
The Cell Division Cycle of Euglena gracilis Indicates That the Level of Circadian Plasticity to the External Light Regime Changes in Prolonged-Stationary Cultures


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
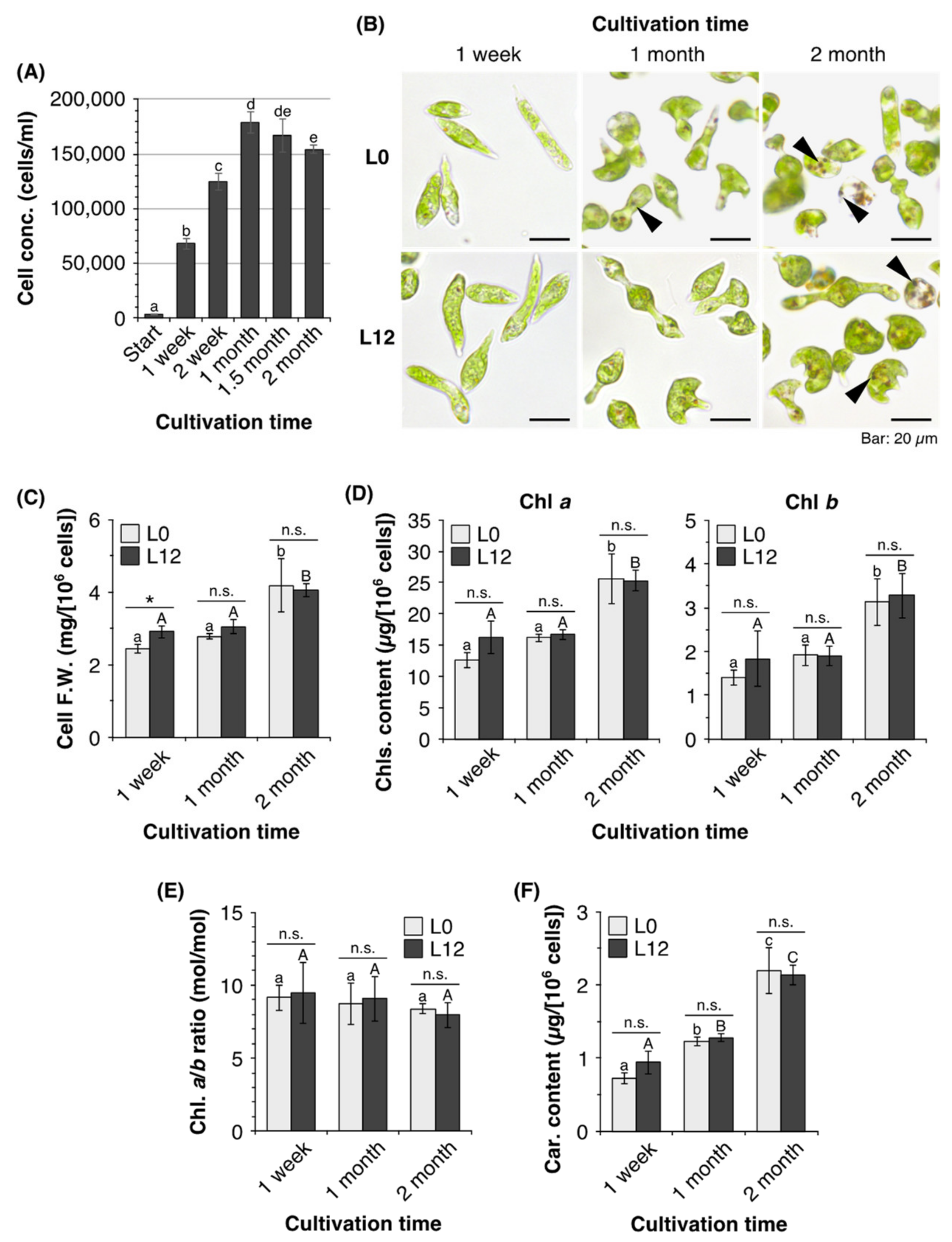
2.1. Characteristics of E. gracilis Cells in Old Cultures
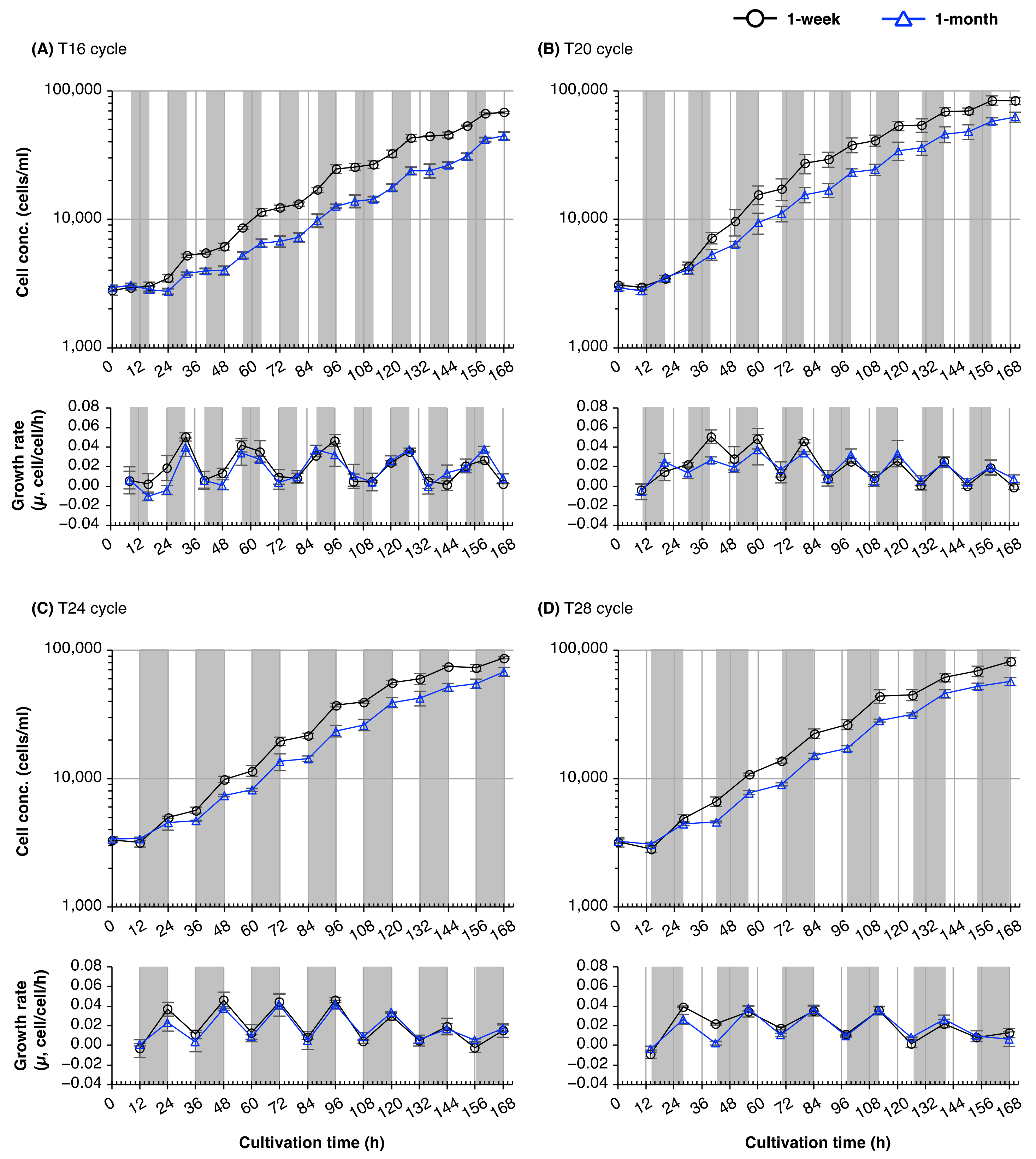
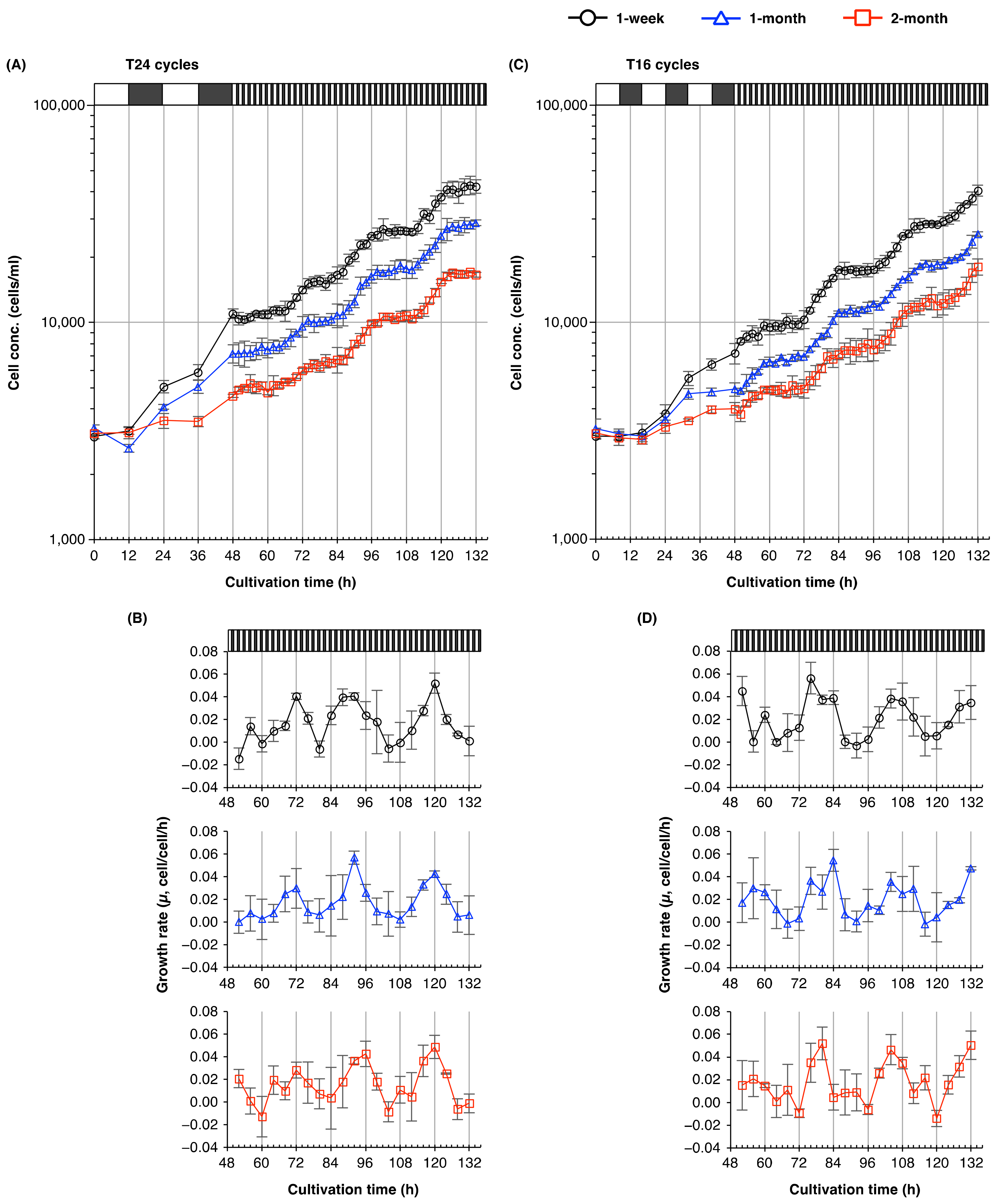
2.2. The Cell Division Cycle of Euglena Shows Circadian Resonance with the Light/Dark Cycle
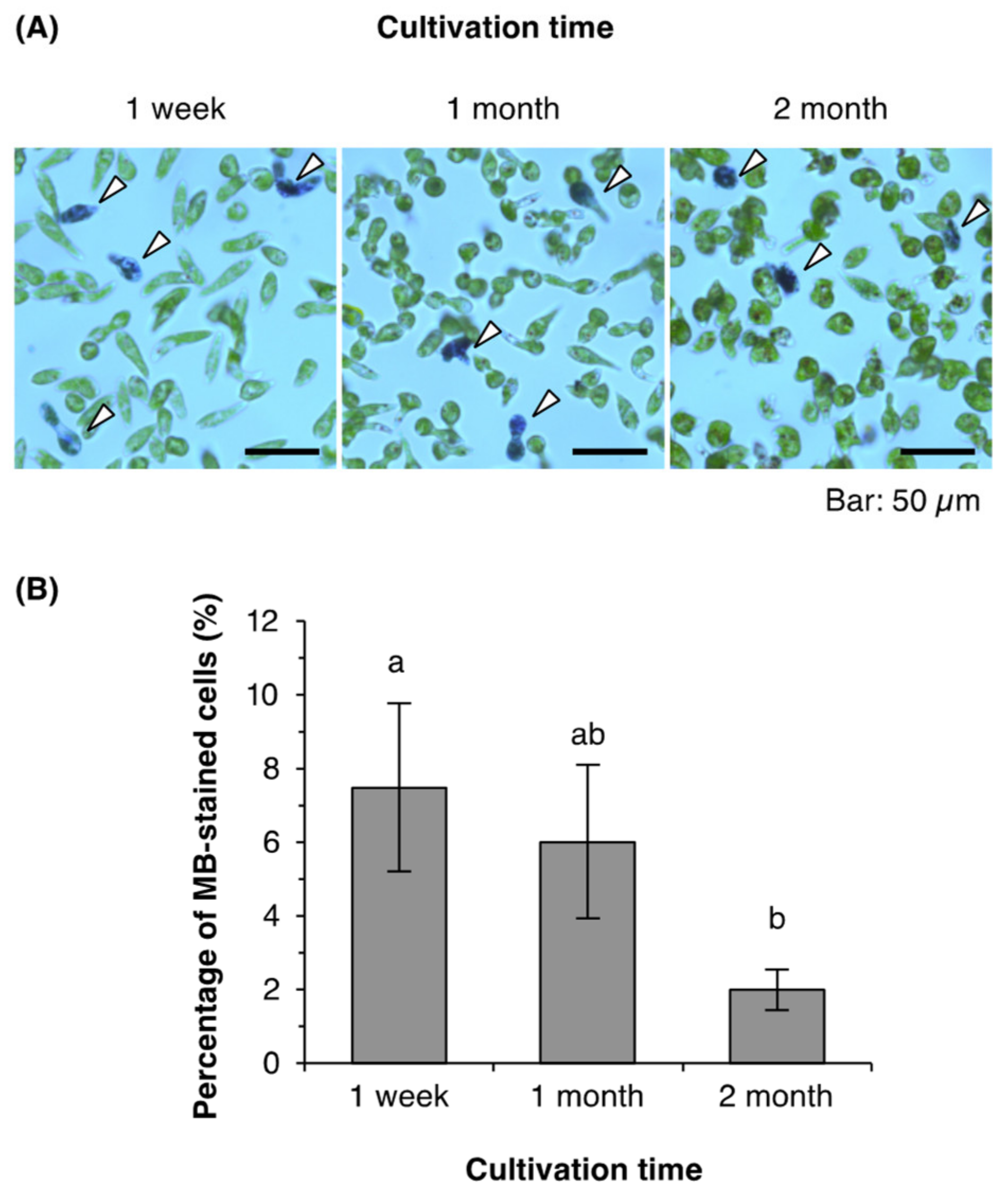
2.3. Cell Viability of Euglena from Young and Old Cultures
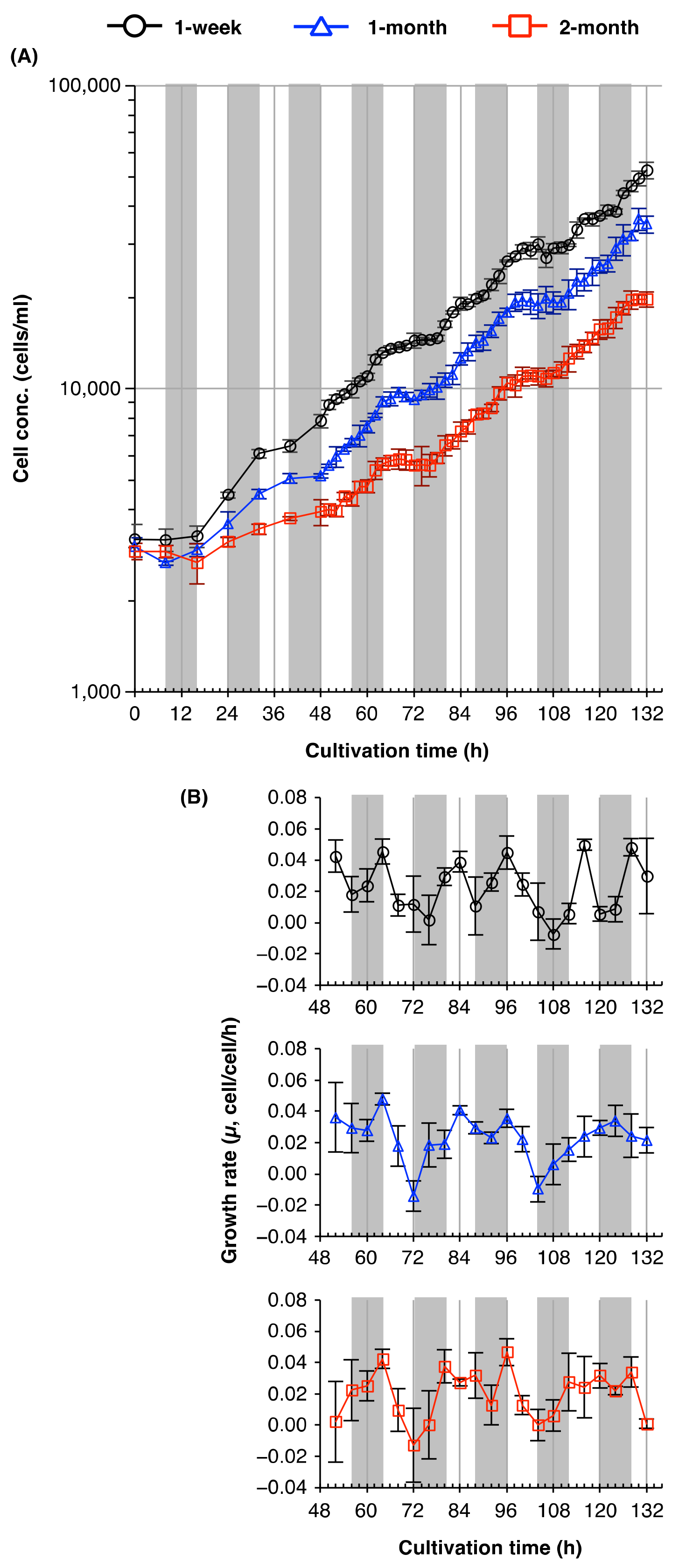
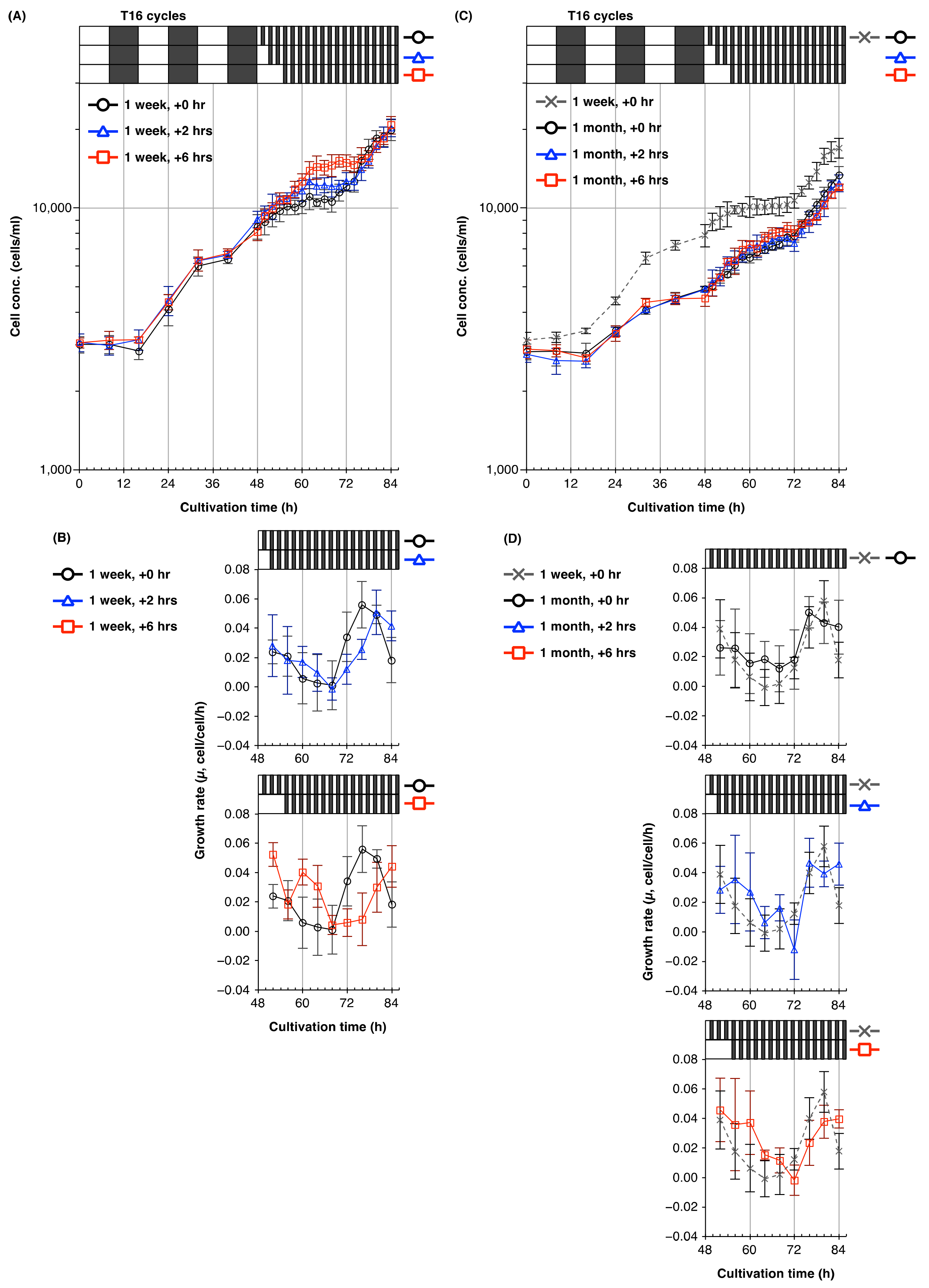
2.4. Plasticity of the Circadian Rhythm of Cell Division in Young and Old Cultures of Euglena
3. Discussion
3.1. Characteristics of E. gracilis Cells in Old Cultures
3.2. Circadian Resonance of Cell Division of Euglena with Light/Dark Cycles
3.3. Viability of Euglena Cells from Young and Old Cultures
3.4. Plasticity of Circadian Rhythms of Cell Division in Euglena from Young and Old Cultures
4. Materials and Methods
4.1. Biological Materials
4.2. Time-Course Analysis of Euglena Cell Division in Young and Old Cultures
4.3. Determination of Photosynthetic Pigment Content
4.4. Dye Exclusion Assay
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inoue, K.; Araki, T.; Endo, M. Circadian clock during plant development. J. Plant Res. 2018, 131, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Greenwood, M.; Locke, J.C.W. The circadian clock coordinates plant development through specificity at the tissue and cellular level. Curr. Opin. Plant Biol. 2020, 53, 65–72. [Google Scholar] [CrossRef]
- Noordally, Z.B.; Millar, A.J. Clocks in algae. Biochemistry 2015, 54, 171–183. [Google Scholar] [CrossRef]
- Mori, T.; Binder, B.; Johnson, C.H. Circadian gating of cell division in cyanobacteria growing with average doubling times of less than 24 hours. Proc. Natl. Acad. Sci. USA 1996, 93, 10183–10188. [Google Scholar] [CrossRef] [Green Version]
- Hagiwara, S.; Bolige, A.; Zhang, Y.; Takahashi, M.; Yamagishi, A.; Goto, K. Circadian gating of photoinduction of commitment to cell-cycle transitions in relation to photoperiodic control of cell reproduction in Euglena. Photochem. Photobiol. 2002, 76, 105–115. [Google Scholar] [CrossRef]
- Goto, K.; Johnson, C.H. Is the cell division cycle gated by a circadian clock? The case of Chlamydomonas reinhardtii. J. Cell Biol. 1995, 129, 1061–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyagishima, S.; Fujiwara, T.; Sumiya, N.; Hirooka, S.; Nakano, A.; Kabeya, Y.; Nakamura, M. Translation-independent circadian control of the cell cycle in a unicellular photosynthetic eukaryote. Nat. Commun. 2014, 5, 3807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, H.; Nakamichi, N.; Hisatsune, M.; Murase, H.; Mizuno, T. Synchronization of plant circadian oscillators with a phase delay effect of the vein network. Phys. Rev. Lett. 2007, 99, 098102. [Google Scholar] [CrossRef]
- Endo, M.; Shimizu, H.; Nohales, M.A.; Araki, T.; Kay, S.A. Tissue-specific clocks in Arabidopsis show asymmetric coupling. Nature 2014, 515, 419–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, H.; Araki, T.; Endo, M. Photoperiod sensitivity of the Arabidopsis circadian clock is tissue-specific. Plant Signal. Behav. 2015, 10, e1010933. [Google Scholar] [CrossRef] [Green Version]
- Bordage, S.; Sullivan, S.; Laird, J.; Millar, A.J.; Nimmo, H.G. Organ specificity in the plant circadian system is explained by different light inputs to the shoot and root clocks. New Phytol. 2016, 212, 136–149. [Google Scholar] [CrossRef]
- Greenwood, M.; Domijan, M.; Gould, P.D.; Hall, A.J.W.; Locke, J.C.W. Coordinated circadian timing through the integration of local inputs in Arabidopsis thaliana. PLoS Biol. 2019, 17, e3000407. [Google Scholar] [CrossRef] [Green Version]
- Lim, P.O.; Nam, H.G. Aging and senescence of the leaf organ. J. Plant Biol. 2007, 50, 291–300. [Google Scholar] [CrossRef]
- Lim, P.O.; Kim, H.J.; Nam, H.G. Leaf senescence. Annu. Rev. Plant Biol. 2007, 58, 115–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, P.O.; Woo, H.R.; Nam, H.G. Molecular genetics of leaf senescence in Arabidopsis. Trends Plant Sci. 2003, 8, 272–278. [Google Scholar] [CrossRef]
- Kim, H.; Kim, Y.; Yeom, M.; Lim, J.; Nam, H.G. Age-associated circadian period changes in Arabidopsis leaves. J. Exp. Bot. 2016, 67, 2665–2673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, M.; Mitesser, O.; Hovestadt, T.; Yoshii, T.; Rieger, D.; Helfrich-Förster, C. The circadian clock improves fitness in the fruit fly, Drosophila melanogaster. Front. Physiol. 2019, 10, 1374. [Google Scholar] [CrossRef]
- Ouyang, Y.; Andersson, C.R.; Kondo, T.; Golden, S.S.; Johnson, C.H. Resonating circadian clocks enhance fitness in cyanobacteria. Proc. Natl. Acad. Sci. USA 1998, 95, 8660–8664. [Google Scholar] [CrossRef] [Green Version]
- Dodd, A.N.; Salathia, N.; Hall, A.; Kévei, E.; Tóth, R.; Nagy, F.; Hibberd, J.M.; Millar, A.J.; Webb, A.A.R. Plant circadian clocks increase photosynthesis, growth, survival, and competitive advantage. Science 2005, 309, 630–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adl, S.M.; Simpson, A.G.B.; Farmer, M.A.; Andersen, R.A.; Anderson, O.R.; Barta, J.R.; Bowser, S.S.; Brugerolle, G.; Fensome, R.A.; Fredericq, S.; et al. The new higher level classification of Eukaryotes with emphasis on the taxonomy of protists. J. Eukaryot. Microbiol. 2005, 52, 399–451. [Google Scholar] [CrossRef]
- Schwartzbach, S.D.; Shigeoka, S. Euglena: Biochemistry, Cell and Molecular Biology; Schwartzbach, S.D., Shigeoka, S., Eds.; Springer: Cham, Switzerland, 2017; Volume 979, ISBN 9783319549088. [Google Scholar]
- Edmunds, L.N., Jr.; Tay, D.E.; Laval-Martin, D.L. Cell division cycles and circadian clocks: Phase-response curves for light perturbations in synchronous cultures of Euglena. Plant Physiol. 1982, 70, 297–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edmunds, L.N., Jr.; Funch, R. Effects of “skeleton” photoperiods and high frequency light-dark cycles on the rhythm of cell division in synchronized cultures of Euglena. Planta 1969, 87, 134–163. [Google Scholar] [CrossRef] [PubMed]
- Goto, K.; Laval-Martin, D.L.; Edmunds, L.N., Jr. Biochemical modeling of an autonomously oscillatory circadian clock in Euglena. Science 1985, 228, 1284–1288. [Google Scholar] [CrossRef]
- Gomez, M.P.; Harris, J.B.; Walne, P.L. Studies of Euglena gracilis in aging cultures. I. Light microscopy and cytochemistry. Brit. Phycol. J. 1974, 9, 163–174. [Google Scholar] [CrossRef]
- Kashiyama, Y.; Yokoyama, A.; Shiratori, T.; Hess, S.; Not, F.; Bachy, C.; Gutierrez-Rodriguez, A.; Kawahara, J.; Suzaki, T.; Nakazawa, M.; et al. Taming chlorophylls by early eukaryotes underpinned algal interactions and the diversification of the eukaryotes on the oxygenated earth. ISME J. 2019, 13, 1899–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, S.; Takaichi, S.; Ishikawa, T.; Asahina, M.; Takahashi, S.; Shinomura, T. Identification and functional analysis of the geranylgeranyl pyrophosphate synthase gene (crtE) and phytoene synthase gene (crtB) for carotenoid biosynthesis in Euglena gracilis. BMC Plant Biol. 2016, 16, 4. [Google Scholar] [CrossRef] [Green Version]
- Kato, S.; Tanno, Y.; Takaichi, S.; Shinomura, T. Low temperature stress alters the expression of phytoene desaturase genes (crtP1 and crtP2) and the ζ-carotene desaturase gene (crtQ) together with the cellular carotenoid content of Euglena gracilis. Plant Cell Phys. 2019, 60, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Soshino, M.; Takaichi, S.; Ishikawa, T.; Nagata, N.; Asahina, M.; Shinomura, T. Suppression of the phytoene synthase gene (EgcrtB) alters carotenoid content and intracellular structure of Euglena gracilis. BMC Plant Biol. 2017, 17, 125. [Google Scholar] [CrossRef]
- Gomez, M.P.; Harris, J.B.; Walne, P.L. Studies of Euglena gracilis in aging cultures II. Ultrastructure. Brit. Phycol. J. 1974, 9, 175–193. [Google Scholar] [CrossRef] [Green Version]
- Hofman, M.A.; Swaab, D.F. Living by the clock: The circadian pacemaker in older people. Ageing Res. Rev. 2006, 5, 33–51. [Google Scholar] [CrossRef]
- Rakshit, K.; Krishnan, N.; Guzik, E.M.; Pyza, E.; Giebultowicz, J.M. Effects of aging on the molecular circadian oscillations in Drosophila. Chronobiol. Int. 2012, 29, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Dharmananda, S.; Feldman, J.F. Spatial distribution of circadian clock phase in aging cultures of Neurospora crassa. Plant Physiol. 1979, 63, 1049–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pittendrigh, C.S.; Daan, S. Circadian oscillations in rodents: A systematic increase of their frequency with age. Science 1974, 186, 548–550. [Google Scholar] [CrossRef] [Green Version]
- Hagiwara, S.; Takahashi, M.; Yamagishi, A.; Zhang, Y.; Goto, K. Novel findings regarding photoinduced commitments of G1-, S- and G2-phase cells to cell-cycle transitions in darkness and dark-induced G1-, S- and G2-phase arrests in Euglena. Photochem. Photobiol. 2001, 74, 726–733. [Google Scholar] [CrossRef]
- Yamazaki, S.; Straume, M.; Tei, H.; Sakaki, Y.; Menaker, M.; Block, G.D. Effects of aging on central and peripheral mammalian clocks. Proc. Natl. Acad. Sci. USA 2002, 99, 10801–10806. [Google Scholar] [CrossRef] [Green Version]
- Dodd, A.N.; Dalchau, N.; Gardner, M.J.; Baek, S.-J.; Webb, A.A.R. The circadian clock has transient plasticity of period and is required for timing of nocturnal processes in Arabidopsis. New Phytol. 2014, 201, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Aton, S.J.; Block, G.D.; Tei, H.; Yamazaki, S.; Herzog, E.D. Plasticity of circadian behavior and the suprachiasmatic nucleus following exposure to non-24-hour light cycles. J. Biol. Rhythm. 2004, 19, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Pittendrigh, C.S.; Daan, S. A functional analysis of circadian pacemakers in nocturnal rodents. J. Comp. Physiol. 1976, 106, 223–252. [Google Scholar] [CrossRef]
- Barrett, R.K.; Page, T.L. Effects of light on circadian pacemaker development. J. Comp. Physiol. A 1989, 165, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Cramer, M.; Myers, J. Growth and photosynthetic characteristics of Euglena gracilis. Arc. Microbiol. 1952, 17, 384–402. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta Bba Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kato, S.; Nam, H.G. The Cell Division Cycle of Euglena gracilis Indicates That the Level of Circadian Plasticity to the External Light Regime Changes in Prolonged-Stationary Cultures. Plants 2021, 10, 1475. https://doi.org/10.3390/plants10071475
Kato S, Nam HG. The Cell Division Cycle of Euglena gracilis Indicates That the Level of Circadian Plasticity to the External Light Regime Changes in Prolonged-Stationary Cultures. Plants. 2021; 10(7):1475. https://doi.org/10.3390/plants10071475
Chicago/Turabian StyleKato, Shota, and Hong Gil Nam. 2021. "The Cell Division Cycle of Euglena gracilis Indicates That the Level of Circadian Plasticity to the External Light Regime Changes in Prolonged-Stationary Cultures" Plants 10, no. 7: 1475. https://doi.org/10.3390/plants10071475
APA StyleKato, S., & Nam, H. G. (2021). The Cell Division Cycle of Euglena gracilis Indicates That the Level of Circadian Plasticity to the External Light Regime Changes in Prolonged-Stationary Cultures. Plants, 10(7), 1475. https://doi.org/10.3390/plants10071475