
Characterization of Chlamydomonas reinhardtii Mutants That Exhibit Strong Positive Phototaxis

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Mutant Screening for Phototactic Signs
2.2. Morphology and Motility of ap Mutants Are Normal
2.3. Photosynthesis Phenotypes of ap Mutants
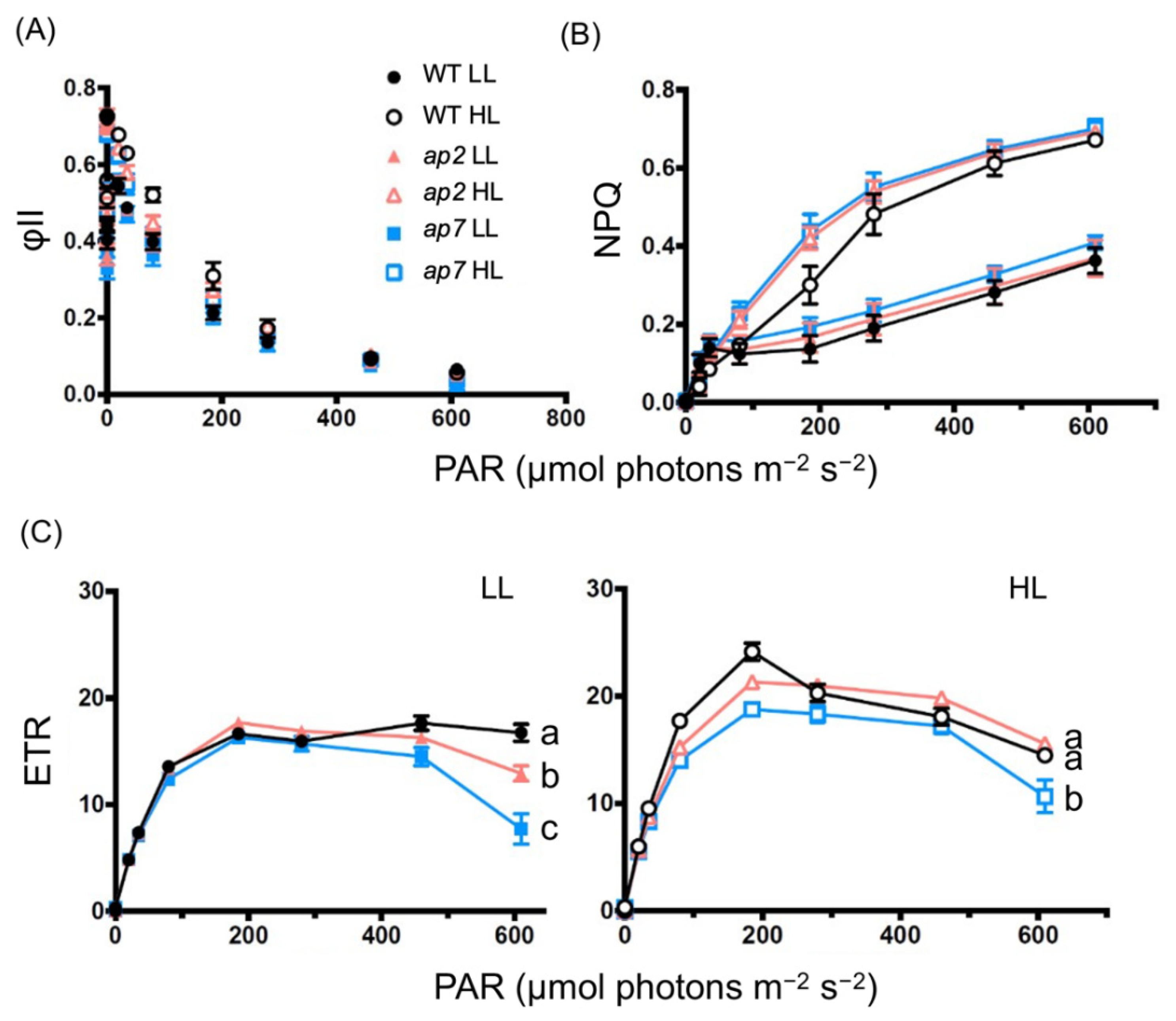
2.4. Detailed Photosynthesis Phenotype Analyses of ap2 and ap7
3. Discussion
3.1. Phenotypic Discrepancy among ap Mutants
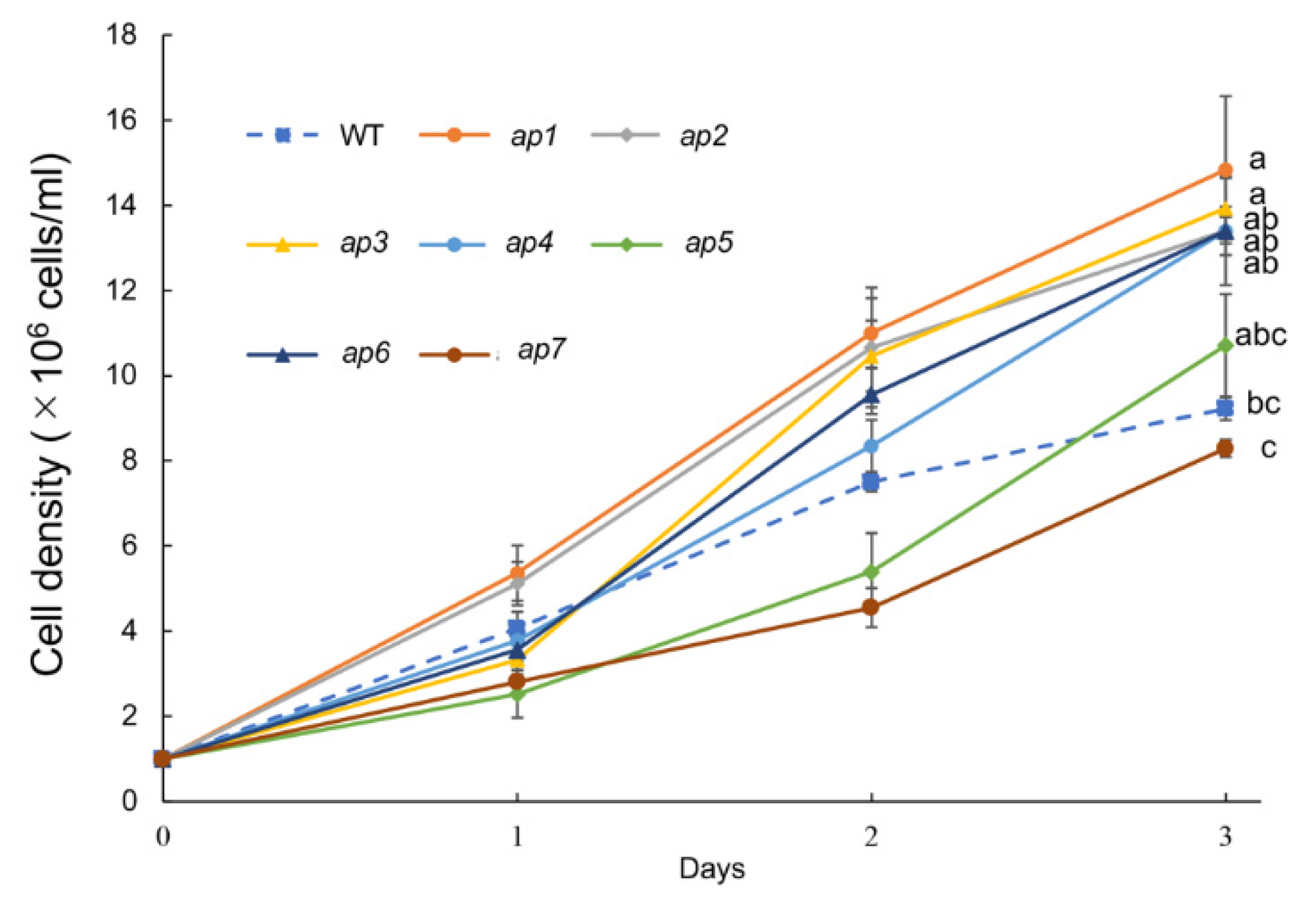
3.2. Growth Phenotype
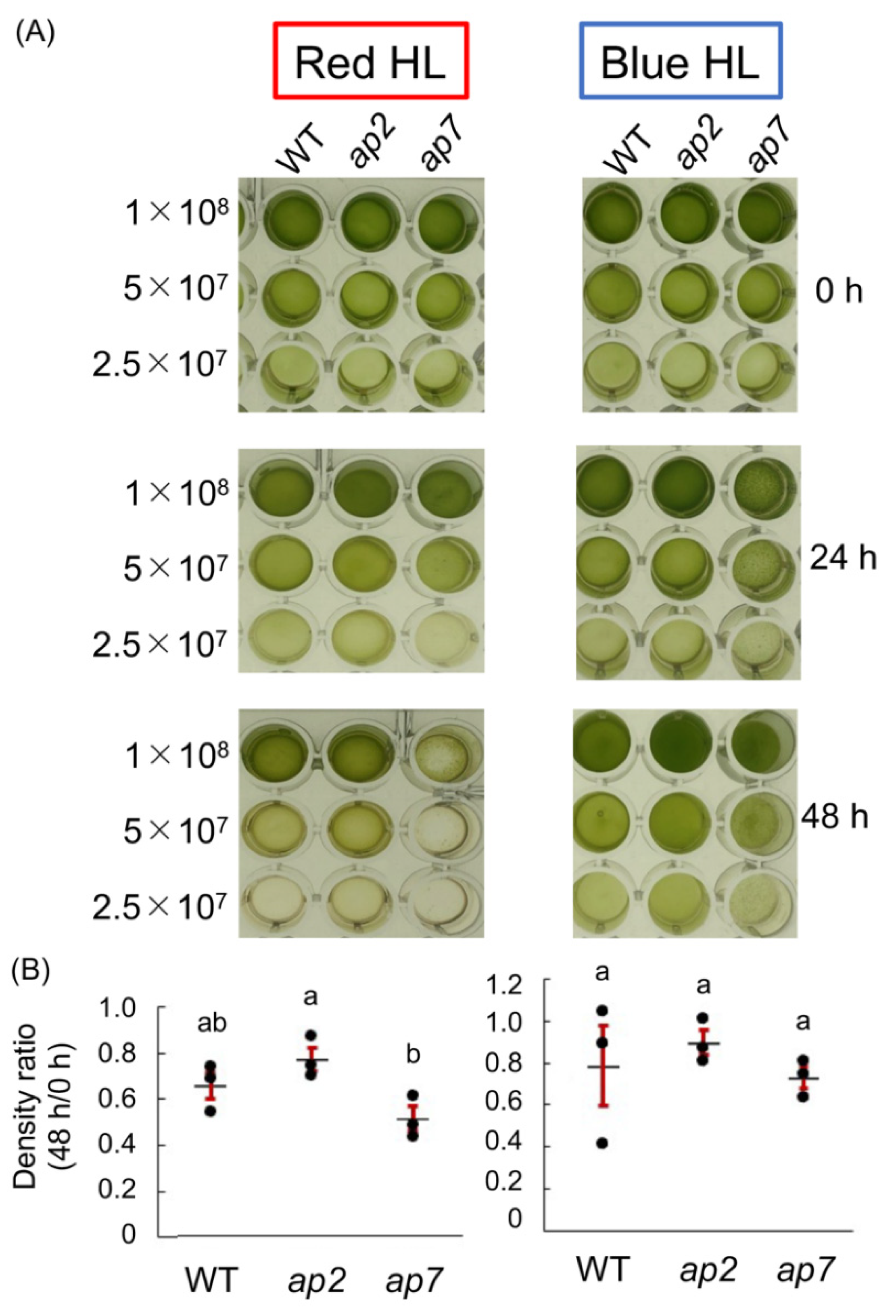
3.3. High-Light Tolerance
4. Conclusions
5. Materials and Methods
5.1. Cell Culture and Strains
5.2. Mutagenesis and Screening for ap Mutants
5.3. Cell-Level Phototaxis Assay
5.4. Ciliary Beating Frequency Measurement
5.5. Cell Density and Cell Size Measurement
5.6. Growth Rate Assay
5.7. Chlorophyll Amount Measurement
5.8. High-Light Tolerance Assay
5.9. Photosynthetic Parameter Analyses
5.10. PCR against Tetrad Progenies
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wakabayashi, K.; Isu, A.; Ueki, N. Channelrhodopsin-dependent photo-behavioral responses in the unicellular green alga Chlamydomonas reinhardtii. Adv. Exp. Med. Biol. 2021, 1293, 21–33. [Google Scholar]
- Harris, E.H. Motility and behavior. In The Chlamydomonas Sourcebook, 2nd ed.; Academic Press: Cambridge, MA, USA, 2009; Volume 1, pp. 89–117. [Google Scholar]
- Hartshorne, J.N. The function of the eyespot in Chlamydomonas. New Phytol. 1953, 52, 292–297. [Google Scholar] [CrossRef]
- Dieckmann, C.L. Eyespot placement and assembly in the green alga Chlamydomonas. BioEssays 2003, 25, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.W.; Smyth, R.D. Light Antennas in phototactic algae. Microbiol. Rev. 1980, 44, 572–630. [Google Scholar] [CrossRef]
- Morel-Laurens, N.M.L.; Feinleib, M.E.H. Photomovement in an “eyeless” mutant of Chlamydomonas. Photochem. Photobiol. 1983, 37, 189–194. [Google Scholar] [CrossRef]
- Ueki, N.; Ide, T.; Mochiji, S.; Kobayashi, Y.; Tokutsu, R.; Ohnishi, N.; Yamaguchi, K.; Shigenobu, S.; Tanaka, K.; Minagawa, J.; et al. Eyespot-dependent determination of the phototactic sign in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2016, 113, 5299–5304. [Google Scholar] [CrossRef] [Green Version]
- Nagel, G.; Ollig, D.; Fuhrmann, M.; Kateriya, S.; Musti, A.M.; Bamberg, E.; Hegemann, P. Channelrhodopsin-1: A light-gated proton channel in green algae. Science 2002, 296, 2395–2398. [Google Scholar] [CrossRef] [PubMed]
- Nagel, G.; Szellas, T.; Huhn, W.; Kateriya, S.; Adeishvili, N.; Berthold, P.; Ollig, D.; Hegemann, P.; Bamberg, E. Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc. Natl. Acad. Sci. USA 2003, 100, 13940–13945. [Google Scholar] [CrossRef] [Green Version]
- Nagel, G.; Szellas, T.; Kateriya, S.; Adeishvili, N.; Hegemann, P.; Bamberg, E. Channelrhodopsins: Directly light-gated cation channels. Biochem. Soc. Trans. 2005, 33, 863–866. [Google Scholar] [CrossRef]
- Kamiya, R.; Witman, G.B. Submicromolar levels of calcium control the balance of beating between the two flagella in demembranated models of Chlamydomonas. J. Cell Biol. 1984, 98, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Feinleib, M.E.H.; Curry, G.M. The relationship between stimulus intensity and oriented phototactic response (topotaxis) in Chlamydomonas. Physiol. Plant 1971, 25, 346–352. [Google Scholar] [CrossRef]
- Morel-Laurens, N. Calcium control of phototactic orientation in Chlamydomonas reinhardtii: Sign and strength of response. Photochem. Photobiol. 1987, 45, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Watanabe, M. Photosynthesis modulates the sign of phototaxis of wild-type Chlamydomonas reinhardtii. Effects of red background illumination and 3-(3′,4′-dichlorophenyl)-1,1-dimethylurea. FEBS Lett. 1993, 336, 516–520. [Google Scholar] [CrossRef] [Green Version]
- Wakabayashi, K.; Misawa, Y.; Mochiji, S.; Kamiya, R. Reduction-oxidation poise regulates the sign of phototaxis in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2011, 108, 11280–11284. [Google Scholar] [CrossRef] [Green Version]
- Ide, T.; Mochiji, S.; Ueki, N.; Yamaguchi, K.; Shigenobu, S.; Hirono, M.; Wakabayashi, K. Identification of the agg1 mutation responsible for negative phototaxis in a “wild-type” strain of Chlamydomonas reinhardtii. Biochem. Biophys. Rep. 2016, 7, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Demmig-Adams, B.; Adams, W.W. 3rd Photoprotection and other responses of plants to high light stress. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 599–626. [Google Scholar] [CrossRef]
- Niyogi, K.K. Photoprotection Revisited: Genetic and molecular approaches. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 333–359. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Xie, H.; Kang, Y.; Lin, Y.; Liu, G.; Sakato-Antoku, M.; Patel-King, R.S.; Wang, B.; Wan, C.; King, S.M.; et al. Heme-binding protein CYB5D1 is a radial spoke component required for coordinated ciliary beating. Proc. Natl. Acad. Sci. USA 2021, 118, e2015689118. [Google Scholar] [CrossRef]
- Böhm, M.; Boness, D.; Fantisch, E.; Erhard, H.; Frauenholz, J.; Kowalzyk, Z.; Marcinkowski, N.; Kateriya, S.; Hegemann, P.; Kreimer, G. Channelrhodopsin-1 phosphorylation changes with phototactic behavior and responds to physiological stimuli in Chlamydomonas. Plant Cell 2019, 31, 886–910. [Google Scholar] [CrossRef]
- Sizova, I.; Fuhrmann, M.; Hegemann, P. A Streptomyces rimosus aphVIII gene coding for a new type phosphotransferase provides stable antibiotic resistance to Chlamydomonas reinhardtii. Gene 2001, 277, 221–229. [Google Scholar] [CrossRef]
- Wakabayashi, K.; King, S.M. Modulation of Chlamydomonas reinhardtii flagellar motility by redox poise. J. Cell Biol. 2006, 173, 743–754. [Google Scholar] [CrossRef]
- Petroutsos, D.; Tokutsu, R.; Maruyama, S.; Flori, S.; Greiner, A.; Magneschi, L.; Cusant, L.; Kottke, T.; Mittag, M.; Hegemann, P.; et al. A blue-light photoreceptor mediates the feedback regulation of photosynthesis. Nature 2016, 537, 563–566. [Google Scholar] [CrossRef]
- Allorent, G.; Lefebvre-Legendre, L.; Chappuis, R.; Kuntz, M.; Truong, T.B.; Niyogi, K.K.; Ulm, R.; Goldschmidt-Clermont, M. UV-B photoreceptor-mediated protection of the photosynthetic machinery in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2016, 113, 14864–14869. [Google Scholar] [CrossRef] [Green Version]
- Sirikhachornkit, A.; Shin, J.W.; Baroli, I.; Niyogi, K.K. Replacement of alpha-tocopherol by beta-tocopherol enhances resistance to photooxidative stress in a xanthophyll-deficient strain of Chlamydomonas reinhardtii. Eukaryot Cell 2009, 8, 1648–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dent, R.M.; Haglund, C.M.; Chin, B.L.; Kobayashi, M.C.; Niyogi, K.K. Functional genomics of eukaryotic photosynthesis using insertional mutagenesis of Chlamydomonas reinhardtii. Plant Physiol. 2005, 137, 545–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, B.B.; Ledford, H.K.; Wakao, S.; Huang, S.G.; Casero, D.; Pellegrini, M.; Merchant, S.S.; Koller, A.; Eggen, R.I.; Niyogi, K.K. Singlet oxygen RESISTANT 1 links reactive electrophile signaling to singlet oxygen acclimation in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2012, 109, E1302–E1311. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.P.; Wang, M.; Wang, C. Nuclear transformation of Chlamydomonas reinhardtii: A review. Biochimie 2021, 181, 1–11. [Google Scholar] [CrossRef]
- Dayer, R.; Fischer, B.B.; Eggen, R.I.; Lemaire, S.D. The peroxiredoxin and glutathione peroxidase families in Chlamydomonas reinhardtii. Genetics 2008, 179, 41–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murchie, E.H.; Ruban, A.V. Dynamic non-photochemical quenching in plants: From molecular mechanism to productivity. Plant J. 2020, 101, 885–896. [Google Scholar] [CrossRef] [Green Version]
- Baroli, I.; Do, A.D.; Yamane, T.; Niyogi, K.K. Zeaxanthin accumulation in the absence of a functional xanthophyll cycle protects Chlamydomonas reinhardtii from photooxidative stress. Plant Cell 2003, 15, 992–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, B.B.; Eggen, R.I.; Niyogi, K.K. Characterization of singlet oxygen-accumulating mutants isolated in a screen for altered oxidative stress response in Chlamydomonas reinhardtii. BMC Plant Biol. 2010, 10, 279. [Google Scholar] [CrossRef] [Green Version]
- Tokutsu, R.; Fujiwara-Kamada, K.; Matsuo, T.; Yamasaki, T.; Minagawa, J. The CONSTANS flowering complex controls the prote4ctive response of photosynthesis in the green alga Chlamydomonas. Nat Commun. 2019, 10, 4099. [Google Scholar] [CrossRef] [PubMed]
- Gorman, D.S.; Levine, R.P. Cytochrome f and plastocyanin: Their sequence in the photosynthetic electron transport chain of Chlamydomonas reinhardi. Proc. Natl. Acad. Sci. USA 1965, 54, 1665–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sueoka, N. Mitotic replication of deoxyribonucleic acid in Chlamydomonas Reinhardi. Proc. Natl. Acad. Sci. USA 1960, 46, 83–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamano, T.; Iguchi, H.; Fukuzawa, H. Rapid transformation of Chlamydomonas reinhardtii without cell-wall removal. J. Biosci. Bioeng. 2013, 115, 691–694. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, R. Analysis of cell vibration for assessing axonemal motility in Chlamydomonas. Methods 2000, 22, 383–387. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and Simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Dang, K.V.; Plet, J.; Tolleter, D.; Jokel, M.; Cuiné, S.; Carrier, P.; Auroy, P.; Richaud, P.; Johnson, X.; Alric, J.; et al. Combined increases in mitochondrial cooperation and oxygen photoreduction compensate for deficiency in cyclic electron flow in Chlamydomonas reinhardtii. Plant Cell. 2014, 26, 3036–3050. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.; Fu, Y.; Guo, Y.; Pan, J. Chlamydomonas (Chlorophyceae) colony PCR. Protoplasma 2009, 235, 1. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT | ap1 | ap2 | ap3 | ap4 | ap5 | ap6 | ap7 | |
|---|---|---|---|---|---|---|---|---|
| Cell size (μm) * | 7.5 ± 0.1 | 7.0 ± 0.1 | 7.1 ± 0.2 | 7.3 ± 0.3 | 7.0 ± 0.1 | 7.0 ± 0.2 | 6.9 ± 0.1 | 7.4 ± 0.1 |
| Ciliary length (μm) ** | 13.2 ± 1.5 | 13.9 ± 1.6 | 13.6 ± 1.7 | 13.2 ± 1.6 | 13.7 ± 2.0 | 13.6 ± 1.6 | 13.3 ± 1.5 | 13.5 ± 1.4 |
| Ciliary beating frequency (Hz) * | 57.4 ± 0.5 | 56.0 ± 1.6 | 57.4 ± 1.1 | 56.1 ± 2.0 | 59.0 ± 0.6 | 53.1 ± 1.1 | 55.2 ± 1.2 | 54.7 ± 1.3 |
| WT | ap1 | ap2 | ap3 | ap4 | ap5 | ap6 | ap7 | |
|---|---|---|---|---|---|---|---|---|
| Chlorophyll (pg/cell) * | 0.78 ± 0.10 | 0.73 ± 0.13 | 0.82 ± 0.05 | 0.75 ± 0.07 | 0.83 ± 0.06 | 0.81 ± 0.14 | 0.70 ± 0.05 | 0.97 ± 0.07 |
| Photosynthetic efficiency (φII) ** | 0.59 ± 0.01 | 0.61 ± 0.02 | 0.59 ± 0.01 | 0.59 ± 0.02 | 0.57 ± 0.01 | 0.58 ± 0.02 | 0.59 ± 0.02 | 0.57 ± 0.01 |
| NPQ ** | 0.30 ± 0.02 | 0.30 ± 0.02 | 0.31 ± 0.01 | 0.29 ± 0.04 | 0.29 ± 0.03 | 0.28 ± 0.01 | 0.29 ± 0.01 | 0.28 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morishita, J.; Tokutsu, R.; Minagawa, J.; Hisabori, T.; Wakabayashi, K.-i. Characterization of Chlamydomonas reinhardtii Mutants That Exhibit Strong Positive Phototaxis. Plants 2021, 10, 1483. https://doi.org/10.3390/plants10071483
Morishita J, Tokutsu R, Minagawa J, Hisabori T, Wakabayashi K-i. Characterization of Chlamydomonas reinhardtii Mutants That Exhibit Strong Positive Phototaxis. Plants. 2021; 10(7):1483. https://doi.org/10.3390/plants10071483
Chicago/Turabian StyleMorishita, Jun, Ryutaro Tokutsu, Jun Minagawa, Toru Hisabori, and Ken-ichi Wakabayashi. 2021. "Characterization of Chlamydomonas reinhardtii Mutants That Exhibit Strong Positive Phototaxis" Plants 10, no. 7: 1483. https://doi.org/10.3390/plants10071483
APA StyleMorishita, J., Tokutsu, R., Minagawa, J., Hisabori, T., & Wakabayashi, K.-i. (2021). Characterization of Chlamydomonas reinhardtii Mutants That Exhibit Strong Positive Phototaxis. Plants, 10(7), 1483. https://doi.org/10.3390/plants10071483