
Antimicrobial Activity of Extracts from the Humiria balsamifera (Aubl)


 , ,
, , 
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Activity Evaluation
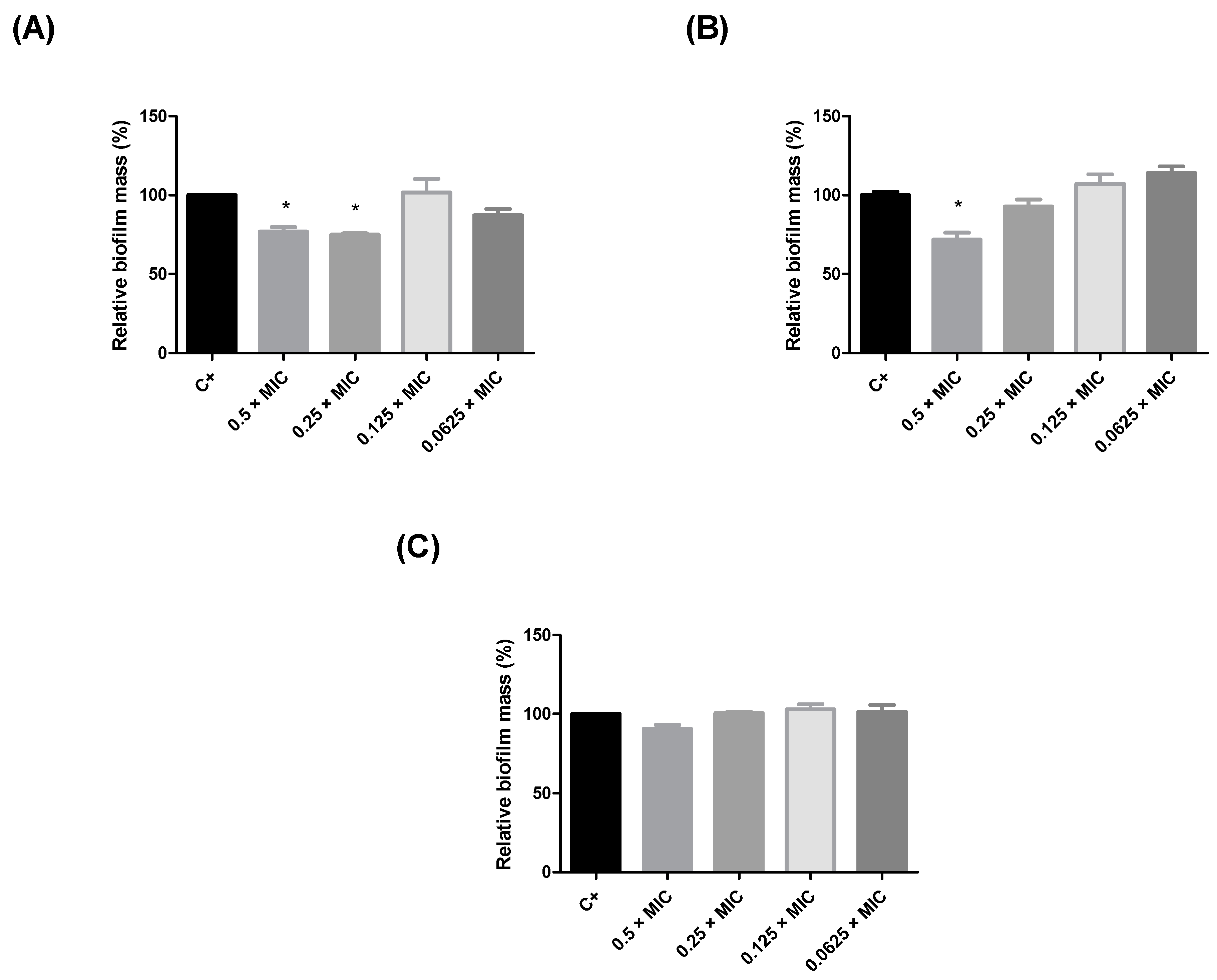
2.2. Evaluation of the Antibiofilm Activity of Humiria balsamifera (Aubl) Extracts
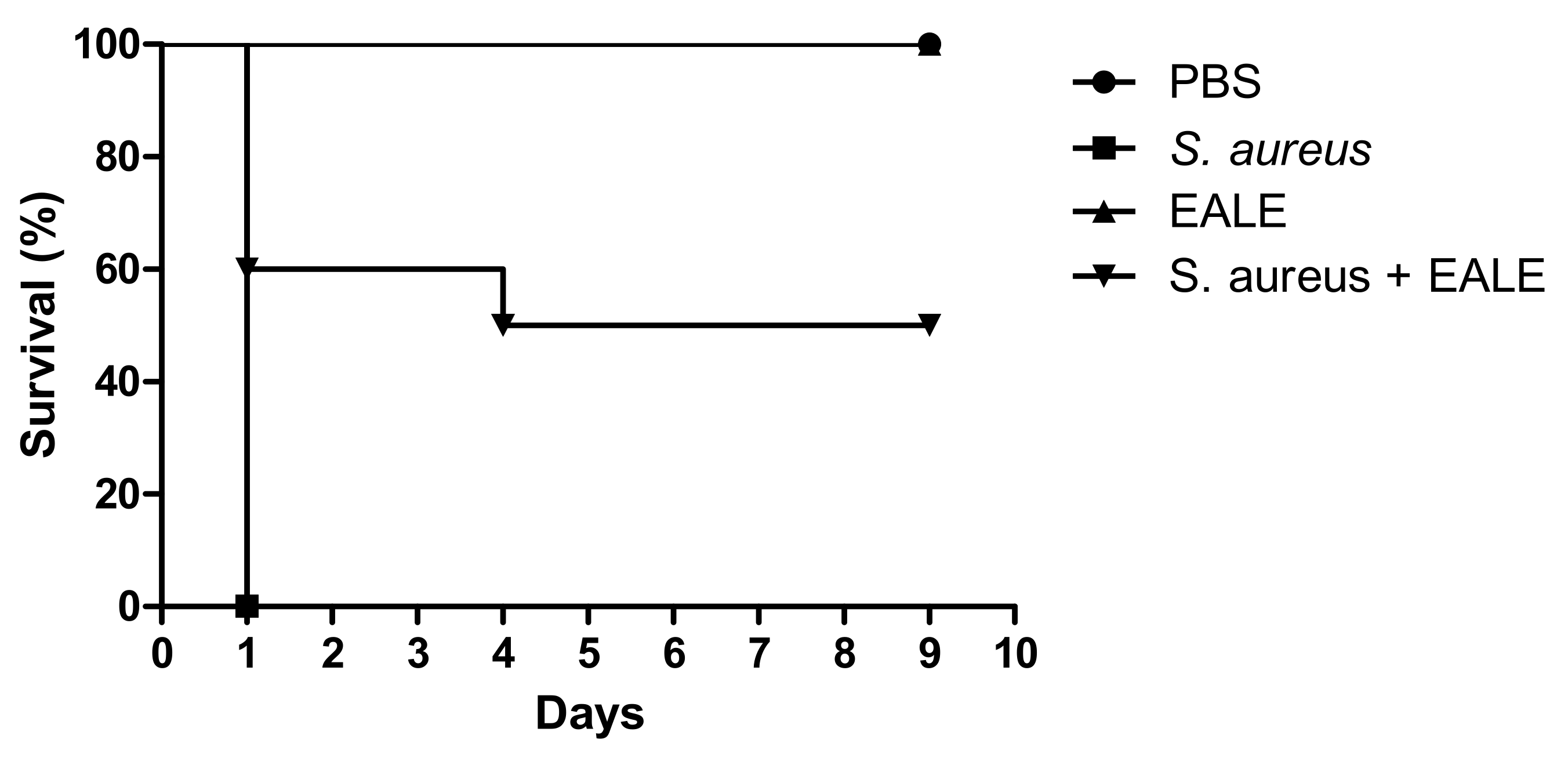
2.3. Evaluation of the In Vivo Activity of the Ethyl Acetate Leaf Extract of Humiria balsamifera (Aubl)
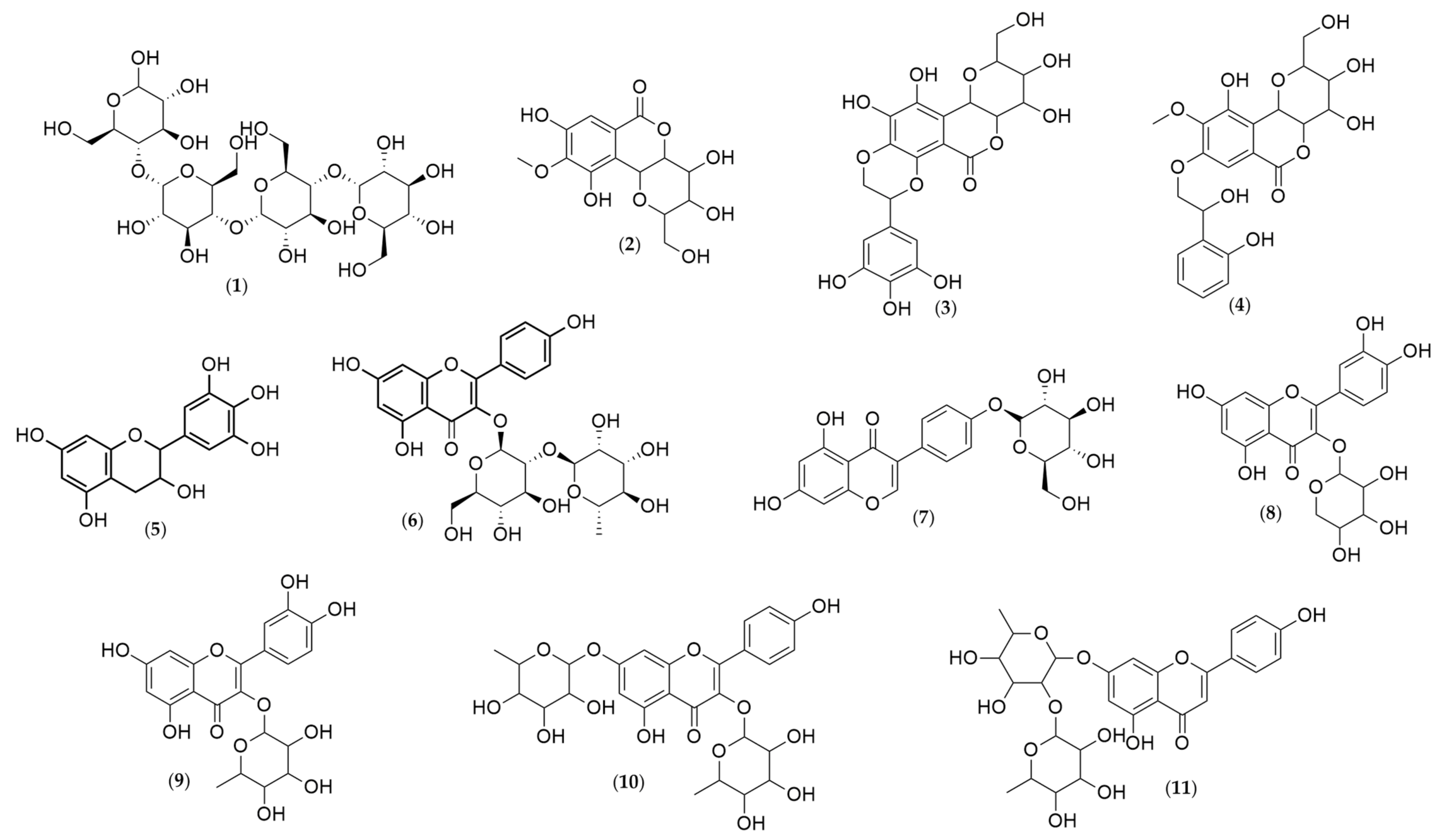
2.4. Chemical Characterization of Humiria balsamifera (Aubl) Leaf and Stem Bark Extracts
3. Discussion
4. Materials and Methods
4.1. Botanical Material
4.2. Preparation of the Extracts
4.3. Test Microorganisms
4.4. Minimum Inhibitory Concentration (MIC)
4.5. Antibiofilm Test
4.6. Infection Model Using Tenebrio molitor Larvae
4.7. Extracts Characterization by HPLC-ESI-MS and FIA-ESI-IT/MS
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gonelimali, F.D.; Lin, J.; Miao, W.; Xuan, J.; Charles, F.; Chen, M.; Hatab, S.R. Antimicrobial Properties and Mechanism of Action of Some Plant Extracts Against Food Pathogens and Spoilage Microorganisms. Front. Microbiol. 2018, 9, 1–9. [Google Scholar] [CrossRef]
- Franz, C.M.A.P.; Den Besten, H.M.W.; Böhnlein, C.; Gareis, M.; Zwietering, M.H.; Fusco, V. Microbial food safety in the 21st century: Emerging challenges and foodborne pathogenic bactéria. Trends Food Sci. Technol. 2018, 81, 155–158. [Google Scholar] [CrossRef]
- Parlapani, F.F.; Mallouchos, A.; Haroutounian, S.A.; Boziaris, I.S. Volatile organic compounds of microbial and non-microbial origin produced on model fish substrate un-inoculated and inoculated with gilt-head sea bream spoilage bacteria. LWT Food Sci. Technol. 2018, 78, 54–62. [Google Scholar] [CrossRef]
- Lianou, A.; Panagou, E.Z.; Nychas, G.-J.E. Microbiological spoilage of foods and beverages. In The Stability and Shelf Life of Food, 2nd ed.; Subramaniam, P., Ed.; Woodhead Publishing: Cambridge, UK, 2016; pp. 3–42. [Google Scholar] [CrossRef]
- Nahar, S.; Jeong, H.L.; Kim, Y.; Ha, A.J.; Roy, P.K.; Park, S.H.; Ashrafudoulla, M.; Mizan, M.F.R.; Ha, S. Inhibitory effects of Flavourzyme on biofilm formation, quorum sensing, and virulence genes of foodborne pathogens Salmonella Typhimurium and Escherichia coli. Food Res. Int. 2021, 147, 110461. [Google Scholar] [CrossRef]
- WHO (World Health Organisation). WHO Estimates of the Global Burden of Foodborne Diseases Foodborne Diseases Burden Epidemiology Reference Group 2007–2015. 2015. Available online: https://www.who.int/en/news-room/fact-sheets/detail/food-safety (accessed on 1 June 2021).
- Moyer, C.L.; Morita, R.Y. Psychrophiles and psychrotrophs. In Encyclopedia of Life Sciences; John Wiley & Sons Ltd.: Chichester, UK, 2007; pp. 1–6. [Google Scholar] [CrossRef]
- Wei, Q.; Wang, X.; Sun, D.; Pu, H. Rapid detection and control of psychrotrophic microorganisms in cold storage foods: A review. Trends Food Sci. Technol. 2019, 86, 453–464. [Google Scholar] [CrossRef]
- Kirk, M.D.; Ângulo, F.J.; Havelaar, A.H.; Black, R.E. Diarrhoeal disease in children due to contaminated food. Bull. World Health Organ. 2017, 95, 233–234. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.J.; Kendall, M.M. Salmonella typhimurium Strategies for Host Adaptation. Front. Microbiol. 2017, 8, 1983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neri, D.; Antoci, S.; Iannetti, L.; Ciorba, A.B.; D’Aurelio, R.; Del Matto, I.; Di Leonardo, M.; Giovannini, A.; Prencipe, V.A.; Pomilio, F.; et al. EU and US control measures on Listeria monocytogenes and Salmonella spp. in certain ready-to-eat meat products: An equivalence study. Food Control 2019, 96, 98–103. [Google Scholar] [CrossRef]
- Shahid, A.H.; Nazir, K.H.M.; El Zowalaty, M.E.; Kabir, A.; Sarker, S.A.; Siddique, M.P.; Ashour, H.M. Molecular detection of vancomycin resistance and methicillin resistance in Staphylococcus aureus from food processing environments. One Health 2021, 100276, 1–14. [Google Scholar] [CrossRef]
- Bello, A.; Dingle, T.C. What’s That Resistance Mechanism? Understanding Genetic Determinants of Gram-Negative Bacterial Resistance. Clin. Microbiol. Newsl. 2018, 40, 165–174. [Google Scholar] [CrossRef]
- Holmes, A.H.; Moore, L.S.P.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddock, L.J.V. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Chandra, H.; Bishnoi, P.; Yadav, A.; Patni, B.; Mishra, A.P.; Nautiyal, A.R. Antimicrobial Resistance and the Alternative Resources with Special Emphasis on Plant-Based Antimicrobials—A Review. Plants 2017, 6, 16. [Google Scholar] [CrossRef]
- Da Silva, L.C.N.; Da Silva, M.V.; Correia, M.T.S. New Frontiers in the Search of Antimicrobials Agents from Natural Products. Front. Microbiol. 2017, 8, 210. [Google Scholar] [CrossRef] [PubMed]
- Valli, M.; Bolzani, V.S. Natural Products: Perspectives and Challenges for use of Brazilian Plant Species in the Bioeconomy. An. Acad. Bras. Cienc. 2019, 91, e20190208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, T.; Reker, D.; Schneider, P.; Schneider, G. Counting on natural products for drug design. Nat. Chem. 2016, 8, 531–541. [Google Scholar] [CrossRef]
- Imai, Y.; Meyer, K.J.; Iinishi, A. A new antibiotic selectively kills Gram-negative pathogens. Nature 2019, 576, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Melander, R.J.; Basak, A.K.; Melander, C. Natural products as inspiration for the development of bacterial antibiofilm agents. Nat. Prod. Rep. 2020, 11. [Google Scholar] [CrossRef]
- Carneiro, D.B.; Barboza, M.S.L.; Menezes, M.P. Plantas nativas úteis na Vila dos Pescadores da Reserva Extrativista Marinha Caeté-Taperaçu, Pará, Brasil. Acta Botânica Bras. 2010, 24, 1027–1033. [Google Scholar] [CrossRef]
- Santos, M.G.; Fevereiro, P.C.A.; Reis, G.L.; Barcelos, J.I. Recursos vegetais da restinga de Carapebus, Rio de Janeiro, Brasil. J. Neotrop. Biol. 2009, 6, 35–54. [Google Scholar] [CrossRef]
- Silva, T.B.C.; Alves, V.L.; Mendonça, L.V.H.; Conserva, L.M.; Rocha, E.M.M.; Andrade, E.H.A.; Lemos, R.P.L. Chemical Constituents and Preliminary Antimalarial Activity of Humiria balsamifera. Pharm. Biol. 2004, 42, 94–97. [Google Scholar] [CrossRef] [Green Version]
- Paz, S.A.; Marques, G.E.C. Formulação de Hidratante com Extratos de Humiria balsamifera (Aubl A. St. Hil) com Ação Antioxidante. In Cadernos de Agroecologia; Anais do XI Congresso Brasileiro de Agroecologia; ABA—Associação Brasileira de Agroecologia: Rio de Janeiro, Brasil, 2020; Volume 15, p. 2. Available online: http://cadernos.aba-agroecologia.org.br/index.php/cadernos/issue/view/4 (accessed on 1 May 2021).
- Dutra, A.R.; Sutilli, F.K.; Muzitano, M.F.; Guimarães, D.O.; Souza, R.O.; Leal, I.C. Microwave-assisted extraction of metabolites from Humiria balsamifera leaves: A rapid and efficient methodology for antioxidant constituents obtainment. Planta Med. 2013, 79. [Google Scholar] [CrossRef]
- Lima, C.N.S.; Farias, F.A.C.; Marques, G.E.C. Atividade Antimicrobiana in vitro dos Extratos de Humiria balsamífera (Aulb) A. St. Hil. Frente a Cepas Bacaterianas e Fúngicas. In Mirim (Humiria balsamifera (Aubl.): Riqueza da Biodiversidade Maranhense, 1st ed.; Luís, S., Ed.; IFMA: São Luís, Brazil, 2019; Volume 1, pp. 99–107. Available online: https://editora.ifma.edu.br/livraria-edifma-2019/ (accessed on 2 May 2021).
- Tüting, W.; Adden, R.; Mischnick, P. Fragmentation pattern of regioselectively O-methylated maltooligosaccharides in electrospray ionisation-mass spectrometry/collision induced dissociation. Int. J. Mass Spectrom. 2004, 232, 107–115. [Google Scholar] [CrossRef]
- Song, H.; Wang, J.; Zhang, R.; Liu, X.; Yuan, G.; Wei, C.; Zhao, W.; Li, R.; Wang, B.; Guo, R. In vivo metabolism study of bergenin in rats by HPLC-QTOF mass spectrometry. Biomed. Chromatogr. 2013, 27, 1398–1405. [Google Scholar] [CrossRef] [PubMed]
- Callemien, D.; Collin, S. Use of RP-HPLC-ESI (–)—MS/MS to Differentiate Various Proanthocyanidin Isomers in Lager Beer Extracts. J. Am. Soc. Brew. Chem. 2008, 66, 109–115. [Google Scholar] [CrossRef]
- March, R.E.; Miao, X. A fragmentation study of kaempferol using electrospray quadrupole time-of-flight mass spectrometry at high mass resolution. Int. J. Mass Spectrom. 2004, 231, 157–167. [Google Scholar] [CrossRef]
- Ablajan, K. A study of characteristic fragmentation of isoflavonoids by using negative ion ESI-MSn. Int. J. Mass Spectrom. 2011, 46, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Ćirić, A.; Prosen, H.; Jelikić-Stankov, M.; Đurđević, P. Evaluation of matrix effect in determination of some bioflavonoids in food samples by LC–MS/MS method. Talanta 2012, 99, 780–790. [Google Scholar] [CrossRef]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Ng, K.R.; Lyu, X.; Mark, R.; Chen, W.N. Antimicrobial and antioxidant activities of phenolic metabolites from flavonoid-producing yeast: Potential as natural food preservatives. Food Chem. 2019, 270, 123–129. [Google Scholar] [CrossRef]
- Lalani, S.; Poh, C.L. Flavonoids as Antiviral Agents for Enterovirus A71 (EV-A71). Viruses 2020, 12, 184. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Wang, D.; Song, X. Natural Prenylchalconaringenins and Prenylnaringenins as Antidiabetic Agents: α-Glucosidase and α-Amylase Inhibition and in Vivo Antihyperglycemic and Antihyperlipidemic Effects. J. Agric. Food Chem. 2017, 65, 1574–1581. [Google Scholar] [CrossRef]
- Samy, R.P.; Gopalakrishnakone, P. Therapeutic Potential of Plants as Anti-microbials for Drug Discovery. Evid. Based Complementary Altern. Med. 2010, 7, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Buer, C.S.; Imin, N.; Djordjevic, M.A. Flavonoids: New roles for old molecules. J. Integr. Plant Biol. 2010, 52, 98–111. [Google Scholar] [CrossRef]
- Petrussa, E.; Braidot, E.; Zancani, M.; Peresson, C.; Bertolini, A.; Patui, S.; Vianello, A. Plant flavonoids-biosynthesis, transport and involvement in stress responses. Int. J. Mol. Med. Sci. 2013, 14, 14950–14973. [Google Scholar] [CrossRef]
- Biharee, A.; Sharma, A.; Kumar, A.; Jaitak, V. Antimicrobial flavonoids as a potential substitute for overcoming antimicrobial resistance. Fitoterapia 2020, 146, 104720. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Solnier, J.; Martin, L.; Bhakta, S.; Bucar, F. Flavonoids as Novel Efflux Pump Inhibitors and Antimicrobials Against Both Environmental and Pathogenic Intracellular Mycobacterial Species. Molecules 2020, 25, 734. [Google Scholar] [CrossRef] [Green Version]
- Araya-Cloutier, C.; Vincken, J.; Ederen, R.V.; Den Besten, H.M.W.; Gruppen, H. Rapid membrane permeabilization of Listeria monocytogenes and Escherichia coli induced by antibacterial prenylated phenolic compounds from legumes. Food Chem. 2018, 240, 147–155. [Google Scholar] [CrossRef]
- Silva, S.L.; Oliveira, V.G.; Yano, T.; Nunomura, R.C.S. Atividade antimicrobiana da bergenina de Endopleura uchi (Huber) Cuatrec. Acta Amaz. 2009, 39, 187–191. [Google Scholar] [CrossRef] [Green Version]
- Maduka, H.C.C.; Okoye, Z.S.C.; Eje, A. The influence of Sacoglottis gabonensis stem bark extract and its isolate bergenin, Nigerian alcoholic beverage additives, on the metabolic and haematological side effects of 2,4-dinitrophenyl hydrazine-induced tissue damage. Vasc. Pharmacol. 2003, 39, 317–324. [Google Scholar] [CrossRef]
- Silva Neto, O.C.; Teodoro, M.T.F.; Nascimento, B.O. Bergenin of Peltophorum dubium (Fabaceae) Roots and Its Bioactive Semi-Synthetic Derivatives. J. Braz. Chem. Soc. 2020, 31, 2644–2650. [Google Scholar] [CrossRef]
- Nunomura, R.C.S.; Oliveira, V.G.; Silva, S.L.; Nunomura, S.M. Characterization of bergenin in Endopleura uchi bark and its anti-inflammatory activity. J. Braz. Chem. Soc. 2009, 20, 1060–1064. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.R.; Arfan, M.; Amin, H. Synthesis of new bergenin derivatives as potent inhibitors of inflammatory mediators NO and TNF-α. Bioorganic Med. Chem. Lett. 2012, 22, 2744–2747. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Nonao, F.B.; Lima, F.O. Antinociceptive Properties of Bergenin. J. Nat. Prod. 2011, 74, 2062–2068. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Kumar, A.; Sharma, A. Antianxiety activity guided isolation and characterization of bergenin from Caesalpinia digyna Rottler roots. J. Ethnopharmacol. 2017, 195, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Uddin, G.; Sadat, A.; Siddiqui, B.S. Comparative antioxidant and antiplasmodial activities of 11-O-galloylbergenin and bergenin isolated from Bergenia ligulata. Trop. Biomed. 2014, 31, 143–148. [Google Scholar] [CrossRef]
- Muniz, M.P.; Nunomura, S.M.; Lima, E.S.; Lima, A.S.; Almeida, P.D.O.; Nunomura, R.C.S. Quantification of bergenin, antioxidant activity and nitric oxide inhibition from bark, leaf and twig of Endopleura uchi. Química Nova 2020, 43, 413–418. [Google Scholar] [CrossRef]
- Cantanhede Filho, A.J.; Santos, L.S.; Guilhon, G.M.S.P.; Zoghbi, M.G.B.; Ports, P.S.; Rodrigues, I.C.S. Triterpenoides, Fenólicos e Efeito Fitotóxico de Eugenia flavescens DC (Myrtaceae). Química Nova 2017, 40, 252–259. [Google Scholar] [CrossRef]
- Da Fonseca Amorim, E.A.; Castro, E.J.M.; da Souza, S.V.; Alves, M.S.; Dias, L.R.L.; Melo, M.H.F.; de Miranda, R.D.C.M. Antimicrobial Potential of Streptomyces ansochromogenes (PB3) Isolated from a Plant Native to the Amazon Against Pseudomonas aeruginosa. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Serra, M.B.; Barroso, W.A.; Rocha, C.; Furtado, P.G.R.; Borges, A.C.R.; Silva, S.N.; Tangerina, M.M.P.; Nascimento, J.R.; Vilegas, W.; Alves, A.C.; et al. Chemical Characterization and Wound Healing Property of Jacaranda decurrens Cham. (Bignoniaceae): An Experimental Study Based on Molecular Mechanisms. Evidence-Based. Complementary Altern. Med. 2020. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Bacteria Species | HSBE | EASBE | MSBE | HLE | EALE | MLE |
|---|---|---|---|---|---|---|
| E. coli ATCC 25922 | >12,500 | >12,500 | >12,500 | >12,500 | >12,500 | >12,500 |
| L. monocytogenes ATCC 6538 | >12,500 | >12,500 | >12,500 | >12,500 | 3120 | 3120 |
| S. aureus ATCC 6538 | >12,500 | >12,500 | >12,500 | 1560 | 780 | 780 |
| S. enterica Typhimurium ATCC 14028 | >12,500 | >12,500 | >12,500 | >12,500 | >12,500 | >12,500 |
| RT (min) | [M-H] | MSn | Proposed Substance | Reference |
|---|---|---|---|---|
| 1.97 | 665 | 664, 305 | Maltotetraose | [28] |
| 2.85 | 327 | 249, 234, 207 | Bergenin | [29] |
| 3.58 | 479 | 327, 249, 234, 207 | Galloylbergenin | [29] |
| 4.87 | 463 | 327, 249, 234, 207 | Hydroxybenzoyl bergenin | [29] |
| RT (min) | [M-H] | MSn | Proposed Substance | Reference |
|---|---|---|---|---|
| 3.01 | 305 | 179, 165 | Gallocatechin | [30] |
| 3.81 | 327 | 234, 207, 192 | Bergenin | [29] |
| 6.35 | 593 | 431, 411 | Kaempferol 3-neohesperidoside | [31] |
| RT (min) | [M-H] | MSn | Proposed Substance | Reference |
|---|---|---|---|---|
| 2.83 | 327 | 249, 234, 207, 192 | Bergenin | [29] |
| 5.27 | 431 | 311, 283, 341 | Sophoricoside | [32] |
| 6.09 | 433 | 300, 271, 243, 227 | Quercetin 3-arabinoside | [33] |
| 6.46 | 447 | 300, 271, 243 | Quercetin-O-rhamnoside | [33] |
| 7.17 | 577 | 431, 413, 341, 300 | Kaempferol-dirhamnoside | [33] |
| 8.38 | 561 | 431, 293 | Vitexin-dirhamnoside | [32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dias, E.d.J.S.; Cantanhede Filho, A.J.; Carneiro, F.J.C.; da Rocha, C.Q.; da Silva, L.C.N.; Santos, J.C.B.; Barros, T.F.; Santos, D.M. Antimicrobial Activity of Extracts from the Humiria balsamifera (Aubl). Plants 2021, 10, 1479. https://doi.org/10.3390/plants10071479
Dias EdJS, Cantanhede Filho AJ, Carneiro FJC, da Rocha CQ, da Silva LCN, Santos JCB, Barros TF, Santos DM. Antimicrobial Activity of Extracts from the Humiria balsamifera (Aubl). Plants. 2021; 10(7):1479. https://doi.org/10.3390/plants10071479
Chicago/Turabian StyleDias, Edelson de J. S., Antônio J. Cantanhede Filho, Fernando J. C. Carneiro, Cláudia Q. da Rocha, Luís Cláudio N. da Silva, Joice C. B. Santos, Thayná F. Barros, and Deivid M. Santos. 2021. "Antimicrobial Activity of Extracts from the Humiria balsamifera (Aubl)" Plants 10, no. 7: 1479. https://doi.org/10.3390/plants10071479
APA StyleDias, E. d. J. S., Cantanhede Filho, A. J., Carneiro, F. J. C., da Rocha, C. Q., da Silva, L. C. N., Santos, J. C. B., Barros, T. F., & Santos, D. M. (2021). Antimicrobial Activity of Extracts from the Humiria balsamifera (Aubl). Plants, 10(7), 1479. https://doi.org/10.3390/plants10071479