
The Chloroplast Phylogenomics and Systematics of Zoysia (Poaceae)


Abstract
:1. Introduction
2. Results
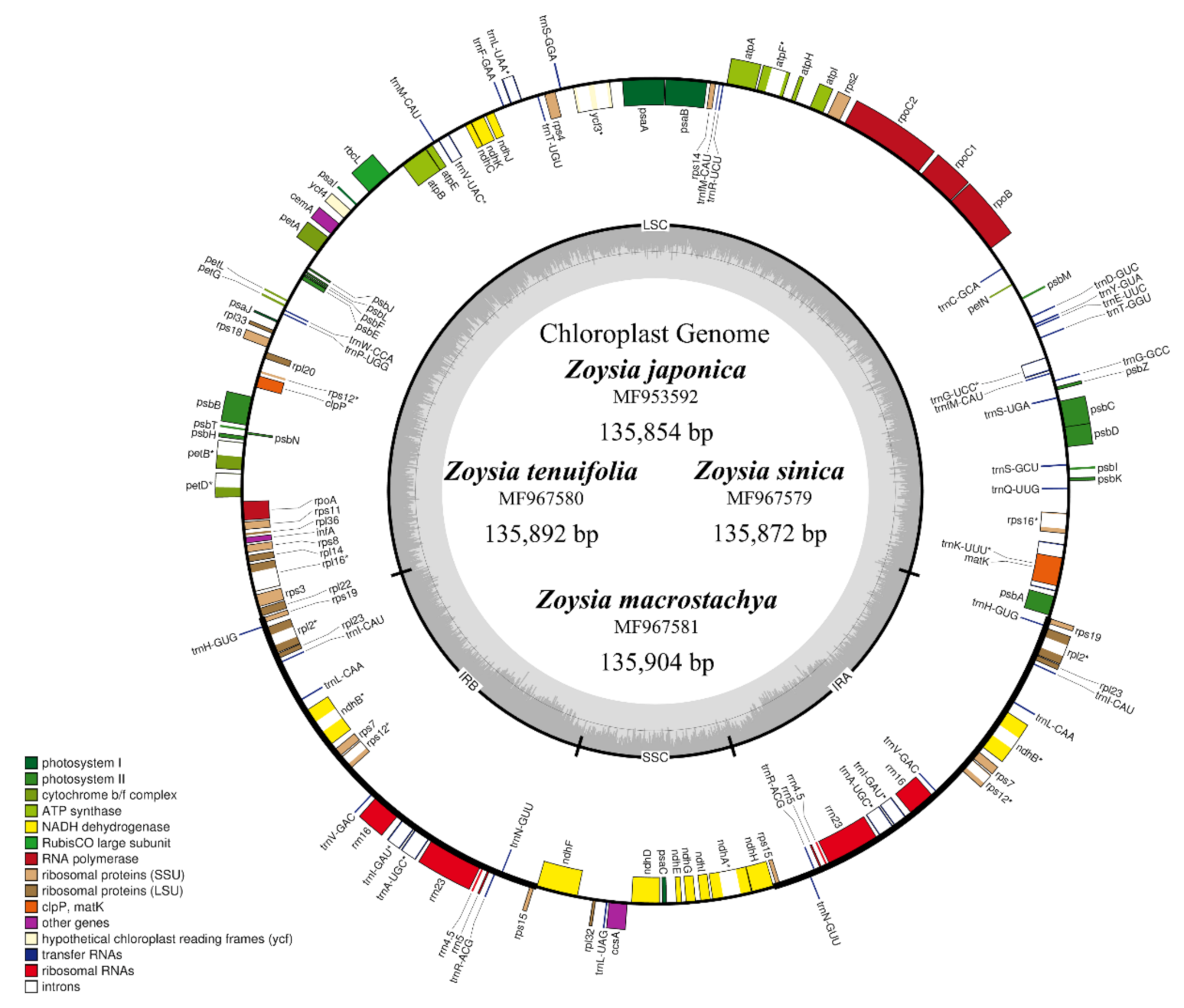
2.1. Chloroplast Genome Structure
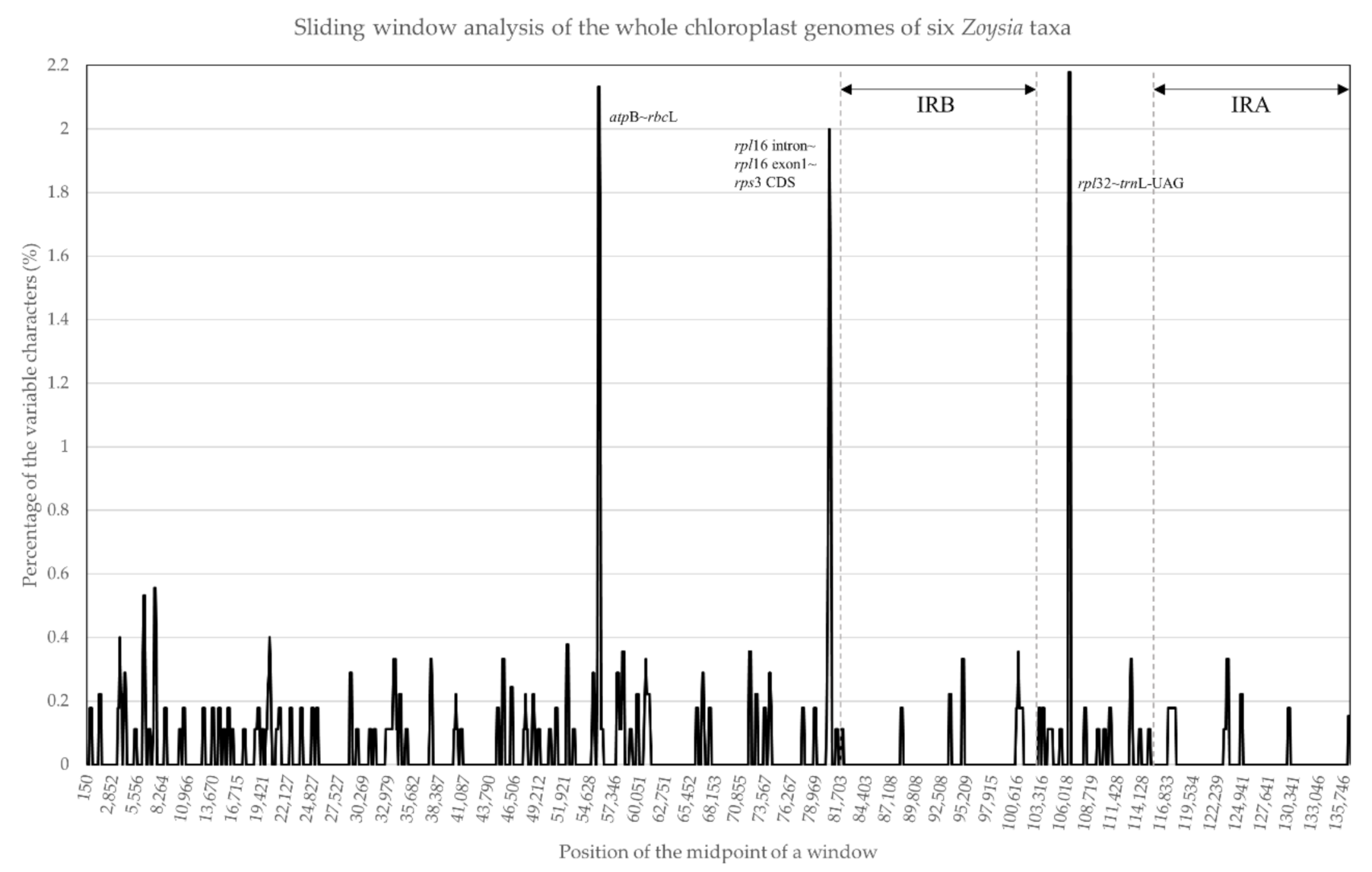
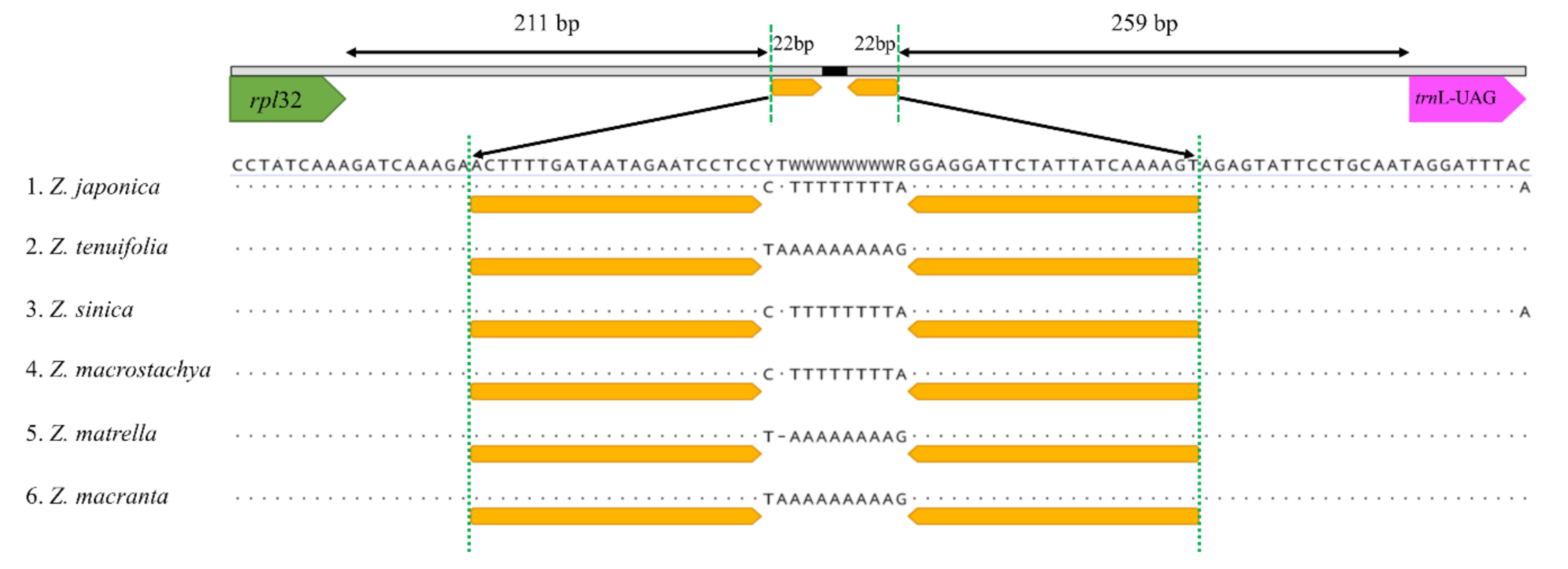
2.2. Sequence Divergence in the Zoysia Chloroplast Genomes
2.3. Simple Sequence Repeats
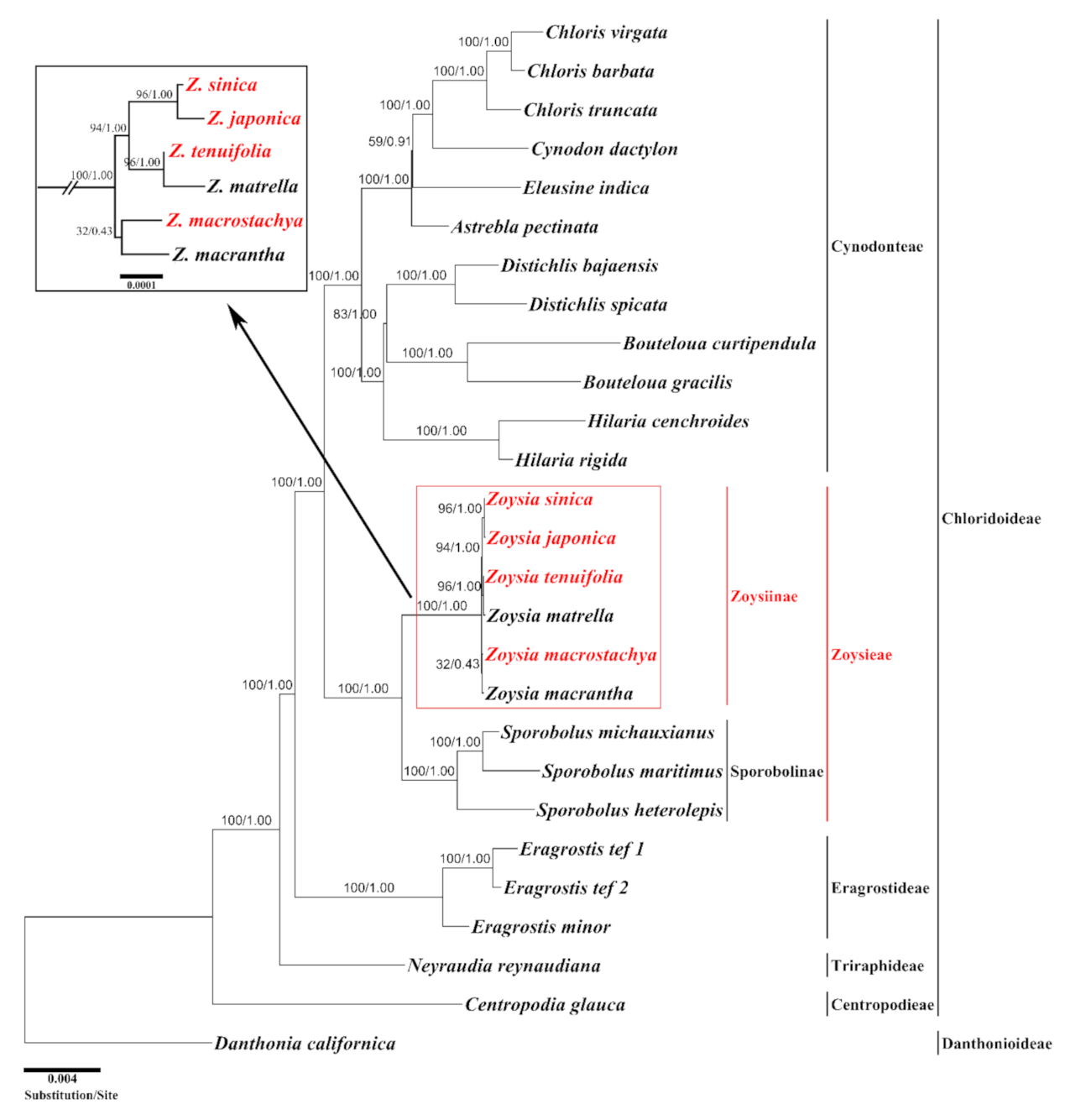
2.4. Phylogenetic Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials, DNA Extraction, and Sequencing
4.2. Chloroplast Genome Assembly and Annotation
4.3. Sequence Analysis
4.4. Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| LSC | Large Single Copy |
| SSC | Small Single Copy |
| IR | Inverted Repeat |
| SSR | Simple Sequence Repeat |
| ML | Maximum Likelihood |
References
- Soreng, R.; Davidse, G.; Peterson, P.; Zuloaga, F.; Judziewicz, E.; Filgueiras, T.; Morrone, O.; Romaschenko, K. A World-Wide Phylogenetic Classification of Poaceae (Gramineae). Available online: http://legacy.tropicos.org/projectwebportal.aspx?pagename=ClassificationNWG&projectid=10 (accessed on 20 September 2020).
- Clayton, W.D.; Renvoize, S.A. Genera graminum. Grasses of the world. Kew Bull. Addit. Ser. 1986, 13, 1–389. [Google Scholar]
- Soreng, R.J.; Peterson, P.M.; Romaschenko, K.; Davidse, G.; Zuloaga, F.O.; Judziewicz, E.J.; Filgueiras, T.S.; Davis, J.I.; Morrone, O. A worldwide phylogenetic classification of the Poaceae (Gramineae). J. Syst. Evol. 2015, 53, 117–137. [Google Scholar] [CrossRef]
- Watson, L.; Dallwitz, M.J. The Grass Genera of the World; CAB international: Wallingford, UK, 1992. [Google Scholar]
- Wu, Z.-Q.; Ge, S. The phylogeny of the BEP clade in grasses revisited: Evidence from the whole-genome sequences of chloroplasts. Mol. Phylogenet. Evol. 2012, 62, 573–578. [Google Scholar] [CrossRef]
- Brown, R. Gramineae. In A Voyage to Terra Australis; Flinders, M., Ed.; W. Bulmer & Company: London, UK, 1814; pp. 5583–5803. [Google Scholar]
- Bouchenak-Khelladi, Y.; Salamin, N.; Savolainen, V.; Forest, F.; van der Bank, M.; Chase, M.W.; Hodkinson, T.R. Large multi-gene phylogenetic trees of the grasses (Poaceae): Progress towards complete tribal and generic level sampling. Mol. Phylogenet. Evol. 2008, 47, 488–505. [Google Scholar] [CrossRef] [PubMed]
- Bouchenak-Khelladi, Y.; Muasya, A.M.; Linder, H.P. A revised evolutionary history of Poales: Origins and diversification. Bot. J. Linn. Soc. 2014, 175, 4–16. [Google Scholar] [CrossRef] [Green Version]
- Grass Phylogeny Working Group II. New grass phylogeny resolves deep evolutionary relationships and discovers C4 origins. New Phytol. 2012, 193, 304–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grass Phylogeny Working Group; Barker, N.P.; Clark, L.G.; Davis, J.I.; Duvall, M.R.; Guala, G.F.; Hsiao, C.; Kellogg, E.A.; Linder, H.P.; Mason-Gamer, R.J. Phylogeny and subfamilial classification of the grasses (Poaceae). Ann. Mo. Bot. Gard. 2001, 88, 373–457. [Google Scholar]
- Peterson, P.M.; Romaschenko, K.; Johnson, G. A classification of the Chloridoideae (Poaceae) based on multi-gene phylogenetic trees. Mol. Phylogenet. Evol. 2010, 55, 580–598. [Google Scholar] [CrossRef]
- Peterson, P.M.; Romaschenko, K.; Arrieta, Y.H.; Saarela, J.M. A molecular phylogeny and new subgeneric classification of Sporobolus (Poaceae: Chloridoideae: Sporobolinae). Taxon 2014, 63, 1212–1243. [Google Scholar] [CrossRef] [Green Version]
- Casler, M.D. Turfgrass Biology, Genetics, and Breeding; John Wiley & Sons: Hoboken, NJ, USA, 2003. [Google Scholar]
- Lee, T.B. Coloured Flora of Korea; Hayangmunsa: Seoul, Korea, 2003. [Google Scholar]
- Tanaka, H.; Hirakawa, H.; Muguerza, M.; Hashiguchi, M.; Tabata, S.; Akashi, R.; Sato, S. The Complete Chloroplast Genome Sequence of Zoysia matrella (L.) Merr. Crop. Sci. 2016, 56, 1206–1212. [Google Scholar] [CrossRef]
- Hajek, T.J., III. Characterization of Microstructural Mutation Events in Plastomes of Chloridoid Grasses (Chloridoideae; Poaceae). Master’s Dissertation, Northern Illinois University, De Kalb, IL, USA, 2016. [Google Scholar]
- Rousseau-Gueutin, M.; Bellot, S.; Martin, G.E.; Boutte, J.; Chelaifa, H.; Lima, O.; Michon-Coudouel, S.; Naquin, D.; Salmon, A.; Ainouche, K.; et al. The chloroplast genome of the hexaploid Spartina maritima (Poaceae, Chloridoideae): Comparative analyses and molecular dating. Mol. Phylogenet. Evol. 2015, 93, 5–16. [Google Scholar] [CrossRef]
- Wambugu, P.W.; Brozynska, M.; Furtado, A.; Waters, D.L.; Henry, R.J. Relationships of wild and domesticated rices (Oryza AA genome species) based upon whole chloroplast genome sequences. Sci. Rep. 2015, 5, 13957. [Google Scholar] [CrossRef] [Green Version]
- Maier, R.M.; Neckermann, K.; Igloi, G.L.; Kössel, H. Complete sequence of the maize chloroplast genome: Gene content, hotspots of divergence and fine tuning of genetic information by transcript editing. J. Mol. Biol. 1995, 251, 614–628. [Google Scholar] [CrossRef]
- Cotton, J.L.; Wysocki, W.P.; Clark, L.G.; Kelchner, S.A.; Pires, J.C.; Edger, P.P.; Mayfield-Jones, D.; Duvall, M.R. Resolving deep relationships of PACMAD grasses: A phylogenomic approach. BMC Plant Biol. 2015, 15, 178. [Google Scholar] [CrossRef] [Green Version]
- Cheon, S.-H.; Jo, S.; Kim, H.-W.; Kim, Y.-K.; Sohn, J.-Y.; Kim, K.-J. The complete plastome sequence of Durian, Durio zibethinus L. (Malvaceae). Mitochondrial DNA B Resour. 2017, 2, 763–764. [Google Scholar] [CrossRef]
- Jo, S.; Kim, H.-W.; Kim, Y.-K.; Cheon, S.-H.; Kim, K.-J. Complete plastome sequence of Averrhoa carambola L. (Oxalidaceae). Mitochondrial DNA B Resour. 2016, 1, 609–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-K.; Kwak, M.H.; Chung, M.G.; Kim, H.-W.; Jo, S.; Sohn, J.-Y.; Cheon, S.-H.; Kim, K.-J. The complete plastome sequence of the endangered orchid Oberonia japonica (Orchidaceae). Mitochondrial DNA B Resour. 2017, 2, 711–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, D.-K.; Kim, K.-J. Complete chloroplast genome sequences of important oilseed crop Sesamum indicum L. PLoS ONE 2012, 7, e35872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guisinger, M.M.; Chumley, T.W.; Kuehl, J.V.; Boore, J.L.; Jansen, R.K. Implications of the plastid genome sequence of Typha (Typhaceae, Poales) for understanding genome evolution in Poaceae. J. Mol. Evol. 2010, 70, 149–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, M.E.; Meyer, G.; Vandergon, T.; Vandergon, V.O. Loss of the acetyl-CoA carboxylase (accD) gene in Poales. Plant Mol. Biol. Report. 2013, 31, 21–31. [Google Scholar] [CrossRef]
- Wysocki, W.P.; Burke, S.V.; Swingley, W.D.; Duvall, M.R. The First Complete Plastid Genome from Joinvilleaceae (J. ascendens; Poales) Shows Unique and Unpredicted Rearrangements. PLoS ONE 2016, 11, e0163218. [Google Scholar]
- Hiratsuka, J.; Shimada, H.; Whittier, R.; Ishibashi, T.; Sakamoto, M.; Mori, M.; Kondo, C.; Honji, Y.; Sun, C.-R.; Meng, B.-Y. The complete sequence of the rice (Oryza sativa) chloroplast genome: Intermolecular recombination between distinct tRNA genes accounts for a major plastid DNA inversion during the evolution of the cereals. Mol. Gen. Genet. 1989, 217, 185–194. [Google Scholar] [CrossRef]
- Huang, Y.-Y.; Cho, S.-T.; Haryono, M.; Kuo, C.-H. Complete chloroplast genome sequence of common bermudagrass (Cynodon dactylon (L.) Pers.) and comparative analysis within the family Poaceae. PLoS ONE 2017, 12, e0179055. [Google Scholar]
- Ogihara, Y.; Isono, K.; Kojima, T.; Endo, A.; Hanaoka, M.; Shiina, T.; Terachi, T.; Utsugi, S.; Murata, M.; Mori, N. Structural features of a wheat plastome as revealed by complete sequencing of chloroplast DNA. Mol. Genet. Genom. 2002, 266, 740–746. [Google Scholar] [CrossRef]
- Jansen, R.K.; Wojciechowski, M.F.; Sanniyasi, E.; Lee, S.-B.; Daniell, H. Complete plastid genome sequence of the chickpea (Cicer arietinum) and the phylogenetic distribution of rps12 and clpP intron losses among legumes (Leguminosae). Mol. Phylogenet. Evol. 2008, 48, 1204–1217. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-L.; Jansen, R.K.; Chumley, T.W.; Kim, K.-J. Gene relocations within chloroplast genomes of Jasminum and Menodora (Oleaceae) are due to multiple, overlapping inversions. Mol. Biol. Evol. 2007, 24, 1161–1180. [Google Scholar] [CrossRef] [Green Version]
- Hupfer, H.; Swiatek, M.; Hornung, S.; Herrmann, R.; Maier, R.; Chiu, W.-L.; Sears, B. Complete nucleotide sequence of the Oenothera elata plastid chromosome, representing plastome I of the five distinguishable Euoenothera plastomes. Mol. Gen. Genet. 2000, 263, 581–585. [Google Scholar] [CrossRef]
- Asano, T.; Tsudzuki, T.; Takahashi, S.; Shimada, H.; Kadowaki, K.-i. Complete nucleotide sequence of the sugarcane (Saccharum officinarum) chloroplast genome: A comparative analysis of four monocot chloroplast genomes. DNA Res. 2004, 11, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Leseberg, C.H.; Duvall, M.R. The complete chloroplast genome of Coix lacryma-jobi and a comparative molecular evolutionary analysis of plastomes in cereals. J. Mol. Evol. 2009, 69, 311–318. [Google Scholar] [CrossRef]
- Saski, C.; Lee, S.-B.; Fjellheim, S.; Guda, C.; Jansen, R.K.; Luo, H.; Tomkins, J.; Rognli, O.A.; Daniell, H.; Clarke, J.L. Complete chloroplast genome sequences of Hordeum vulgare, Sorghum bicolor and Agrostis stolonifera, and comparative analyses with other grass genomes. Theor. Appl. Genet. 2007, 115, 571–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Xia, H.; Cao, M.; Zhang, X.; Zeng, W.; Hu, S.; Tong, W.; Wang, J.; Wang, J.; Yu, J. A comparison of rice chloroplast genomes. Plant Physiol. 2004, 135, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.-H.; Kan, D.-P.; Lee, S.-B.; Daniell, H.; Lee, Y.-W.; Lin, C.-C.; Lin, N.-S.; Lin, C.-S. Complete nucleotide sequence of Dendrocalamus latiflorus and Bambusa oldhamii chloroplast genomes. Tree Physiol. 2009, 29, 847–856. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Hirakawa, H.; Kosugi, S.; Nakayama, S.; Ono, A.; Watanabe, A.; Hashiguchi, M.; Gondo, T.; Ishigaki, G.; Muguerza, M. Sequencing and comparative analyses of the genomes of zoysiagrasses. DNA Res. 2016, 23, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.-J.; Lee, H.-L. Widespread occurrence of small inversions in the chloroplast genomes of land plants. Mol. Cells 2005, 19, 104–113. [Google Scholar]
- Yaneshita, M.; Nagasawa, R.; Engelke, M.C.; Sasakuma, T. Genetic variation and interspecific hybridization among natural populations of zoysiagrasses detected by RFLP analyses of chloroplast and nuclear DNA. Genes Genet. Syst. 1997, 72, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Hashiguchi, M.; Tsuruta, S.; Matsuo, T.; Ebina, M.; Kobayashi, M.; Akamine, H.; Akashi, R. Analysis of genetic resource in Zoysia spp, 2: Evaluation of genetic diversity in zoysiagrass indigenous to southwest islands of Japan based on simple sequence repeat markers. Jpn. J. Grassl. Sci. 2007, 53, 133–137. [Google Scholar]
- Tsuruta, S.I.; Hosaka, F.; Otabara, T.; Hashiguchi, M.; Yamamoto, T.; Akashi, R. Genetic diversity of chloroplast DNA in Zoysia and other warm-season turfgrasses. Grassl. Sci. 2008, 54, 151–159. [Google Scholar] [CrossRef]
- Saarela, J.M.; Burke, S.V.; Wysocki, W.P.; Barrett, M.D.; Clark, L.G.; Craine, J.M.; Peterson, P.M.; Soreng, R.J.; Vorontsova, M.S.; Duvall, M.R. A 250 plastome phylogeny of the grass family (Poaceae): Topological support under different data partitions. PeerJ 2018, 6, e4299. [Google Scholar] [CrossRef] [PubMed]
- Soreng, R.J.; Peterson, P.M.; Romaschenko, K.; Davidse, G.; Teisher, J.K.; Clark, L.G.; Barberá, P.; Gillespie, L.J.; Zuloaga, F.O. A worldwide phylogenetic classification of the Poaceae (Gramineae) II: An update and a comparison of two 2015 classifications. J. Syst. Evol. 2017, 55, 259–290. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.; Doyle, J. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Sambrook, J.; Fritsch, E.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Palmer, J.D. Isolation and structural analysis of chloroplast DNA. Methods Enzymol. 1986, 118, 167–186. [Google Scholar]
- Young, H.A.; Lanzatella, C.L.; Sarath, G.; Tobias, C.M. Chloroplast genome variation in upland and lowland switchgrass. PLoS ONE 2011, 6, e23980. [Google Scholar] [CrossRef] [Green Version]
- Jansen, R.K.; Raubeson, L.A.; Boore, J.L.; Chumley, T.W.; Haberle, R.C.; Wyman, S.K.; Alverson, A.J.; Peery, R.; Herman, S.J.; Fourcade, H.M. Methods for obtaining and analyzing whole chloroplast genome sequences. Methods Enzymol. 2005, 395, 348–384. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef] [PubMed]
- Lohse, M.; Drechsel, O.; Kahlau, S.; Bock, R. OrganellarGenomeDRAW—A suite of tools for generating physical maps of plastid and mitochondrial genomes and visualizing expression data sets. Nucleic Acids Res. 2013, 41, W575–W581. [Google Scholar] [CrossRef]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-Del Barrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Leese, F.; Tollrian, R. Genome-wide analysis of tandem repeats in Daphnia pulex-a comparative approach. BMC Genom. 2010, 11, 277. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: Piscataway, NJ, USA; 2010. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Zoysia japonica | Zoysia tenuifolia | Zoysia sinica | Zoysia macrostachya | Zoysia matrella * | Zoysia macrantha * |
|---|---|---|---|---|---|---|
| Accession number | MF953592 | MF967580 | MF967579 | MF967581 | AP014937 | KT168390 |
| Genome length (bp) | 135,854 | 135,892 | 135,872 | 135,904 | 135,810 | 135,845 |
| LSC length (bp) | 81,348 | 81,376 | 81,366 | 81,392 | 81,308 | 81,352 |
| SSC length (bp) | 12,582 | 12,584 | 12,582 | 12,586 | 12,583 | 12,576 |
| IR length (bp) | 20,962 | 20,966 | 20,962 | 20,963 | 20,960 | 20,959 |
| AT content | 61.6% | 61.6% | 61.6% | 61.6% | 61.6% | 61.6% |
| Total gene contents | 110 | 110 | 110 | 110 | 110 | 110 |
| Genes Category | Group of Genes | Name of Genes |
|---|---|---|
| Self-replication | rRNA genes | rrn16 (x2), rrn23 (x2), rrn4.5 (x2), rrn5 (x2) |
| tRNA genes | 30 trn genes (8 are in IR regions) | |
| Small subunit of ribosome | rps2, rps3, rps4, rps7 (x2), rps8, rps11, rps12 * (x2), rps14, rps15 (x2), rps16 *, rps18, rps19 (x2) | |
| Large subunit of ribosome | rpl2 * (x2), rpl14, rpl16 *, rpl20, rpl22, rpl23 (x2), rpl32, rpl33, rpl36 | |
| DNA dependent RNA polymerase | rpoA, rpoB, rpoC1, rpoC2 | |
| Genes for photosynthesis | Subunits of NADH-dehydrogenase | ndhA *, ndhB * (x2), ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK |
| Subunits of photosystem 1 | psaA, psaB, psaC, psaI, psaJ | |
| Subunits of photosystem 2 | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ | |
| Subunits of cytochrome b/f complex | petA, petB *, petD *, petG, petL, petN | |
| Subunits of ATP synthase | atpA, atpB, atpE, atpF *, atpH, atpI | |
| Large subunit of rubisco | rbcL | |
| Other genes | Maturase | matK |
| Protease | clpP | |
| Envelope membrane protein | cemA | |
| c-type cytochrome synthesis gene | ccsA | |
| Translational initiation factor | infA | |
| Genes of unknown functions Open Reading Frames (ORF) | ycf3 **, ycf4 | |
| Unit | Length (bp) | Zoysia japonica | Zoysia sinica | Zoysia macrostachya | Zoysia tenuifolia |
|---|---|---|---|---|---|
| No. of SSRs | No. of SSRs | No. of SSRs | No. of SSRs | ||
| A | 17 | 0 | 1 | 0 | 0 |
| 16 | 2 | 1 | 1 | 3 | |
| 15 | 1 | 0 | 0 | 1 | |
| 14 | 2 | 3 | 0 | 1 | |
| 13 | 2 | 2 | 3 | 3 | |
| 12 | 3 | 3 | 6 | 3 | |
| 11 | 4 | 4 | 6 | 6 | |
| 10 | 10 | 9 | 7 | 9 | |
| C | 11 | 1 | 1 | 0 | 0 |
| 10 | 0 | 0 | 0 | 1 | |
| G | 11 | 0 | 0 | 0 | 1 |
| 10 | 1 | 1 | 2 | 0 | |
| T | 22 | 1 | 0 | 0 | 1 |
| 17 | 0 | 4 | 0 | 0 | |
| 16 | 0 | 0 | 1 | 0 | |
| 14 | 0 | 0 | 2 | 1 | |
| 13 | 3 | 4 | 4 | 4 | |
| 12 | 7 | 7 | 5 | 7 | |
| 11 | 5 | 6 | 4 | 7 | |
| 10 | 8 | 8 | 12 | 6 | |
| AT | 12 | 1 | 1 | 1 | 1 |
| 10 | 3 | 3 | 3 | 3 | |
| GA | 10 | 1 | 1 | 1 | 1 |
| TA | 14 | 1 | 1 | 1 | 1 |
| 10 | 1 | 1 | 1 | 1 | |
| TC | 10 | 1 | 1 | 1 | 1 |
| AAT | 12 | 1 | 1 | 1 | 1 |
| Total | 59 | 63 | 62 | 63 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheon, S.-H.; Woo, M.-A.; Jo, S.; Kim, Y.-K.; Kim, K.-J. The Chloroplast Phylogenomics and Systematics of Zoysia (Poaceae). Plants 2021, 10, 1517. https://doi.org/10.3390/plants10081517
Cheon S-H, Woo M-A, Jo S, Kim Y-K, Kim K-J. The Chloroplast Phylogenomics and Systematics of Zoysia (Poaceae). Plants. 2021; 10(8):1517. https://doi.org/10.3390/plants10081517
Chicago/Turabian StyleCheon, Se-Hwan, Min-Ah Woo, Sangjin Jo, Young-Kee Kim, and Ki-Joong Kim. 2021. "The Chloroplast Phylogenomics and Systematics of Zoysia (Poaceae)" Plants 10, no. 8: 1517. https://doi.org/10.3390/plants10081517
APA StyleCheon, S. -H., Woo, M. -A., Jo, S., Kim, Y. -K., & Kim, K. -J. (2021). The Chloroplast Phylogenomics and Systematics of Zoysia (Poaceae). Plants, 10(8), 1517. https://doi.org/10.3390/plants10081517