
Euphorbia characias: Phytochemistry and Biological Activities

 , , , and
, , , and
Abstract
:1. Introduction
2. Chemical Constituents
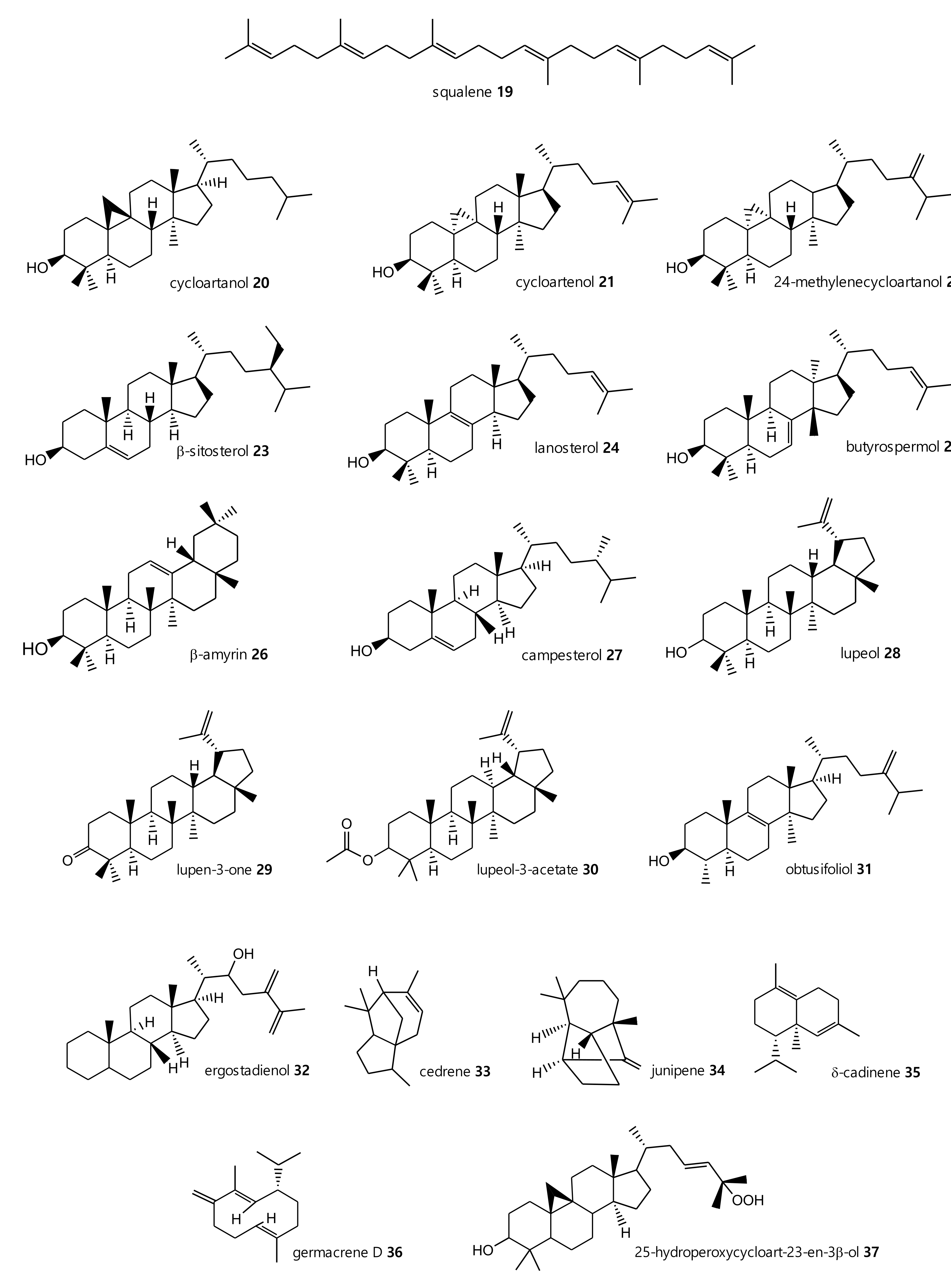
2.1. Terpenoids, Sterol Hydrocarbons, Fatty Acids and Cerebrosides
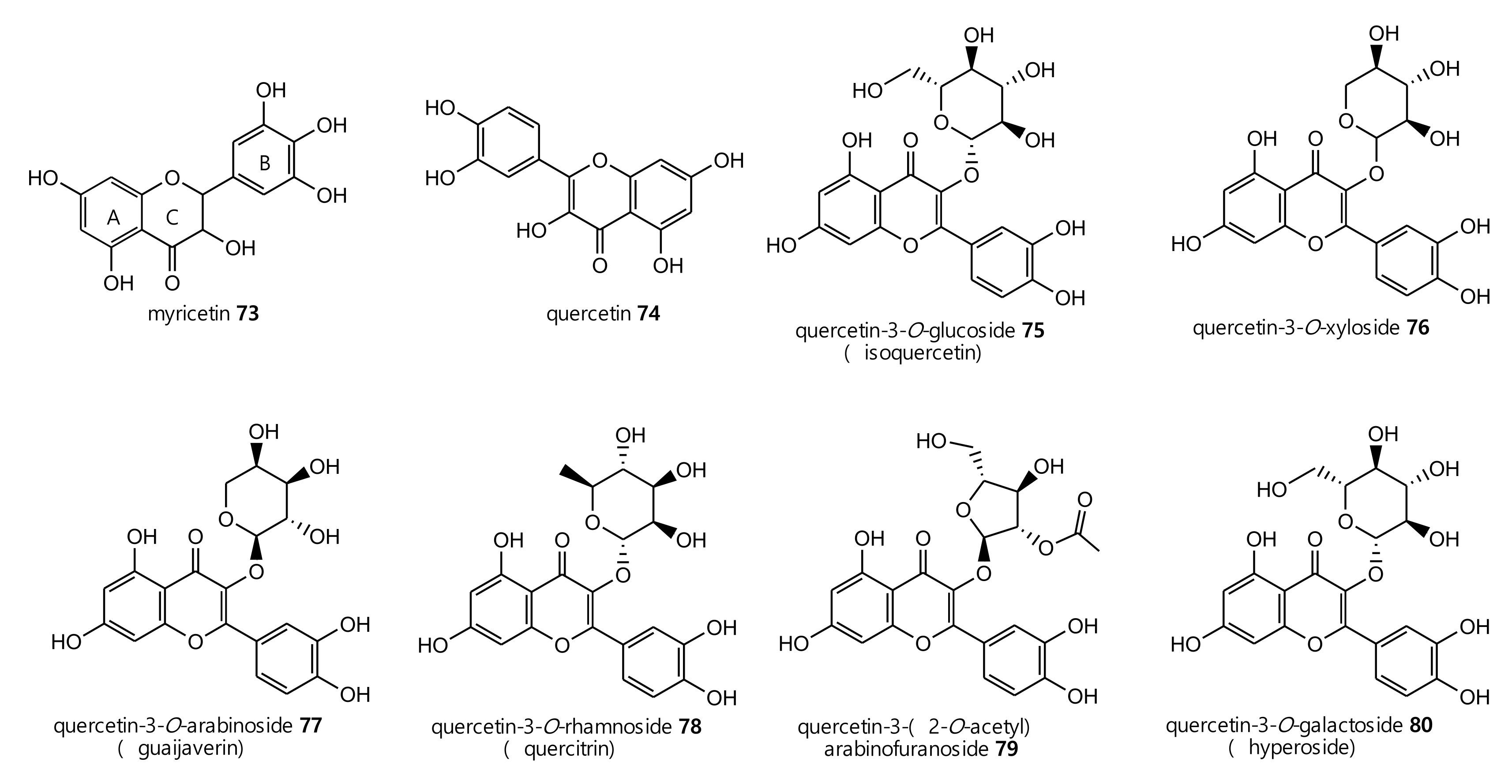
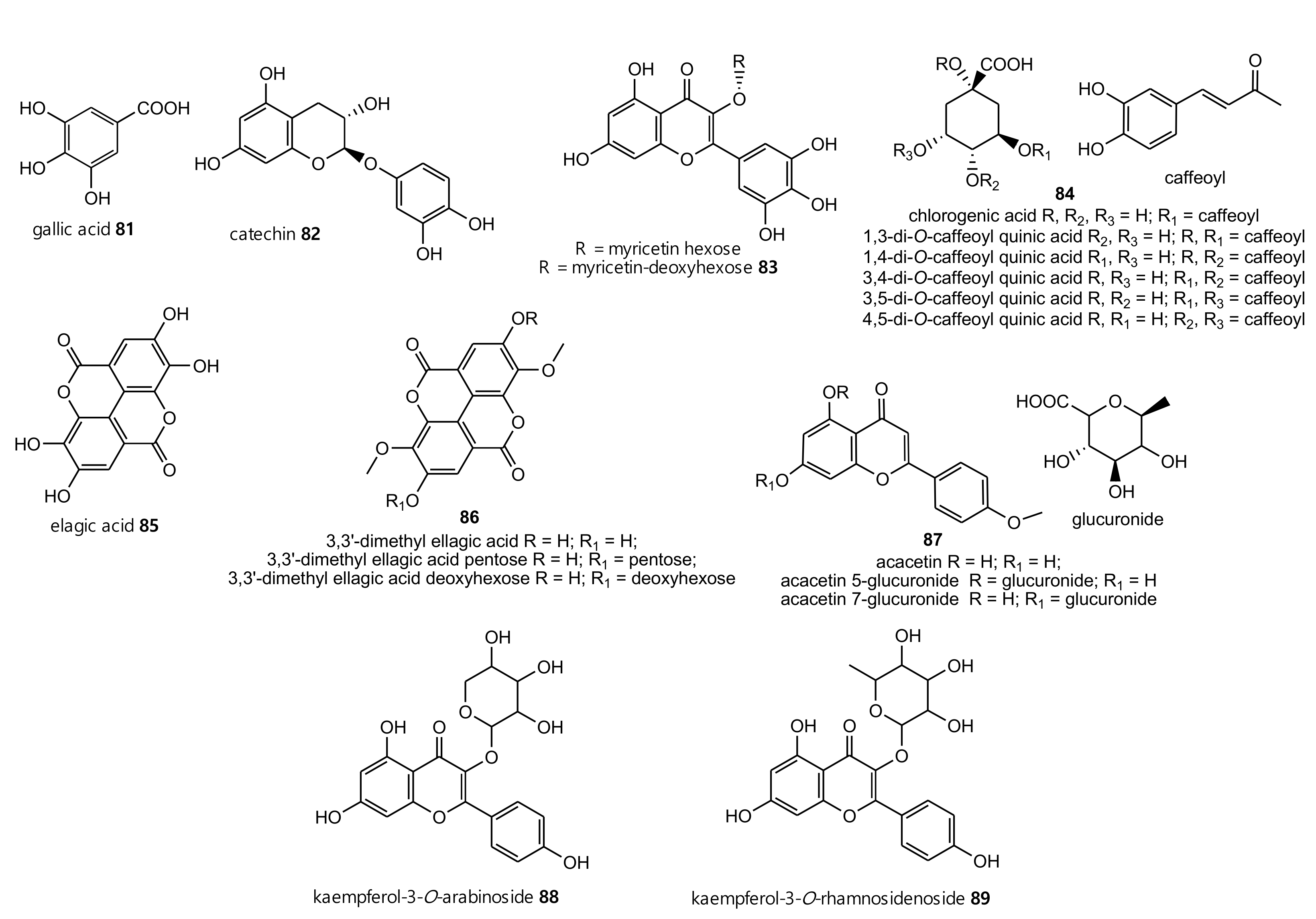
2.2. Phenolic and Carboxylic Acid Compounds
2.3. Natural Rubber and Rubber Particles
3. Biological Activities
3.1. Anti-Aging Properties
3.2. In Vivo Wound-Healing Activity
3.3. Cholinesterases Inhibition
3.4. Xanthine Oxidase Inhibition
3.5. Hypoglycemic Properties
3.6. Antioxidant Activity
3.7. Antiviral, Antimicrobial and Pesticidal Activities
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kemboi, D.; Peter, X.; Langat, M.; Tembu, J. A review of the ethnomedicinal uses, biological activities, and triterpenoids of euphorbia species. Molecules 2020, 25, 4019. [Google Scholar] [CrossRef]
- Mwine, J.T.; van Damme, P. Why do euphorbiaceae tick as medicinal plants? A review of euphorbiaceae family and its medicinal features. J. Med. Plants Res. 2011, 5, 652–662. [Google Scholar]
- Salehi, B.; Iriti, M.; Vitalini, S.; Antolak, H. Euphorbia-derived natural products with potential for use in health maintenance. Biomolecules 2019, 9, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pintus, F.; Medda, R.; Rinaldi, A.C.; Spanò, D.; Floris, G. Euphorbia latex biochemistry: Complex interactions in a complex environment. Plant Biosyst. 2010, 144, 381–391. [Google Scholar] [CrossRef]
- Pintus, F.; Spanò, D.; Medda, R.; Floris, G. Calcium ions and a secreted peroxidase in Euphorbia characias latex are made for each other. Protein J. 2011, 30, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Pintus, F.; Sabatucci, A.; Maccarrone, M.; Dainese, E.; Medda, R. Amine oxidase from Euphorbia characias: Kinetic and structural characterization. Biotechnol. Appl. Biochem. 2018, 65, 81–88. [Google Scholar] [CrossRef]
- Pintus, F.; Spano, D.; Corongiu, S.; Floris, G.; Medda, R. Purification, primary structure, and properties of Euphorbia characias latex purple acid phosphatase. Biochemistry 2011, 76, 694–701. [Google Scholar] [CrossRef]
- Sabatucci, A.; Pintus, F.; Cabras, T.; Vincenzoni, F.; Maccarrone, M.; Medda, R.; Dainese, E. Structure of a nucleotide pyrophosphatase/phosphodiesterase (NPP) from Euphorbia characias latex characterized by small-angle X-ray scattering: Clues for the general organization of plant NPPs. Acta Crystallogr. Sect. D Struct. Biol. 2020, 76, 857–867. [Google Scholar] [CrossRef]
- Spanò, D.; Pospiskova, K.; Safarik, I.; Pisano, M.B.; Pintus, F.; Floris, G.; Medda, R. Chitinase III in Euphorbia characias latex: Purification and characterization. Protein Expr. Purif. 2015, 116, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. Adv. Biochem. Eng. Biotechnol. 2015, 148, 63–106. [Google Scholar] [CrossRef]
- Shi, Q.W.; Su, X.H.; Kiyota, H. Chemical and pharmacological research of the plants in genus Euphorbia. Chem. Rev. 2008, 108, 4295–4327. [Google Scholar] [CrossRef]
- Appendino, G.; Belloro, E.; Cesare Tron, G.; Jakupovic, J.; Ballero, M. Polycyclic diterpenoids from Euphorbia characias. Fitoterapia 2000, 71, 134–142. [Google Scholar] [CrossRef]
- Vasas, A.; Hohmann, J. Euphorbia diterpenes: Isolation, Structure, biological activity, and synthesis (2008–2012). Chem. Rev. 2014, 114, 8579–8612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valente, C.; Pedro, M.; Duarte, A.; Maria, M.S.J.N.; Abreu, P.M.; Ferreira, M.J.U. Bioactive diterpenoids, a new jatrophane and two ent-abietanes, and other constituents from Euphorbia pubescens. J. Nat. Prod. 2004, 67, 902–904. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.C.; Hsieh, C.R.; Hsiao, G.; Chen, P.Y.; Chang, M.L.; Yin, H.W.; Lee, T.H.; Lee, C.K. Regulated expressions of MMP-2, -9 by diterpenoids from Euphorbia formosana hayata. Molecules 2012, 17, 2082–2090. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.E.; Parks, W.C. Metalloproteinases and their inhibitors: Regulators of wound healing. Int. J. Biochem. Cell Biol. 2008, 40, 1334–1347. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, K.R.; Munro, M.H.G.; Blunt, J.W.; Cardellina, J.H.; McMahon, J.B.; Gulakowski, R.J.; Cragg, G.M.; Cox, P.A.; Brinen, L.S.; Clardy, J.; et al. HIV inhibitory natural products. 3. Diterpenes from Homalantus acuminatus and Chrysobalanus icaco. Tetrahedron 1991, 47, 4547–4554. [Google Scholar] [CrossRef]
- Christodoulakis, N.S.; Mamoucha, S.; Termentzi, A.; Fokialakis, N. Leaf structure and histochemistry of the hardy evergreen Euphorbia characias L. (Mediterranean spurge). Flora Morphol. Distrib. Funct. Ecol. Plants 2015, 210, 13–18. [Google Scholar] [CrossRef]
- Seip, E.H.; Hecker, E. Lathyrane type diterpenoid esters from Euphorbia characias. Phytochemistry 1983, 22, 1791–1795. [Google Scholar] [CrossRef]
- Seip, E.H.; Hecker, E. Derivatives of characiol, macrocyclic diterpene esters of the jatrophane type from Euphorbia characias. Phytochemistry 1984, 23, 1689–1694. [Google Scholar] [CrossRef]
- Corea, G.; Fattorusso, E.; Lanzotti, V.; Motti, R.; Simon, P.N.; Dumontet, C.; Di Pietro, A. Structure-activity relationships for euphocharacins A-L, a new series of jatrophane diterpenes, as inhibitors of cancer cell P-glycoprotein. Planta Med. 2004, 70, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Barile, E.; Corea, G.; Lanzotti, V. Diterpenes from Euphorbia as potential leads for drug design. Nat. Prod. Commun. 2008, 3, 1003–1020. [Google Scholar] [CrossRef] [Green Version]
- Carriere, F.; Chagvardieff, P.; Gil, G.; Pean, M.; Sigoillot, J.C.; Tapie, P. Fatty acid patterns of neutral lipids from seeds, leaves and cell suspension cultures of Euphorbia characias. Phytochemistry 1992, 31, 2351–2353. [Google Scholar] [CrossRef]
- Bruni, R.; Muzzoli, M.; Ballero, M.; Loi, M.C.; Fantin, G.; Poli, F.; Sacchetti, G. Tocopherols, fatty acids and sterols in seeds of four Sardinian wild Euphorbia species. Fitoterapia 2004, 75, 50–61. [Google Scholar] [CrossRef]
- Escrig, P.V.; Iglesias, D.J.; Corma, A.; Primo, J.; Primo-Millo, E.; Cabedo, N. Euphorbia characias as bioenergy crop: A study of variations in energy value components according to phenology and water status. J. Agric. Food Chem. 2013, 61, 10096–10109. [Google Scholar] [CrossRef] [PubMed]
- Boieiro, M.; Espadaler, X.; Gómez, C.; Eustaquio, A. Spatial variation in the fatty acid composition of elaiosomes in an ant-dispersed plant: Differences within and between individuals and populations. Flora Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 497–502. [Google Scholar] [CrossRef]
- Palomino-Schätzlein, M.; Escrig, P.V.; Boira, H.; Primo, J.; Pineda-Lucena, A.; Cabedo, N. Evaluation of nonpolar metabolites in plant extracts by 13C NMR spectroscopy. J. Agric. Food Chem. 2011, 59, 11407–11416. [Google Scholar] [CrossRef]
- Falsone, G.; Cateni, F.; Katusian, F.; Wagner, H.; Seligmann, O.; Pellizer, G.; Asaro, F. New cerebrosides from Euphorbia characias. Z. Fur. Naturforsch. Sect. B J. Chem. Sci. 1993, 48, 1121–1126. [Google Scholar] [CrossRef] [Green Version]
- Falsone, G.; Cateni, F.; Baumgartner, M.; Lucchini, V.; Wagner, H.; Seligmann, O. Isolation and structure elucidation of five cerebrosides from Euphorbia characias L. Z. Fur. Naturforsch. Sect. B J. Chem. Sci. 1994, 49, 135–140. [Google Scholar] [CrossRef]
- Pintus, F.; Spanò, D.; Mascia, C.; Macone, A.; Floris, G.; Medda, R. Acetylcholinesterase inhibitory and antioxidant properties of Euphorbia characias latex. Rec. Nat. Prod. 2013, 7, 147–151. [Google Scholar]
- Pisano, M.B.; Cosentino, S.; Viale, S.; Spanò, D.; Corona, A.; Esposito, F.; Tramontano, E.; Montoro, P.; Tuberoso, C.I.G.; Medda, R.; et al. Biological activities of aerial parts extracts of Euphorbia characias. Biomed. Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Fais, A.; Era, B.; Di Petrillo, A.; Floris, S.; Piano, D.; Montoro, P.; Tuberoso, C.I.G.; Medda, R.; Pintus, F. Selected enzyme inhibitory effects of Euphorbia characias extracts. Biomed. Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Özbilgin, S.; Acıkara, Ö.B.; Akkol, E.K.; Süntar, I.; Keleş, H.; İşcan, G.S. In vivo wound-healing activity of Euphorbia characias subsp. wulfenii: Isolation and quantification of quercetin glycosides as bioactive compounds. J. Ethnopharmacol. 2018, 224, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Fokialakis, N.; Melliou, E.; Magiatis, P.; Harvala, C.; Mitaku, S. Composition of the steam volatiles of six Euphorbia spp. from Greece. Flavour Fragr. J. 2003, 18, 39–42. [Google Scholar] [CrossRef]
- Cornish, K.; Blakeslee, J. Rubber Biosynthesis in Plants. Available online: https://lipidlibrary.aocs.org/chemistry/physics/plant-lipid/rubber-biosynthesis-in-plants (accessed on 15 July 2021).
- Cherian, S.; Ryu, S.B.; Cornish, K. Natural rubber biosynthesis in plants, the rubber transferase complex, and metabolic engineering progress and prospects. Plant Biotechnol. J. 2019, 17, 2041–2061. [Google Scholar] [CrossRef] [Green Version]
- Spanò, D.; Pintus, F.; Mascia, C.; Scorciapino, M.A.; Casu, M.; Floris, G.; Medda, R. Extraction and characterization of a natural rubber from Euphorbia characias latex. Biopolymers 2012, 97, 589–594. [Google Scholar] [CrossRef]
- Mekkriengkrai, D.; Ute, K.; Swiezewska, E.; Chojnacki, T.; Tanaka, Y.; Sakdapipanich, J.T. Structural characterization of rubber from Jackfruit and Euphorbia as a model of natural rubber. Biomacromolecules 2004, 5, 2013–2019. [Google Scholar] [CrossRef] [PubMed]
- Spanò, D.; Pintus, F.; Esposito, F.; Loche, D.; Floris, G.; Medda, R. Euphorbia characias latex: Micromorphology of rubber particles and rubber transferase activity. Plant Physiol. Biochem. 2015, 87, 26–34. [Google Scholar] [CrossRef]
- Chen, Q.X.; Kubo, I. Kinetics of mushroom tyrosinase inhibition by quercetin. J. Agric. Food Chem. 2002, 50, 4108–4112. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.L.; Miyakawa, T.; Hayashi, Y.; Okamoto, K.; Hu, F.; Mitani, N.; Furihata, K.; Sawano, Y.; Tanokura, M. Isolation and tyrosinase inhibitory effects of polyphenols from the leaves of persimmon, Diospyros kaki. J. Agric. Food Chem. 2011, 59, 6011–6017. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Zhang, G.; Hu, X.; Xu, X.; Gong, D. Quercetin as a tyrosinase inhibitor: Inhibitory activity, conformational change and mechanism. Food Res. Int. 2017, 100, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Ademosun, A.O.; Oboh, G.; Bello, F.; Ayeni, P.O. Antioxidative properties and effect of quercetin and its glycosylated form (rutin) on acetylcholinesterase and butyrylcholinesterase activities. J. Evid. Based Complement. Altern. Med. 2016, 21, NP11–NP17. [Google Scholar] [CrossRef]
- Khan, M.T.H.; Orhan, I.; Şenol, F.S.; Kartal, M.; Şener, B.; Dvorská, M.; Šmejkal, K.; Šlapetová, T. Cholinesterase inhibitory activities of some flavonoid derivatives and chosen xanthone and their molecular docking studies. Chem. Biol. Interact. 2009, 181, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Park, M. Acetylcholinesterase inhibition by flavonoids from Agrimonia pilosa. Molecules 2007, 12, 2130–2139. [Google Scholar] [CrossRef] [Green Version]
- Orhan, I.; Kartal, M.; Tosun, F.; Şener, B. Screening of various phenolic acids and flavonoid derivatives for their anticholinesterase potential. Z. Fur. Naturforsch. Sect. C J. Biosci. 2007, 62, 829–832. [Google Scholar] [CrossRef] [PubMed]
- Katalinić, M.; Bosak, A.; Kovarik, Z. Flavonoids as inhibitors of human butyrylcholinesterase variants. Food Technol. Biotechnol. 2014, 52, 64–67. [Google Scholar]
- Webster, G.L. Irritant plants in the spurge family (Euphorbiaceae). Clin. Dermatol. 1986, 4, 36–45. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, R.; Zhang, G.; Gong, D. Mechanistic insights into the inhibition of quercetin on xanthine oxidase. Int. J. Biol. Macromol. 2018, 112, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Ademosun, A.O.; Ayeni, P.O.; Omojokun, O.S.; Bello, F. Comparative effect of quercetin and rutin on α-amylase, α-glucosidase, and some pro-oxidant-induced lipid peroxidation in rat pancreas. Comp. Clin. Path. 2015, 24, 1103–1110. [Google Scholar] [CrossRef]
- Yuan, E.; Liu, B.; Wei, Q.; Yang, J.; Chen, L.; Li, Q. Structure activity relationships of flavonoids as potent α-amylase inhibitors. Nat. Prod. Commun. 2014, 9, 1173–1176. [Google Scholar] [CrossRef] [Green Version]
- Utari, F.; Itam, A.; Syafrizayanti, S.; Putri, W.H.; Ninomiya, M.; Koketsu, M.; Tanaka, K.; Efdi, M. Isolation of flavonol rhamnosides from Pometia pinnata leaves and investigation of α-glucosidase inhibitory activity of flavonol derivatives. J. Appl. Pharm. Sci. 2019, 9, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Proença, C.; Freitas, M.; Ribeiro, D.; Oliveira, E.F.T.; Sousa, J.L.C.; Tomé, S.M.; Ramos, M.J.; Silva, A.M.S.; Fernandes, P.A.; Fernandes, E. α-Glucosidase inhibition by flavonoids: An in vitro and in silico structure–activity relationship study. J. Enzyme Inhib. Med. Chem. 2017, 32, 1216–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan-Nadilah, W.A.; Akhtar, M.T.; Shaari, K.; Khatib, A.; Hamid, A.A.; Hamid, M. Variation in the metabolites and α-glucosidase inhibitory activity of Cosmos caudatus at different growth stages. BMC Complement. Altern. Med. 2019, 19, 245. [Google Scholar] [CrossRef] [Green Version]
- Arung, E.T.; Furuta, S.; Ishikawa, H.; Kusuma, I.W.; Shimizu, K.; Kondo, R. Anti-melanogenesis properties of quercetin- and its derivative-rich extract from Allium cepa. Food Chem. 2011, 124, 1024–1028. [Google Scholar] [CrossRef]
- Bhullar, K.S.Z.; Warnakulasuriya, S.N.; Rupasinghe, H.P.V. Biocatalytic synthesis, structural elucidation, antioxidant capacity and tyrosinase inhibition activity of long chain fatty acid acylated derivatives of phloridzin and isoquercitrin. Bioorganic Med. Chem. 2013, 21, 684–692. [Google Scholar] [CrossRef]
- Mehmood, A.; Ishaq, M.; Zhao, L.; Safdar, B.; Rehman, A.; Munir, M.; Raza, A.; Nadeem, M.; Iqbal, W.; Wang, C. Natural compounds with xanthine oxidase inhibitory activity: A review. Chem. Biol. Drug Des. 2019, 93, 387–418. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.; Chen, J.; Liu, L.; Xiao, A. Screening and binding analysis of flavonoids with alpha-amylase inhibitory activity from lotus leaf. J. Braz. Chem. Soc. 2018, 29, 587–593. [Google Scholar] [CrossRef]
- Li, Y.Q.; Zhou, F.C.; Gao, F.; Bian, J.S.; Shan, F. Comparative evaluation of quercetin, isoquercetin and rutin as inhibitors of α-glucosidase. J. Agric. Food Chem. 2009, 57, 11463–11468. [Google Scholar] [CrossRef]
- Aoyama, H.; Sakagami, H.; Hatano, T. Three new flavonoids, proanthocyanidin, and accompanying phenolic constituents from Feijoa sellowiana. Biosci. Biotechnol. Biochem. 2018, 82, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Bu, Y.G.; Zhao, M.L.; Tao, R.; Luo, J.; Li, Y. Studies on antioxidant and α-glucosidase inhibitory constituents of Chinese toon bud (Toona sinensis). J. Funct. Foods 2020, 73. [Google Scholar] [CrossRef]
- Jeong, C.H.; Shim, K.H. Tyrosinase inhibitor isolated from the leaves of Zanthoxylum piperitum. Biosci. Biotechnol. Biochem. 2004, 68, 1984–1987. [Google Scholar] [CrossRef] [Green Version]
- Aderogba, M.A.; Ndhlala, A.R.; Rengasamy, K.R.R.; Van Staden, J. Antimicrobial and selected in vitro enzyme inhibitory effects of leaf extracts, flavonols and indole alkaloids isolated from Croton menyharthii. Molecules 2013, 18, 12633–12644. [Google Scholar] [CrossRef]
- Nithitanakool, S.; Pithayanukul, P.; Bavovada, R.; Saparpakorn, P. Molecular docking studies and anti-tyrosinase activity of thai mango seed kernel extract. Molecules 2009, 14, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.J. Antimelanogenic and antioxidant properties of gallic acid. Biol. Pharm. Bull. 2007, 30, 1052–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhary, D.K.; Mishra, A. In vitro and in silico interaction of faba bean (Vicia faba L.) seed extract with xanthine oxidase and evaluation of antioxidant activity as a therapeutic potential. Nat. Prod. Res. 2019, 33, 2689–2693. [Google Scholar] [CrossRef] [PubMed]
- Kam, A.; Li, K.M.; Razmovski-Naumovski, V.; Nammi, S.; Shi, J.; Chan, K.; Li, G.Q. A comparative study on the inhibitory effects of different parts and chemical constituents of pomegranate on α-amylase and α-glucosidase. Phyther. Res. 2013, 27, 1614–1620. [Google Scholar] [CrossRef]
- Tang, H.; Cui, F.; Li, H.; Huang, Q.; Li, Y. Understanding the inhibitory mechanism of tea polyphenols against tyrosinase using fluorescence spectroscopy, cyclic voltammetry, oximetry, and molecular simulations. RSC Adv. 2018, 8, 8310–8318. [Google Scholar] [CrossRef] [Green Version]
- Suganthy, N.; Pandima Devi, K. In vitro antioxidant and anti-cholinesterase activities of Rhizophora mucronata. Pharm. Biol. 2016, 54, 118–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagao, A.; Seki, M.; Kobayashi, H. Inhibition of xanthine oxidase of flavonoids. Biosci. Biotechnol. Biochem. 1999, 63, 1787–1790. [Google Scholar] [CrossRef]
- Júnior, J.P.L.; de Franco, R.R.; Saraiva, A.L.; Moraes, I.B.; Espindola, F.S. Anacardium humile St. Hil as a novel source of antioxidant, antiglycation and α-amylase inhibitors molecules with potential for management of oxidative stress and diabetes. J. Ethnopharmacol. 2021, 268. [Google Scholar] [CrossRef]
- Zhao, L.; Wen, L.; Lu, Q.; Liu, R. Interaction mechanism between α-glucosidase and A-type trimer procyanidin revealed by integrated spectroscopic analysis techniques. Int. J. Biol. Macromol. 2020, 143, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Hwang, S.H.; Huang, B.; Lim, S.S. Identification of tyrosinase specific inhibitors from Xanthium strumarium fruit extract using ultrafiltration-high performance liquid chromatography. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2015, 1002, 319–328. [Google Scholar] [CrossRef]
- Strzępek-Gomółka, M.; Gaweł-Bęben, K.; Angelis, A.; Antosiewicz, B.; Sakipova, Z.; Kozhanova, K.; Głowniak, K.; Kukula-Koch, W. Identification of mushroom and murine tyrosinase inhibitors from achillea biebersteinii Afan. Extract. Molecules 2021, 26, 964. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.T.T.; Awale, S.; Tezuka, Y.; Le Tran, Q.; Watanabe, H.; Kadota, S. Xanthine oxidase inhibitory activity of vietnamese medicinal plants. Biol. Pharm. Bull. 2004, 27, 1414–1421. [Google Scholar] [CrossRef] [Green Version]
- Olennikov, D.N.; Kashchenko, N.I. Componential profile and amylase inhibiting activity of phenolic compounds from Calendula officinalis L. leaves. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Ooi, K.L.; Muhammad, T.S.T.; Tan, M.L.; Sulaiman, S.F. Cytotoxic, apoptotic and anti-α-glucosidase activities of 3,4-di-O-caffeoyl quinic acid, an antioxidant isolated from the polyphenolic-rich extract of Elephantopus mollis Kunth. J. Ethnopharmacol. 2011, 135, 685–695. [Google Scholar] [CrossRef]
- Park, S.; Jegal, J.; Chung, K.W.; Jung, H.J.; Noh, S.G.; Chung, H.Y.; Ahn, J.; Kim, J.; Yang, M.H. Isolation of tyrosinase and melanogenesis inhibitory flavonoids from Juniperus chinensis fruits. Biosci. Biotechnol. Biochem. 2018, 82, 2041–2048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pintus, F.; Spanò, D.; Corona, A.; Medda, R. Antityrosinase activity of Euphorbia characias extracts. PeerJ 2015, 2015. [Google Scholar] [CrossRef]
- Tsatsop Tsague, R.K.T.; Kenmogne, S.B.; Tchienou, G.E.D.; Parra, K.; Ngassoum, M.B. Sequential extraction of quercetin-3-O-rhamnoside from Piliostigma thonningii Schum. leaves using microwave technology. SN Appl. Sci. 2020, 2, 1230. [Google Scholar] [CrossRef]
- Arima, H.; Danno, G.I. Isolation of antimicrobial compounds from guava (psidium guajava L.) and their structural elucidation. Biosci. Biotechnol. Biochem. 2002, 66, 1727–1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jabir, N.R.; Khan, F.R.; Tabrez, S. Cholinesterase targeting by polyphenols: A therapeutic approach for the treatment of alzheimer’s disease. CNS Neurosci. Ther. 2018, 24, 753–762. [Google Scholar] [CrossRef]
- Szwajgier, D.; Borowiec, K. Phenolic acids from malt are efficient acetylcholinesterase and butyrylcholinesterase inhibitors. J. Inst. Brew. 2012, 118, 40–48. [Google Scholar] [CrossRef]
- Dizdar, M.; Vidic, D.; Požgan, F.; Štefane, B.; Maksimović, M. Acetylcholinesterase inhibition and antioxidant activity of N-trans-caffeoyldopamine and N-trans-feruloyldopamine. Sci. Pharm. 2018, 86, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oboh, G.; Ogunsuyi, O.B.; Ogunbadejo, M.D.; Adefegha, S.A. Influence of gallic acid on α-amylase and α-glucosidase inhibitory properties of acarbose. J. Food Drug Anal. 2016, 24, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Mathew, S.; Abraham, T.E.; Zakaria, Z.A. Reactivity of phenolic compounds towards free radicals under in vitro conditions. J. Food Sci. Technol. 2015, 52, 5790–5798. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Kim, S.; Kim, S.; Song, Y.; Seung, K.; Hong, D.; Khang, G.; Lee, D. Antioxidant and anti-inflammatory activities of hydroxybenzyl alcohol releasing biodegradable polyoxalate nanoparticles. Biomacromolecules 2010, 11, 2103–2108. [Google Scholar] [CrossRef] [PubMed]
- Groussard, C.; Morel, I.; Chevanne, M.; Monnier, M.; Cillard, J.; Delamarche, A. Free radical scavenging and antioxidant effects of lactate ion: An in vitro study. J. Appl. Physiol. 2000, 89, 169–175. [Google Scholar] [CrossRef]
- Zheng, Y.Z.; Deng, G.; Liang, Q.; Chen, D.F.; Guo, R.; Lai, R.C. Antioxidant activity of quercetin and its glucosides from propolis: A theoretical study. Sci. Rep. 2017, 7, 7543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacopini, P.; Baldi, M.; Storchi, P.; Sebastiani, L. Catechin, epicatechin, quercetin, rutin and resveratrol in red grape: Content, in vitro antioxidant activity and interactions. J. Food Compos. Anal. 2008, 21, 589–598. [Google Scholar] [CrossRef]
- Razavi, S.M.; Zahri, S.; Zarrini, G.; Nazemiyeh, H.; Mohammadi, S. Biological activity of quercetin-3-O-glucoside, a known plant flavonoid. Russ. J. Bioorganic Chem. 2009, 35, 376–378. [Google Scholar] [CrossRef]
- Zhao, M.H.; Jiang, Z.T.; Liu, T.; Li, R. Flavonoids in Juglans regia L. leaves and evaluation of in vitro antioxidant activity via intracellular and chemical methods. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Materska, M.; Perucka, I. Antioxidant activity of the main phenolic compounds isolated from hot pepper fruit (Capsicum annuum L.). J. Agric. Food Chem. 2005, 53, 1750–1756. [Google Scholar] [CrossRef]
- Liu, Z.; Tao, X.; Zhang, C.; Lu, Y.; Wei, D. Protective effects of hyperoside (quercetin-3-o-galactoside) to PC12 cells against cytotoxicity induced by hydrogen peroxide and tert-butyl hydroperoxide. Biomed. Pharmacother. 2005, 59, 481–490. [Google Scholar] [CrossRef]
- Hung, T.M.; Na, M.K.; Thuong, P.T.; Su, N.D.; Sok, D.E.; Song, K.S.; Seong, Y.H.; Bae, K.H. Antioxidant activity of caffeoyl quinic acid derivatives from the roots of Dipsacus asper Wall. J. Ethnopharmacol. 2006, 108, 188–192. [Google Scholar] [CrossRef]
- Jun, Y.L.; Eun, J.C.; Hyo, J.K.; Jun, H.P.; Sang, W.C. Antioxidative flavonoids from leaves of Carthamus tinctorius. Arch. Pharm. Res. 2002, 25, 313–319. [Google Scholar] [CrossRef]
- Kilic, I.; Yeşiloǧlu, Y.; Bayrak, Y. Spectroscopic studies on the antioxidant activity of ellagic acid. Spectrochim. Acta Part. A Mol. Biomol. Spectrosc. 2014, 130, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.Y.; Sun, Y.X.; Shahidi, F. Preparation and antioxidant activity of tyrosol and hydroxytyrosol esters. J. Funct. Foods 2017, 37, 66–73. [Google Scholar] [CrossRef]
- Babaei, H.; Sadeghpour, O.; Nahar, L.; Delazar, A.; Nazemiyeh, H.; Mansouri, M.R.; Poursaeid, N.; Asnaashari, S.; Moghadam, S.B.; Sarker, S.D. Antioxidant and vasorelaxant activities of flavonoids from Amygdalus lycioides var. horrida. Turkish J. Biol. 2008, 32, 203–208. [Google Scholar]
- Nazemiyeh, H.; Kazemi, E.M.; Zare, K.; Jodari, M.; Nahar, L.; Sarker, S.D. Free radical scavengers from the aerial parts of Euphorbia petiolata. J. Nat. Med. 2010, 64, 187–190. [Google Scholar] [CrossRef]
- Kumar, S.; Prahalathan, P.; Raja, B. Antihypertensive and antioxidant potential of vanillic acid, a phenolic compound in L-NAME-induced hypertensive rats: A dose-dependence study. Redox Rep. 2011, 16, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Chethan, S.; Dharmesh, S.M.; Malleshi, N.G. Inhibition of aldose reductase from cataracted eye lenses by finger millet (Eleusine coracana) polyphenols. Bioorganic Med. Chem. 2008, 16, 10085–10090. [Google Scholar] [CrossRef] [PubMed]
- Kiliç, I.; Yeşiloǧlu, Y. Spectroscopic studies on the antioxidant activity of p-coumaric acid. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2013, 115, 719–724. [Google Scholar] [CrossRef]
- Kikuzaki, H.; Hisamoto, M.; Hirose, K.; Akiyama, K.; Taniguchi, H. Antioxidant properties of ferulic acid and its related compounds. J. Agric. Food Chem. 2002, 50, 2161–2168. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, I. Antioxidant activity of caffeic acid (3,4-dihydroxycinnamic acid). Toxicology 2006, 217, 213–220. [Google Scholar] [CrossRef]
- Gordon, M.H.; Roedig-Penman, A. Antioxidant activity of quercetin and myricetin in liposomes. Chem. Phys. Lipids 1998, 97, 79–85. [Google Scholar] [CrossRef]
- Pekkarinen, S.S.; Heinonen, I.M.; Hopia, A.I. Flavonoids quercetin, myricetin, kaemferol and (+)-catechin as antioxidants in methyl linoleate. J. Sci. Food Agric. 1999, 79, 499–506. [Google Scholar] [CrossRef]
- Áy, É.; Hunyadi, A.; Mezei, M.; Minárovits, J.; Hohmann, J. Flavonol 7-O-glucoside herbacitrin inhibits HIV-1 replication through simultaneous integrase and reverse transcriptase inhibition. Evid. Based Complement. Altern. Med. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, N.; Pizza, C.; Aquino, R.D.; Tommasi, N.; Piacente, S.; Colman, S.B.A.S.; Hay, A.J. Inhibition of HIV infection by flavanoids. Antiviral Res. 1993, 22, 189–199. [Google Scholar] [CrossRef]
- Kratz, J.M.; Andrighetti-Fröhner, C.R.; Kolling, D.J.; Leal, P.C.; Cirne-Santos, C.C.; Yunes, R.A.; Nunes, R.J.; Trybala, E.; Bergström, T.; Frugulhetti, I.C.P.P.; et al. Anti-HSV-1 and anti-HIV-1 activity of gallic acid and pentyl gallate. Mem. Inst. Oswaldo Cruz 2008, 103, 437–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijaya, S.J.; Yanuar, A.; Handayani, R.; Syahdi, R.R. In silico analysis of flavonoid glycosides and its aglycones as reverse transcriptase inhibitor. Pharmacogn. J. 2019, 11, 1252–1255. [Google Scholar] [CrossRef]
- Akroum, S.; Bendjeddou, D.; Satta, D.; Lalaoui, K. Antibacterial activity and acute toxicity effect of flavonoids extracted from mentha longifolia. Am. J. Sustain. Agric. 2009, 4, 93–96. [Google Scholar]
- Rattanachaikunsopon, P.; Phumkhachorn, P. Contents and antibacterial activity of flavonoids extracted from leaves of Psidium guajava. J. Med. Plants Res. 2010, 4, 393–396. [Google Scholar]
- Gutiérrez-Larraínzar, M.; Rúa, J.; Caro, I.; de Castro, C.; de Arriaga, D.; García-Armesto, M.R.; del Valle, P. Evaluation of antimicrobial and antioxidant activities of natural phenolic compounds against foodborne pathogens and spoilage bacteria. Food Control. 2012, 26, 555–563. [Google Scholar] [CrossRef]
- Rawdkuen, S. Antimicrobial activity of some potential active compounds against food spoilage microorganisms. African J. Biotechnol. 2012, 11, 13914–13921. [Google Scholar] [CrossRef] [Green Version]
- Guzman, J.D. Natural cinnamic acids, synthetic derivatives and hybrids with antimicrobial activity. Molecules 2014, 19, 19292–19349. [Google Scholar] [CrossRef]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Giordani, R.; Trebaux, J.; Masi, M.; Regli, P. Enhanced antifungal activity of ketoconazole by Euphorbia characias latex against Candida albicans. J. Ethnopharmacol. 2001, 78, 1–5. [Google Scholar] [CrossRef]
- Martos, S.; Spanò, D.; Agustí, N.; Poschenrieder, C.; Pintus, F.; Moles, L.; Medda, R. A chitinase from Euphorbia characias latex is a novel and powerful plant-based pesticide against Drosophila suzukii. Ann. Appl. Biol. 2017, 171, 252–263. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Structure | Compound |
|---|---|
| ent-abietanes-1 | |
 | 1 8α,14-dihydro-7-oxojolkinolide E R = H |
| 2 caudicifolin R = OH (8α,14-dihydro-7-oxohelioscopinolide A) | |
| ent-abietanes-2 | |
 | 3 jolkinolide E R1 = R2 = H (ent-abieta-8(14),13(15)-dien-16,12-olide) |
| 4 helioscopinolide A R1 = OH; R2 = H | |
| 5 helioscopinolide B R1 = H; R2 = OH | |
 | 6 helioscopinolide E |
| ent-atisanes | |
 | 7 ent-2-hydroxyatisa-1,16(17)-diene-3,14-dione |
 | R R1 8 ent-atis-16(17)-ene-3,14-dione H C=O 9 ent-3α-hydroxyatis-16(17)-ene-2,14-dione C=O OH |
| ent-kauranes | |
 | 10 16β,17-dihydroxy-ent-kauran-3-one |
| ent-pimaranes | |
 | 11 kaurene |
 | 12 ent-pimara-8(14),15-dien-3α-ol |
 | R R1 13 3β,15,16-triacetoxy-ent-pimar-8(14)-ene H OAc 14 3β,15,16-triacetoxy-ent-pimar-8(14)-en-2-one C=O OAc |
| cembranes | |
 | 15 cembrene |
| Scheme | Compound |
|---|---|
| lathyranes | |
 | 16a 15-O-acetyl-3-O-propionyljolkiol-5β,6β-oxide) R1 = COCH2Me; R2 = COMe; R3 = Me; R4 = H; 16b 15-O-acetyl-3-O-iso-butyryljolkiol-5β,6β-oxide) R1 = COCHMe2; R2 = COMe; R3 = Me; R4 = H; 16c 15-O-acetyl-3-O-tigloyljolkiol-5β,6β-oxide) R1 = COC(Me) ECHMe; R2 = COMe; R3 = Me; R4 = H; 16d 15-O-acetyl-3-O-benzoyljolkiol-5β,6β-oxide) R1 = COC6H5; R2 = COMe; R3 = Me; R4 = H; 16e 15-O-acetyl-3-O-nicotinoyljolkiol-5β,6β-oxide) R1 = COC5H4N; R2 = COMe; R3 = Me; R4 = H; 16f 2α-O-acetyl-3-O-iso-butyryl-15-O-nicotinoyljolkinol-5β,6β-oxide R1 = COCHMe2; R2 = COC5H4N; R3 = Me; R4 = OCOMe |
| jatrophanes | |
 | 17a 15-O-acetyl-3-O-tigloylcharaciol-5β,6β-oxide) R1 = tigloyl (COC(Me) ECHMe); R2 = COCH3; 17b 15-O-acetyl-3-O-benzoylcharaciol-5β,6β-oxide) R1 = benzoyl (COC6H5); R2 = COCH3 |
 | 17c 15-O-acetyl-3-O-propionylcharaciol R1 = propionyl (COCH2Me); R2 = COCH3 |
 | 17d 15-O-acetyl-5β-hydroxyisocharaciol-3-iso-butyrate) R1 = iso-butyryl-(COCHMe2); R2 = COCH3; 17e 15-O-acetyl-5β-hydroxyisocharaciol-3-tigliate R1 = tigloyl (COC(Me) ECHMe); R2 = COCH3; 17f 15-O-acetyl-5β-hydroxyisocharaciol-3-benzoate R1 = benzoyl (COC6H5); R2 = COCH3 |
 | 17g 2,5,8,15-O-triacetyl, nicotinoyl-2,5β,8-trihydroxyisocharaciol-3-benzoate R = acetyl (x 3), benzoyl, nicotinoyl; 17h 2,5,8,15-O-triacetyl, nicotinoyl-2,5β,8-trihydroxyisocharaciol-3-tigliate R = acetyl (x 3), tigloyl, nicotinoyl; |
 | 18a-l euphocharacin 18a R1 = OH; R2 = Bz; R3 = Ac; R4 = Nic; R5 = Ac 18b R1 = OH; R2 = Bz; R3 = Ac; R4 = Nic; R5 = H 18c R1 = OH; R2 = Bz; R3 = Ac; R4 = Bz; R5 = H 18d R1 = OH; R2 = MeBu; R3 = Ac; R4 = Nic; R5 = Ac 18e R1 = H; R2 = Bz; R3 = Ac; R4 = Nic; R5 = H 18f R1 = H; R2 = Bz; R3 = Ac; R4 = Nic; R5 = Ac 18g R1 = H; R2 = iBu; R3 = Ac; R4 = Nic; R5 = H 18h R1 = H; R2 = iBu; R3 = Ac; R4 = Nic; R5 = Ac 18i R1 = H; R2 = Pr; R3 = Ac; R4 = Nic; R5 = Ac 18j R1 = H; R2 = Ac; R3 = Ac; R4 = Nic; R5 = Ac 18k R1 = H; R2 = iBu; R3 = H; R4 = Nic; R5 = Ac 18l R1 = OH; R2 = Bz; R3 = H; R4 = Nic; R5 = Ac Pr = propionyl; iBu = isobutiryl; MeBu = 2-methylbutiryl; Bz = benzoyl; Nic = nicotinoyl |
| cerebrosides 47–50 [28] |
 |
| 47 (2S, 3S, 4R, 5R, 6Z)-l-O-(β-d-glucopyranosyl)-2-N-[2′R)-2′-hydroxy-(15′Z)-tetracosenoylamino]-6(Z)-octadecene-l,3,4,5-tetraol |
| 48 (2S, 3S, 4R, 5R, 6Z)-l-O-(β-d-glucopyranosyl)-2-N-[(2′R)-2′-hydroxy-(17′Z)-hexacosenoy1amino]-6(Z)-octadecene-l,3,4,5-tetraol |
| 49 (2S, 3S, 4R, 5R, 6Z)-l-O-(β-d-glucopyranosyl)-2-N-[(2′R)-2′-hydroxy-(19′Z)-octacosenoylamino]-6(Z)-octadecene-l,3,4,5-tetraol |
| 50 (2S, 3S, 4R, 5R, 6Z)-l-O-(β-d-glucopyranosyl)-2-N-[(2′R)-2′-hydroxyoctacosanoylamino]-(Z)-octadecene-l,3,4,5-tetraol |
| cerebrosides 51–55 [29] |
 |
| 51 (2S, 3S, 4R, 8Z)-l-O-(β-d-glucopyranosyl)-2-N-[(2′R)-2′-hydroxy-(15′Z)-tetracosenoylamino]-8(Z)-octadecene-1,3,4-triol |
| 52 (2S, 3S, 4R, 8Z)-l-O-(β-d-glucopyranosyl)-2-N-[(2′R)-2′-hydroxy-(17′Z)-hexacosenoyl]-8(Z)-octadecene-1,3,4-triol |
| 53 (2S, 3S, 4R, 8Z)-l-O-(β-d-glucopyranosyl)-2-N-[(2′R)-2′-hydroxy-(19′Z)-octacosenoylamino]-8(Z)-octadecene-l,3,4-triol |
| 54 (2S, 3S, 4R, 8Z)-l-O-(β-d-glucopyranosyl)-2-N-[(2′R)-2′-hydroxyhexacosanoylamino]-8(Z)-octadecene-1,3,4-triol |
| 55 (2S, 3S, 4R, 8Z)-l-O-(β-d-glucopyranosyl)-2-N-[(2′R)-2′-hydroxyheptacosanoylamino]-8(Z)-octadecene-l,3,4-triol |
| Compound | Part of the Plant | Reference |
|---|---|---|
| heptacosane 90 | Stems | [25] |
| nonacosane 91 | Leaves, Stems | [25] |
| pentacosene 92 | ||
| hentriacontane 93 | ||
| triacontane 94 | ||
| tritriacontane 95 | ||
| heneicosane 96 | Flowers | [34] |
| docosane 97 | ||
| tricosane 98 | ||
| pentacosane 99 | ||
| heptacosane 100 | ||
| nonacosane 101 | ||
| phytol 102 | Leaves, Flowers | [25,34] |
| tetracosanol 103 | Stems | [25] |
| hexacosanol 104 | Leaves, Stems | [25] |
| octacosanol 105 | ||
| inositol 106 | ||
| myo-inositol 107 | ||
| octacosanal 108 | ||
| nonanal 109 | Flowers | [34] |
| tridecanal 110 | ||
| tetradecanal 111 | ||
| hexadecanal 112 | ||
| hexacosanal 113 | ||
| 6,10,14-trimethyl-2-pentadecanone 114 | ||
| farnesyl acetone 115 | ||
| α-tocopherol 116 | Leaves | [25] |
| hexadecanoic acid 117 | Flowers | [34] |
| pyroglutamic acid 118 | Leaves, Stems | [25] |
| D-glucose 119 | ||
| D-fructose 120 | ||
| sucrose 121 |
| Part of the Plant/Reference Compounds | Extract | IC50 (µg/mL) | |||||
|---|---|---|---|---|---|---|---|
| Tyrosinase | AChE | BChE | XO | α-Amylase | α-Glucosidase | ||
| Leaves | Water | 120 ± 10 [mixed-type] | (4.2 ± 0.25) × 103 | NI | >200 | 74.02 ± 3.06 | 1.4 ± 0.11 |
| Ethanol | 34 ± 2 [competitive] | 600 ± 56 | 390 ± 40 | 68.9 ± 6.6 [mixed-type] | 25.41 ± 1.42 [noncompetitive] | 0.8 ± 0.03 [noncompetitive] | |
| Stems | Water | (1.80 ± 0.13) × 103 | (6.9 ± 0.71) × 103 | NI | - | - | - |
| Ethanol | (1.10 ± 0.090) × 103 | (5.8 ± 0.43) × 103 | NI | - | - | - | |
| Flowers | Water | 490 ± 25 | (5.25 ± 0.35) × 103 | (4.20 ± 0.39) × 103 | >200 | 109.12 ± 10.36 | 1.1 ± 0.07 |
| Ethanol | 150 ± 11 | 600 ± 56 | (1.22 ± 0.08) × 103 | 85.5 ± 6.4 [mixed-type] | 29.39 ± 1.41 [noncompetitive] | 0.9 ± 0.04 [uncompetitive] | |
| Kojic acid | 0.8 ± 0.03 | - | - | - | - | - | |
| Galantamine | - | 0.27 ± 0.07 | 8.12 ± 0.61 | - | - | - | |
| Allopurinol | - | - | - | 0.012 | - | - | |
| Acarbose | - | - | - | - | 8.04 ± 0.65 | 90 ± 7.3 | |
| Compound | Tyrosinase | AChE | BChE | XO | α-Amylase | α-Glucosidase |
|---|---|---|---|---|---|---|
| quercetin 74 | [40,41,42] | [43,44,45,46] | [43,44,46,47] | [48,49] | [50,51] | [50,52,53,54] |
| quercetin-3-O-glucoside 75 | [41,55,56] | - | - | [57] | [58] | [59] |
| quercetin-3-O-xyloside 76 | [60] | - | - | - | - | - |
| quercetin-3-O-arabinoside 77 | [60] | - | - | - | - | [61] |
| quercetin-3-O-rhamnoside 78 | [62] | [63] | - | - | - | [52] |
| gallic acid 81 | [64,65] | [46] | [46] | [66] | NI [67] | [67] |
| catechin 82 | [68] | [69] | [69] | [70] | [71] | [72] |
| di-O-caffeoylquinic acid 84 | [73,74] | - | - | [75] | [76] | [77] |
| kaempferol-3-O-arabinoside 88 | - | - | - | - | - | [61] |
| kaempferol-3-O-rhamnoside 89 | [78] | [63] | - | - | - | [61] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fais, A.; Delogu, G.L.; Floris, S.; Era, B.; Medda, R.; Pintus, F. Euphorbia characias: Phytochemistry and Biological Activities. Plants 2021, 10, 1468. https://doi.org/10.3390/plants10071468
Fais A, Delogu GL, Floris S, Era B, Medda R, Pintus F. Euphorbia characias: Phytochemistry and Biological Activities. Plants. 2021; 10(7):1468. https://doi.org/10.3390/plants10071468
Chicago/Turabian StyleFais, Antonella, Giovanna Lucia Delogu, Sonia Floris, Benedetta Era, Rosaria Medda, and Francesca Pintus. 2021. "Euphorbia characias: Phytochemistry and Biological Activities" Plants 10, no. 7: 1468. https://doi.org/10.3390/plants10071468
APA StyleFais, A., Delogu, G. L., Floris, S., Era, B., Medda, R., & Pintus, F. (2021). Euphorbia characias: Phytochemistry and Biological Activities. Plants, 10(7), 1468. https://doi.org/10.3390/plants10071468