet. Mol. Res. 2018, 17, 1–14. [Google Scholar] [CrossRef]
- De Lima, V.J.; Do Amaral Júnior, A.T.; Kamphorst, S.H.; Bispo, R.B.; Leite, J.T.; De Oliveira Santos, T.; Medeiros Schmitt, K.F.; Chaves, M.M.; De Oliveira, U.A.; Araújo Diniz Santos, P.H.; et al. Combined dominance and additive gene effects in trait inheritance of drought-stressed and full irrigated popcorn. Agronomy 2019, 9, 782. [Google Scholar]
- Liu, Y.; Subhash, C.; Yan, J.; Song, C.; Zhao, J.; Li, J. Maize leaf temperature responses to drought: Thermal imaging and quantitative trait loci (QTL) mapping. Environ. Exp. Bot. 2011, 71, 158–165. [Google Scholar] [CrossRef]
- Tollenaar, M.; Lee, E.A. Dissection of physiological processes underlying grain yield in maize by examining genetic improvement and heterosis. Maydica 2006, 51, 399–408. [Google Scholar]
- Tollenaar, M.; Ahmadzadeh, A.; Lee, E.A. Physiological Basis of Heterosis for Grain Yield in Maize. Crop Sci. 2004, 44, 2086. [Google Scholar] [CrossRef]
- Ali, M.L.; Luetchens, J.; Singh, A.; Shaver, T.M.; Kruger, G.R.; Lorenz, A.J. Greenhouse screening of maize genotypes for deep root mass and related root traits and their association with grain yield under water-deficit conditions in the field. Euphytica 2016, 207, 79–94. [Google Scholar] [CrossRef]
- Holá, D.; Benešová, M.; Fischer, L.; Haisel, D.; Hnilička, F.; Hniličková, H.; Jedelský, P.L.; Kočová, M.; Procházková, D.; Rothová, O.; et al. The disadvantages of being a hybrid during drought: A combined analysis of plant morphology, physiology and leaf proteome in maize. PLoS ONE 2017, 12, e0176121. [Google Scholar] [CrossRef]
- Zhao, J.; Xue, Q.; Jessup, K.E.; Hou, X.; Hao, B.; Marek, T.H.; Xu, W.; Evett, S.R.; O’Shaughnessy, S.A.; Brauer, D.K. Shoot and root traits in drought tolerant maize (Zea mays L.) hybrids. J. Integr. Agric. 2018, 17, 1093–1105. [Google Scholar] [CrossRef]
- Araus, J.L.; Sánchez, C.; Cabrera-Bosquet, L. Is heterosis in maize mediated through better water use? New Phytol. 2010, 187, 392–406. [Google Scholar] [CrossRef]
- Chairi, F.; Elazab, A.; Sanchez-Bragado, R.; Araus, J.L.; Serret, M.D. Heterosis for water status in maize seedlings. Agric. Water Manag. 2016, 164, 100–109. [Google Scholar] [CrossRef]
- Goff, S.A. A unifying theory for general multigenic heterosis: Energy efficiency, protein metabolism, and implications for molecular breeding. New Phytol. 2011, 189, 923–937. [Google Scholar] [CrossRef]
- Ryan, A.C.; Dodd, I.C.; Rothwell, S.A.; Jones, R.; Tardieu, F.; Draye, X.; Davies, W.J. Gravimetric phenotyping of whole plant transpiration responses to atmospheric vapour pressure deficit identifies genotypic variation in water use efficiency. Plant Sci. 2016, 251, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Nemali, K.S.; Bonin, C.; Dohleman, F.G.; Stephens, M.; Reeves, W.R.; Nelson, D.E.; Castiglioni, P.; Whitsel, J.E.; Sammons, B.; Silady, R.A.; et al. Physiological responses related to increased grain yield under drought in the first biotechnology-derived drought-tolerant maize. Plant Cell Environ. 2015, 38, 1866–1880. [Google Scholar] [CrossRef] [Green Version]
- Ali, Q.; Ali, A.; Waseem, M.; Muzaffar, A.; Ahmed, S.; Ali, S.; Bajwa, K.S.; Awan, M.F.; Samiullah, T.R.; Nasir, I.A.; et al. Correlation analysis for morpho-physiological traits of maize (Zea mays L.). Life Sci. J. 2014, 11, 9–13. [Google Scholar]
- Li, C.; Tao, Z.; Liu, P.; Zhang, J.; Zhuang, K.; Dong, S.; Zhao, M. Increased grain yield with improved photosynthetic characters in modern maize parental lines. J. Integr. Agric. 2015, 14, 1735–1744. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Escalona, J.M.; Sampol, B.; Medrano, H. Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. Funct. Plant Biol. 2002, 29, 461–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawlor, D.W. Limitation to photosynthesis in water-stressed leaves: Stomata vs. Metabolism and the role of ATP. Ann. Bot. 2002, 89, 871–885. [Google Scholar] [CrossRef] [PubMed]
- Busch, F.; Hüner, N.P.A.; Ensminger, I. Biochemical constrains limit the potential of the photochemical reflectance index as a predictor of effective quantum efficiency of photosynthesis during the winter spring transition in Jack pine seedlings. Funct. Plant Biol. 2009, 36, 1016. [Google Scholar] [CrossRef]
- Liu, Z.; Zhao, Y.; Guo, S.; Cheng, S.; Guan, Y.; Cai, H.; Mi, G.; Yuan, L.; Chen, F. Enhanced crown root number and length confers potential for yield improvement and fertilizer reduction in nitrogen-efficient maize cultivars. F. Crop. Res. 2019, 241, 107562. [Google Scholar] [CrossRef]
- Gowda, V.R.P.; Henry, A.; Yamauchi, A.; Shashidhar, H.E.; Serraj, R. Root biology and genetic improvement for drought avoidance in rice. F. Crop. Res. 2011, 122, 1–13. [Google Scholar] [CrossRef]
- Iqbal, K.; Azhar, F.M.; Khan, I.A. Ehsan-Ullah Variability for drought tolerance in cotton (Gossypium hirsutum) and its genetic basis. Int. J. Agric. Biol. 2011, 13, 61–66. [Google Scholar]
- Akinwale, R.O.; Awosanmi, F.E.; Ogunniyi, O.O.; Fadoju, A.O. Determinants of drought tolerance at seedling stage in early and extra-early maize hybrids. Maydica 2017, 62, 1. [Google Scholar]
- Kamphorst, S.H.; Gonçalves, G.M.B.; Do Amaral, A.T.; De Lima, V.J.; Leite, J.T.; Schmitt, K.F.M.; Dos Santos, D.R.; Santos, J.S.; De Oliveira, F.T.; Corrêa, C.C.G.; et al. Screening of popcorn genotypes for drought tolerance using canonical correlations. Agronomy 2020, 10, 1519. [Google Scholar] [CrossRef]
- Hussain, M.; Latif, A.; Hassan, W.; Farooq, S.; Hussain, S.; Ahmad, S.; Nawaz, A. Maize hybrids with well-developed root system perform better under deficit supplemental irrigation. Soil Environ. 2019, 38, 203–213. [Google Scholar] [CrossRef]
- Singh, V.; van Oosterom, E.J.; Jordan, D.R.; Messina, C.D.; Cooper, M.; Hammer, G.L. Morphological and architectural development of root systems in sorghum and maize. Plant Soil 2010, 333, 287–299. [Google Scholar] [CrossRef]
- Hauck, A.L.; Novais, J.; Grift, T.E.; Bohn, M.O. Characterization of mature maize (Zea mays L.) root system architecture and complexity in a diverse set of Ex-PVP inbreds and hybrids. Springerplus 2015, 4, 424. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, E.R.; Ferreira, V.M.; Silva, J.V.; Endres, L.; Barbosa, T.P.; Duarte, W.D.G. Trocas gasosas e fluorescência da clorofila a em variedades de cana-de-açúcar submetidas à deficiência hídrica 1 Gas exchange and chlorophyll a fluorescence of sugarcane varieties submitted to water stress. Rev. Bras. Eng. Agrícola Ambient. 2010, 14, 378–386. [Google Scholar] [CrossRef] [Green Version]
- Qi, W.Z.; Liu, H.H.; Liu, P.; Dong, S.T.; Zhao, B.Q.; So, H.B.; Li, G.; De Liu, H.; Zhang, J.W.; Zhao, B. Morphological and physiological characteristics of corn (Zea mays L.) roots from cultivars with different yield potentials. Eur. J. Agron. 2012, 38, 54–63. [Google Scholar] [CrossRef]
- Kamphorst, S.H.; do Amaral Júnior, A.T.; de Lima, V.J.; Santos, P.H.A.D.; Rodrigues, W.P.; Vivas, J.M.S.; Gonçalves, G.M.B.; Schmitt, K.F.M.; Leite, J.T.; Vivas, M.; et al. Comparison of Selection Traits for Effective Popcorn (Zea mays L. var. Everta) Breeding Under Water Limiting Conditions. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Brugnera, A.; Von Pinho, R.G.; Pacheco, C.A.P.; Alvarez, C.G.D. Resposta de cultivares de milho-pipoca a doses de adubação de semeadura. Rev. Ceres 2003, 50, 417–429. [Google Scholar]
- Albuquerque, P.E.P.; Resende, M. Cultivo do Milho. EMBRAPA milho e sorgo 2015, 3, 1–22. [Google Scholar]
- Cruz, C.D. GENES-A software package for analysis in experimental statistics and quantitative genetics. Acta Sci. Agron. 2013, 35, 271–276. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Combined Analysis | Water-Stressed | Well-Watered | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gen | WR | Gen * WR | WUEL | WUIL | C1 | HY | C2 | HET | WUEL | WUIL | C1 | HY | C2 | HET | |
| PH | ** | ** | * | 79.12 ± 4.20 | 68.8 ± 1.80 | ** | 78.9 ± 5.36 | ns | 7.87 | 147.5 ± 18.54 | 116.2 ± 7.58 | ** | 136.10 ± 14.00 | ns | 3.69 |
| LA | ** | ** | ** | 0.22 ± 0.01 | 0.21 ± 0.01 | ns | 0.27 ± 0.01 | ** | 4.89 | 0.36 ± 0.01 | 0.28 ± 0.01 | ns | 0.44 ± 0.02 | ** | 18.10 |
| SDW | ** | ** | ** | 7.00 ± 1.48 | 12.01 ± 2.16 | * | 13.80 ± 0.69 | ns | 20.8 | 31.32 ± 2.42 | 25.53 ± 5.48 | ** | 33.32 ± 5.19 | *** | 42.60 |
| LDW | ** | ** | * | 13.35 ± 1.29 | 12.41 ± 1.94 | ns | 15.56 ± 1.93 | *** | −5.05 | 17.05 ± 2.21 | 15.14 ± 2.85 | ns | 22.93 ± 2.89 | *** | 7.09 |
| ShDW/RDW | ** | ** | ns | 4.03 ± 0.03 | 4.15 ± 0.03 | ** | 4.50 ± 0.03 | ** | 7.31 | 3.66 ± 0.08 | 4.6 ± 0.06 | * | 4.43 ± 0.06 | * | 18.01 |
| LW | ** | ** | ns | 16.67 ± 1.25 | 17.99 ± 1.62 | ** | 4.60 ± 9.06 | ** | 13.4 | 36.59 ± 6.53 | 44.49 ± 8.30 | ** | 43.41 ± 6.87 | ns | 14.50 |
| LL | ** | ** | * | 92.55 ± 6.27 | 70.1 ± 2.76 | *** | 92.01 ± 4.86 | *** | 10.6 | 101.3 ± 7.37 | 75.3 ± 5.99 | *** | 101.10 ± 5.71 | *** | 37.10 |
| SD | ** | ** | ns | 14.56 ± 0.06 | 14.4 ± 0.07 | ** | 15.77 ± 0.93 | ** | 8.83 | 17.51 ± 0.02 | 16.63 ± 0.18 | ** | 18.87 ± 0.93 | *** | 10.60 |
| Trait | Combined Analysis | Water-Stressed | Well-Watered | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gen | WR | Gen * WR | WUEL | WUIL | C1 | HY | C2 | HET | WUIL | C1 | HY | C2 | HET | |
| A | ** | *** | ns | 11.53 ± 2.31 | 7.31 ± 4.78 | *** | 4.45 ± 1.50 | *** | −47.43 | 30.21 ± 5.45 | ns | 31.90 ± 6.30 | ns | −0.78 |
| gs | ** | *** | ** | 0.04 ± 0.02 | 0.03 ± 0.02 | * | 0.02 ± 0.02 | * | −30.84 | 0.22 ± 0.08 | ns | 0.18 ± 0.02 | ** | −26.60 |
| E | *** | *** | ns | 0.83 ± 0.38 | 0.94 ± 0.81 | ns | 0.45 ± 0.15 | ** | −49.01 | 3.31 ± 0.82 | ns | 3.58 ± 0.70 | ns | −1.50 |
| VPDleaf/air | *** | *** | ns | 2.01 ± 0.72 | 2.11 ± 0.13 | * | 2.20 ± 0.13 | ** | 7.10 | 1.71 ± 0.19 | ns | 1.66 ± 0.20 | ns | 1.76 |
| TL | * | ** | ns | 30.47 ± 0.77 | 30.89 ± 0.24 | * | 31.06 ± 0.50 | ns | 1.25 | 30.25 ± 0.61 | ns | 29.90 ± 0.62 | ns | −0.61 |
| Fv/Fm | ns | *** | ** | 0.76 ± 0.02 | 0.78 ± 0.02 | * | 0.77 ± 0.01 | ns | −0.95 | 0.79 ± 0.01 | ns | 0.80 ± 0.006 | ns | 0.98 |
| SPAD index | * | ** | ns | 44.54 ± 1.23 | 45.48 ± 3.35 | ** | 44.14 ± 1.28 | ns | −1.86 | 50.93 ± 3.14 | ns | 49.59 ± 3.11 | ns | −2.84 |
| AWUE | *** | ** | * | 3.35 ± 0.02 | 3.08 ± 0.04 | * | 3.60 ± 1.28 | ** | 12.16 | 2.66 ± 0.55 | * | 3.80 ± 0.65 | *** | 32.7 |
| WF | *** | * | * | 0.3 ± 0.02 | 0.33 ± 0.06 | ns | 0.28 ± 0.02 | ** | −11.51 | 0.39 ± 0.08 | * | 0.27 ± 0.05 | *** | −24.7 |
| Trait | Combined Analysis | Water-Stressed | Well-Watered | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gen | WR | Gen * WR | WUEL | WUIL | C1 | HY | C2 | HET | WUEL | WUIL | C1 | HY | C2 | HET | |
| RL | * | ** | ns | 131.20 ± 18.44 | 124.2 ± 22.75 | * | 126.10 ± 20.01 | ns | −1.26 | 149.80 ± 19.34 | 132.20 ± 24.16 | * | 155.30 ± 12.77 | ns | 10.17 |
| RDW | *** | *** | ** | 6.86 ± 2.17 | 5.88 ± 1.19 | ** | 6.90 ± 1.06 | ns | 4.03 | 13.21 ± 2.15 | 8.84 ± 3.59 | ns | 15.53 ± 4.42 | *** | 20.12 |
| SRA | ** | ns | ns | 42.30 ± 9.99 | 34,12 ± 9.81 | ns | 32.80 ± 10.12 | ns | −13.39 | 31.50 ± 8.75 | 38.90 ± 8.50 | * | 33.10 ± 11.35 | ns | −5.85 |
| NSR | ns | ** | ns | 6.40 ± 1.51 | 6.22 ± 1.96 | ns | 5.60 ± 1.58 | ns | −11.06 | 7.50 ± 1.73 | 7.50 ± 2.04 | ns | 8.20 ± 1.22 | ns | 9.53 |
| SRD | ns | *** | ns | 4.23 ± 1.03 | 4.22 ± 1.45 | ns | 4.80 ± 1.99 | ns | 14.03 | 5.87 ± 1.11 | 5.73 ± 1.33 | ns | 6.33 ± 0.93 | ns | 9.19 |
| CRA | *** | ns | ** | 34.20 ± 3.32 | 35.75 ± 6.11 | ns | 26.70 ± 6.66 | ** | −23.21 | 31.60 ± 4.22 | 35.10 ± 5.94 | * | 27.60 ± 4.92 | ** | −2.56 |
| NCR | ns | * | ns | 17.40 ± 2.91 | 17.85 ± 3.89 | ns | 18.20 ± 4.61 | ns | 3.41 | 22.00 ± 3.13 | 19.20 ± 4.35 | ns | 19.90 ± 2.58 | ns | −3.23 |
| CRD | ns | ns | ns | 5.07 ± 1.00 | 4.55 ± 1.19 | ns | 4.93 ± 0.44 | ns | 3.50 | 5.27 ± 0.83 | 4.67 ± 0.81 | * | 5.03 ± 0.76 | ns | 2.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leite, J.T.; Amaral Junior, A.T.d.; Kamphorst, S.H.; Lima, V.J.d.; Santos Junior, D.R.d.; Schmitt, K.F.M.; Souza, Y.P.d.; Santos, T.d.O.; Bispo, R.B.; Mafra, G.S.; et al. Water Use Efficiency in Popcorn (Zea mays L. var. everta): Which Physiological Traits Would Be Useful for Breeding? Plants 2021, 10, 1450. https://doi.org/10.3390/plants10071450
Leite JT, Amaral Junior ATd, Kamphorst SH, Lima VJd, Santos Junior DRd, Schmitt KFM, Souza YPd, Santos TdO, Bispo RB, Mafra GS, et al. Water Use Efficiency in Popcorn (Zea mays L. var. everta): Which Physiological Traits Would Be Useful for Breeding? Plants. 2021; 10(7):1450. https://doi.org/10.3390/plants10071450
Chicago/Turabian StyleLeite, Jhean Torres, Antonio Teixeira do Amaral Junior, Samuel Henrique Kamphorst, Valter Jário de Lima, Divino Rosa dos Santos Junior, Kátia Fabiane Mereiros Schmitt, Yure Pequeno de Souza, Talles de Oliveira Santos, Rosimeire Barboza Bispo, Gabrielle Sousa Mafra, and et al. 2021. "Water Use Efficiency in Popcorn (Zea mays L. var. everta): Which Physiological Traits Would Be Useful for Breeding?" Plants 10, no. 7: 1450. https://doi.org/10.3390/plants10071450
APA StyleLeite, J. T., Amaral Junior, A. T. d., Kamphorst, S. H., Lima, V. J. d., Santos Junior, D. R. d., Schmitt, K. F. M., Souza, Y. P. d., Santos, T. d. O., Bispo, R. B., Mafra, G. S., Campostrini, E., & Rodrigues, W. P. (2021). Water Use Efficiency in Popcorn (Zea mays L. var. everta): Which Physiological Traits Would Be Useful for Breeding? Plants, 10(7), 1450. https://doi.org/10.3390/plants10071450