
Ultrastructural Alterations in Cells of Sunflower Linear Glandular Trichomes during Maturation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
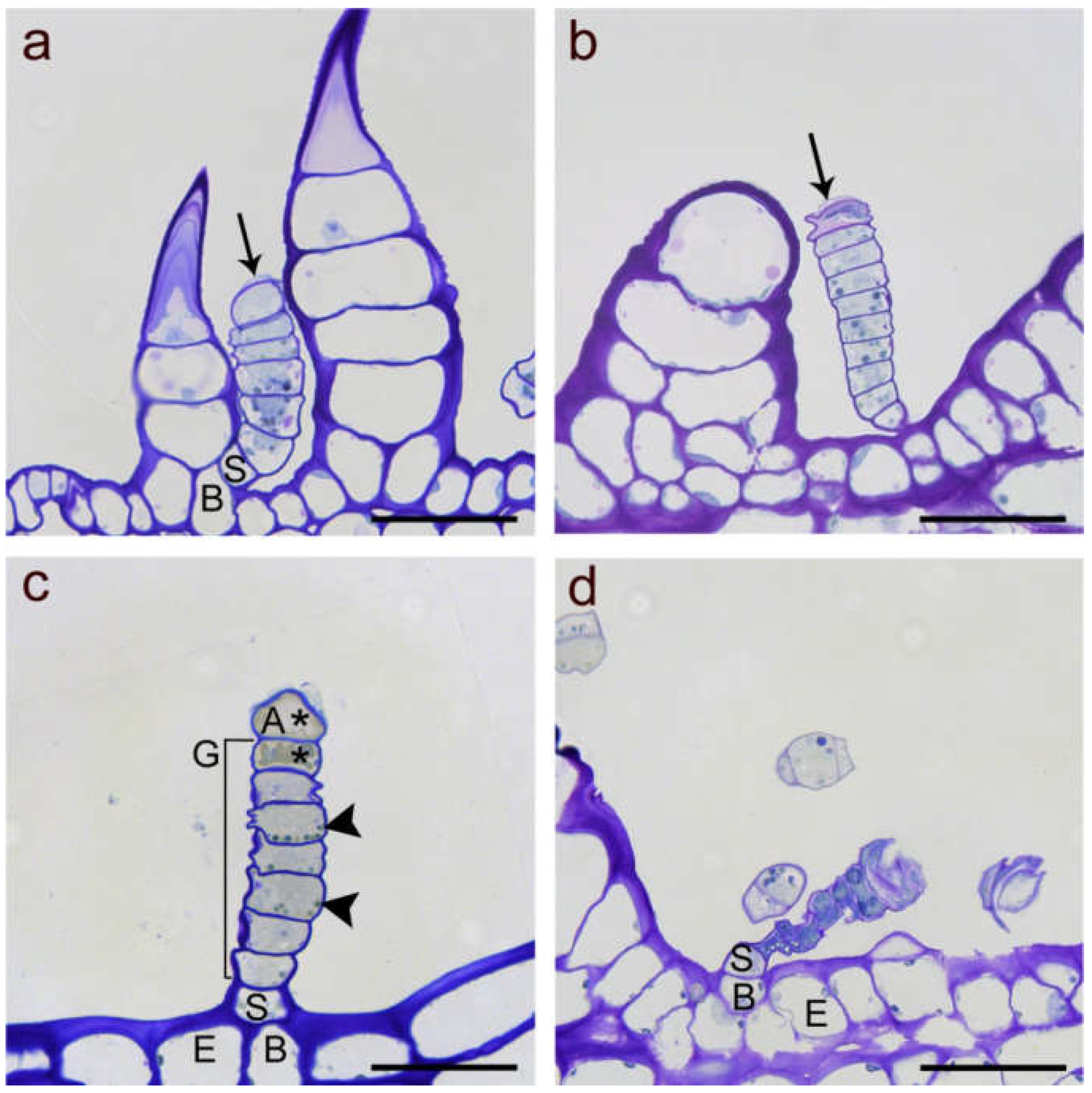
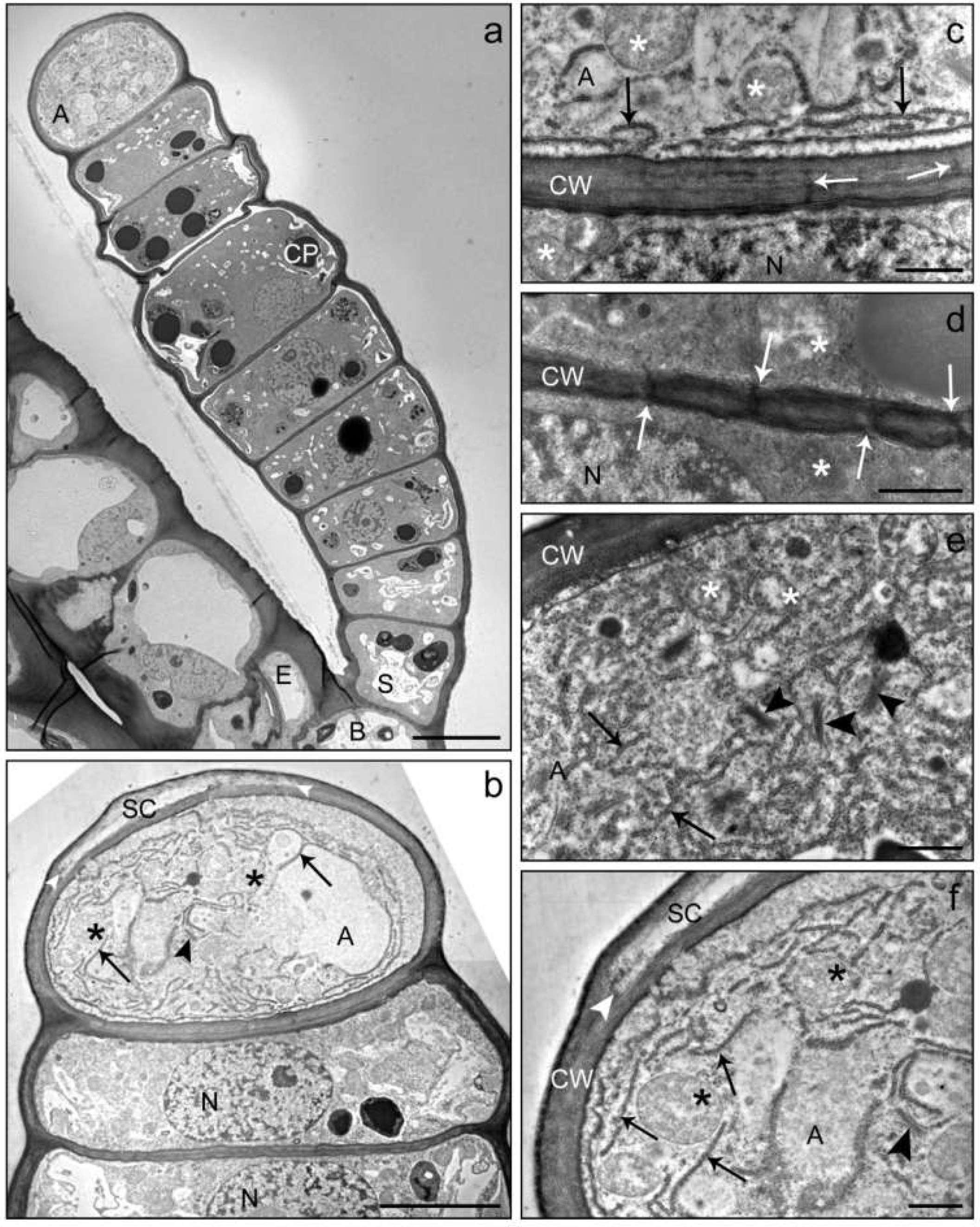
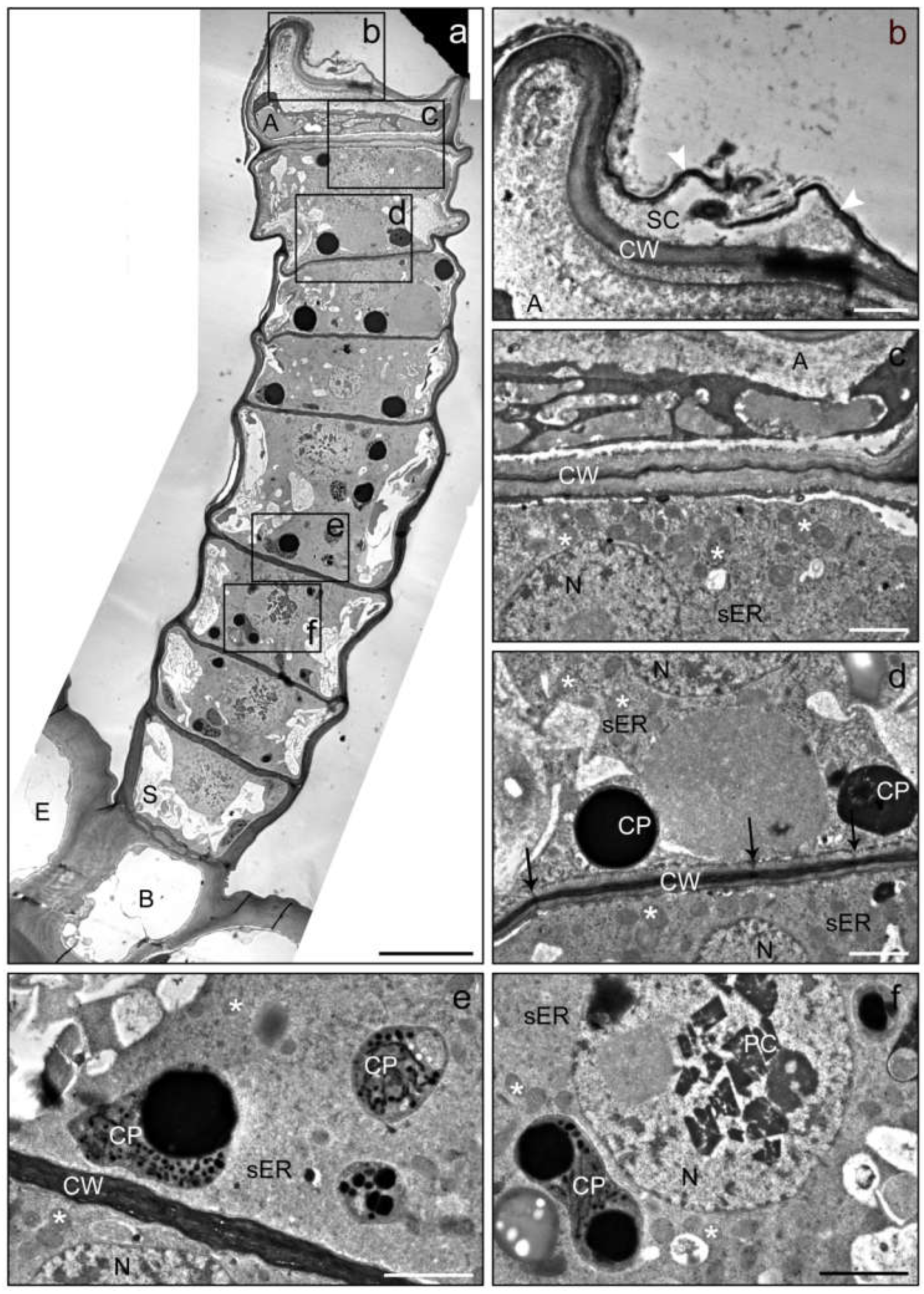
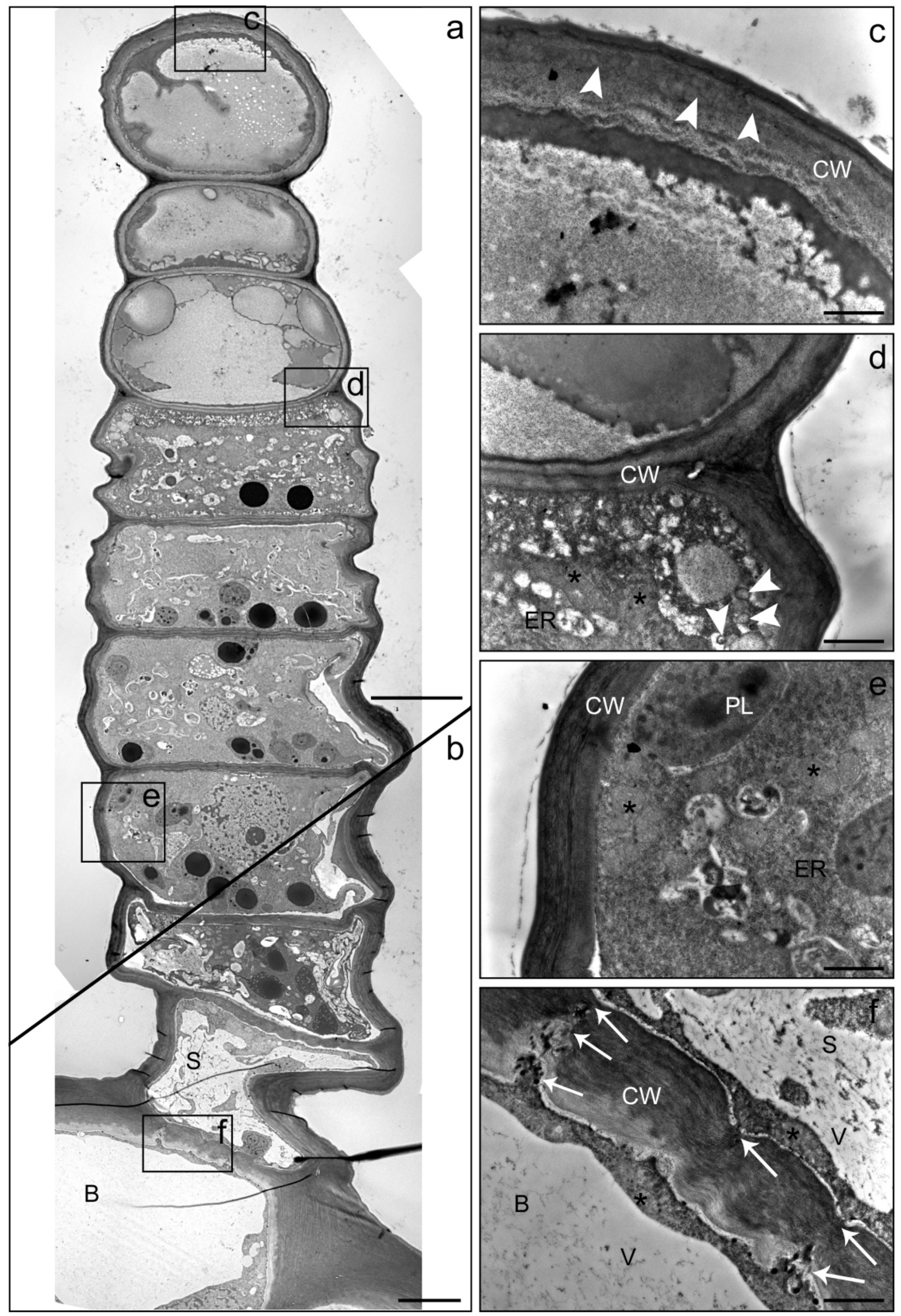
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material and Cultivation
4.2. Sample Preparation and Microscopy
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wagner, G.J.; Wang, E.; Shepherd, R.W. New approaches for studying and exploiting an old protuberance, the plant trichome. Ann. Bot. 2004, 93, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Werker, E. Trichome diversity and development. Adv. Bot. Res. 2000, 31, 1–35. [Google Scholar] [CrossRef]
- Muravnik, L.E. The structural peculiarities of the leaf glandular trichomes: A review. In Plant Cell and Tissue Differentiation and Secondary Metabolites, Reference Series in Phytochemistry; Ramawat, K.G., Ekiert, H.M., Goyal, S., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 1–34. [Google Scholar] [CrossRef]
- Dai, X.; Wang, G.; Yang, D.S.; Tang, Y.; Broun, P.; Marks, M.D.; Sumner, L.W.; Dixon, R.A.; Zhao, P.X. TrichOME: A comparative omics database for plant trichomes. Plant Physiol. 2010, 152, 44–54. [Google Scholar] [CrossRef] [Green Version]
- Croteau, R.B.; Davis, E.M.; Ringer, K.L.; Wildung, M.R. (−)-Menthol biosynthesis and molecular genetics. Naturwissenschaften 2005, 92, 562–577. [Google Scholar] [CrossRef] [PubMed]
- Duke, M.V.; Paul, R.N.; Elsohly, H.N.; Sturtz, G.; Duke, S.O. Localization of artemisinin and artemisitene in foliar tissues of glanded and glandless biotypes of Artemisia annua. Int. J. Plant Sci. 1994, 155, 365–373. [Google Scholar] [CrossRef]
- Spring, O.; Bienert, U.; Klemt, V. Sesquiterpene lactones in glandular trichomes of sunflower leaves. J. Plant Physiol. 1987, 130, 433–439. [Google Scholar] [CrossRef]
- Aschenbrenner, A.K.; Horakh, S.; Spring, O. Linear glandular trichomes of Helianthus (Asteraceae): Morphology, localization, metabolite activity and occurrence. AoB Plants 2013, 5, 1–9. [Google Scholar] [CrossRef]
- Göpfert, J.C.; Heil, N.; Conrad, J.; Spring, O. Cytological development and sesquiterpene lactone secretion in capitate glandular trichomes of sunflower. Plant Biol. 2005, 7, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Aschenbrenner, A.-K.; Amrehn, E.; Bechtel, L.; Spring, O. Trichome differentiation on leaf primordia of Helianthus annuus (Asteraceae): Morphology, gene expression and metabolite profile. Planta 2015, 241, 837–846. [Google Scholar] [CrossRef]
- Spring, O. Chemotaxonomy based on metabolites from glandular trichomes. Adv. Bot. Res. 2000, 31, 153–174. [Google Scholar]
- Amrehn, E.; Heller, A.; Spring, O. Capitate glandular trichomes of Helianthus annuus (Asteraceae): Ultrastructure and cytological development. Protoplasma 2014, 251, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Göpfert, J.C.; MacNevin, G.; Ro, D.K.; Spring, O. Identification, functional characterization and developmental regulation of sesquiterpene synthases from sunflower capitate glandular trichomes. BMC Plant Biol. 2009, 9, 86. [Google Scholar] [CrossRef] [Green Version]
- Spring, O.; Kupka, J.; Maier, B.; Hager, A. Biological activities of sesquiterpene lactones from Helianthus annuus: Antimicrobial and cytotoxic properties; influence on DNA, RNA, and protein synthesis. Z. Naturforsch. Sect. C 1982, 37, 1087–1091. [Google Scholar] [CrossRef] [PubMed]
- Prasifka, J.R.; Spring, O.; Conrad, J.; Cook, L.W.; Palmquist, D.E.; Foley, M.E. Sesquiterpene lactone composition of wild and cultivated sunflowers and biological activity against an insect pest. J. Agric. Food Chem. 2015, 63, 4042–4049. [Google Scholar] [CrossRef]
- Spring, O.; Rodon, U.; Macias, F.A. Sesquiterpenes from non-capitate glandular trichomes of Helianthus annuus L. Phytochemistry 1992, 31, 1541–1544. [Google Scholar] [CrossRef]
- Brentan Silva, D.; Aschenbrenner, A.K.; Lopes, N.P.; Spring, O. Direct analyses of secondary metabolites by mass spectrometry imaging (MSI) from sunflower (Helianthus annuus L.) trichomes. Molecules 2017, 22, 774. [Google Scholar] [CrossRef] [PubMed]
- Spring, O.; Pfannstiel, J.; Klaiber, I.; Conrad, J.; Beifuß, U.; Apel, L.; Aschenbrenner, A.-K.; Zipper, R. The nonvolatile metabolome of sunflower linear glandular trichomes. Phytochemistry 2015, 119, 83–89. [Google Scholar] [CrossRef]
- Ascensao, L.; Pais, M.S. The leaf capitate trichomes of Leonotis leonurus: Histochemistry, ultrastructure and secretion. Ann. Bot. 1998, 81, 263–271. [Google Scholar] [CrossRef]
- Spring, O. Microsampling: An alternative approach using sesquiterpene lactones for systematics. Biochem. Syst. Ecol. 1989, 17, 509–517. [Google Scholar] [CrossRef]
- Giuliani, C.; Ascrizzi, R.; Corrà, S.; Maleci, B.L.; Flamini, G.; Fico, G. Salvia uliginosa Benth: Glandular trichomes as bio-factories of volatiles and essential oil. Flora 2017, 233, 12–21. [Google Scholar] [CrossRef]
- Waigmann, E.; Zambryski, P. Trichome plasmodesmata: A model system for cell-to-cell movement. Adv. Bot. Res. 2000, 31, 261–283. [Google Scholar]
- Lange, B.M.; Turner, G.W. Terpenoid biosynthesis in trichomes—Current status and future opportunities. Plant Biotechnol. J. 2013, 11, 2–22. [Google Scholar] [CrossRef]
- De Vargas, W.; Fortuna-Perez, A.P.; Lewis, G.P.; Piva, T.C.; Vatanparast, M.; Machado, S.R. Ultrastructure and secretion of glandular trichomes in species of subtribe Cajaninae Benth (Leguminosae, Phaseoleae). Protoplasma 2019, 256, 431–445. [Google Scholar] [CrossRef]
- Tissier, A. Glandular trichomes: What comes after expressed sequence tags? Plant J. 2012, 70, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Huchelmann, A.; Boutry, M.; Hachez, C. Plant glandular trichomes: Natural cell factories of high biotechnological interest. Plant Physiol. 2017, 175, 6–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey, M.; Klaiber, I.; Conrad, J.; Spring, O. CYP71BL9, the missing link in costunolide synthesis of sunflower. Phytochemistry 2020, 177, 112430. [Google Scholar] [CrossRef] [PubMed]
- Amrehn, E.; Aschenbrenner, A.-K.; Heller, A.; Spring, O. Localization of sesquiterpene lactone biosynthesis in cells of capitate glandular trichomes of Helianthus annuus (Asteraceae). Protoplasma 2016, 253, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, E.S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amrehn, E.; Spring, O. Ultrastructural Alterations in Cells of Sunflower Linear Glandular Trichomes during Maturation. Plants 2021, 10, 1515. https://doi.org/10.3390/plants10081515
Amrehn E, Spring O. Ultrastructural Alterations in Cells of Sunflower Linear Glandular Trichomes during Maturation. Plants. 2021; 10(8):1515. https://doi.org/10.3390/plants10081515
Chicago/Turabian StyleAmrehn, Evelyn, and Otmar Spring. 2021. "Ultrastructural Alterations in Cells of Sunflower Linear Glandular Trichomes during Maturation" Plants 10, no. 8: 1515. https://doi.org/10.3390/plants10081515
APA StyleAmrehn, E., & Spring, O. (2021). Ultrastructural Alterations in Cells of Sunflower Linear Glandular Trichomes during Maturation. Plants, 10(8), 1515. https://doi.org/10.3390/plants10081515