
Response of Normal and Low-Phytate Genotypes of Pea (Pisum sativum L.) on Phosphorus Foliar Fertilization

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. The Effect of Foliar P Fertilization on Content of Chlorophyll and Chlorophyll Fluorescence Parameters
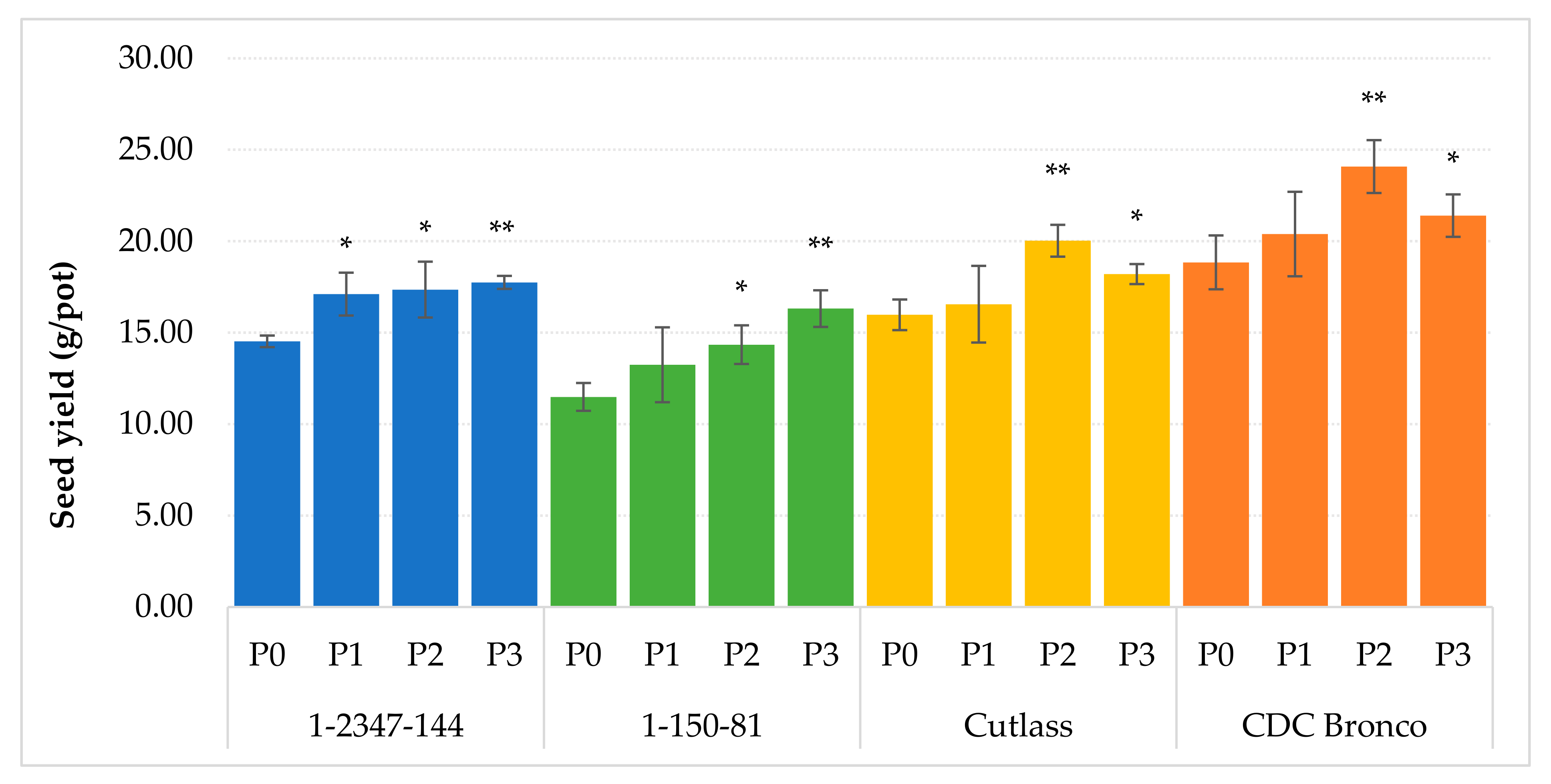
2.2. The Effect of Foliar P Fertilization, on Yield Parameters of Pea
2.3. The Effect of Foliar P Fertilization, on Pea Seed Quality
3. Materials and Methods
3.1. Plant Materials, Plant Cultivation and Conditions of Growth
3.2. Measurement of Photosynthetic Parameters of Pea Plants
3.2.1. Content of Chlorophyll (N-Tester Value)
3.2.2. Chlorophyll Fluorescence Parameters
3.3. Yield Parameters and Seed Quality
3.3.1. Determination of Phytic Acid (Phytate)
3.3.2. Analysis of Crude Protein and Ash
3.3.3. Determination of Nutrient Contents
3.4. Statistical Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kosev, V.; Pachev, I. Genetic Improvement of field pea (Pisum sativum L.) in Bulgaria. Field Veg. Crop Res. 2010, 47, 403–408. [Google Scholar]
- Tesfaye, M.; Liu, J.; Allan, D.L.; Vance, C.P. Genomic and genetic control of phosphate stress in legumes. Plant Physiol. 2007, 144, 594–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotaru, V.; Sinclair, T.R. Influence of plant phosphorus and iron concentrations on growth of soybean. J. Plant Nutr. 2009, 32, 1513–1526. [Google Scholar] [CrossRef]
- Mitran, T.; Meena, R.S.; Lal, R.; Layek, J.; Kumar, S.; Datta, R. Role of Soil Phosphorus on Legume Production. In Legumes for Soil Health and Sustainable Management; Meena, R.S., Das, A., Yadav, G.S., Lal, R., Eds.; Springer: Singapore; New York, NY, USA, 2018; pp. 487–510. [Google Scholar]
- Ejaz, S.; Batool, S.; Anjum, M.A.; Naz, S.; Qayyum, M.F.; Naqqash, T.; Shah, K.H.; Ali, S. Effects of inoculation of root-associative Azospirillum and Agrobacterium strains on growth, yield and quality of pea (Pisum sativum L.) grown under different nitrogen and phosphorus regimes. Sci. Hortic. 2020, 270, 109401. [Google Scholar] [CrossRef]
- Agegnehu, G.; Fessehaie, R. Response of faba bean to phosphate fertilizer and weed control on Nitisols of Ethiopian highlands. Ital. J. Agron. 2006, 1, 281–290. [Google Scholar] [CrossRef]
- Khadraji, A.; Bouhadi, M.; Ghoulam, C. Effect of Soil Available Phosphorus Levels on Chickpea (Cicer arietinum L.)—Rhizobia Symbiotic Association. Legum. Res. 2020, 43, 878–883. [Google Scholar]
- Alene, A.A.; Raffi, M.M.; Tiruneh, K.J. Phosphorus use efficiency, yield and nodulation of mung bean (Vigna radiata L.) as influenced by the rate of phosphorus and Rhizobium strains inoculation in Metema district, Ethiopia. J. Plant Nutr. 2021, 44, 1300–1315. [Google Scholar] [CrossRef]
- Bi, Y.; Zhou, P.; Li, S.; Wei, Y.; Xiong, X.; Shi, Y.; Liu, N.; Zhang, Y. Interspecific interactions contribute to higher forage yield and are affected by phosphorus application in a fully-mixed perennial legume and grass intercropping system. Field Crops Res. 2019, 244, 107636. [Google Scholar] [CrossRef]
- Qader, H.R. Effect of foliar application of Phosphorus on Growth and development of Vicia faba L. under Magnesium levels. ZANCO J. Pure Appl. Sci. 2019, 31, 173–179. [Google Scholar]
- Froese, S.; Wiens, J.T.; Warkentin, T.; Schoenau, J.J.; Beres, B. Response of canola, wheat, and pea to foliar phosphorus fertilization at a phosphorus-deficient site in eastern Saskatchewan. Can. J. Plant Sci. 2020, 100, 642–652. [Google Scholar] [CrossRef]
- Gibson, R.S.; Yeudall, F.; Drost, N.; Mtitimuni, B.; Cullinan, T. Dietary interventions to prevent zinc deficiency. Am. J. Clin. Nutr. 1998, 68, 484S–487S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnlund, J.R.; King, J.C.; Gong, B.; Keyes, W.R.; Michel, M.C. A stable isotope study of copper absorption in young men: Effect of phytate and a-cellulose. Am. J. Clin. Nutr. 1985, 42, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Raboy, V. Myo-Inositol-1,2,3,4,5,6-hexakisphosphate. Phytochemistry 2003, 64, 1033–1043. [Google Scholar] [CrossRef]
- Raboy, V. Approaches and challenges to engineering seed phytate and total phosphorus. Plant Sci. 2009, 177, 281–296. [Google Scholar] [CrossRef]
- Wilcox, J.R.; Premachandra, G.S.; Young, K.A.; Raboy, V. Isolation of high seed inorganic P, low-phytate soybean mutants. Crop. Sci. 2000, 40, 1601–1605. [Google Scholar] [CrossRef] [Green Version]
- Warkentin, T.D.; Delgerjav, O.; Arganosa, G.; Rehman, A.U.; Bett, K.E.; Anbessa, Y.; Rossnagel, B.; Raboy, V. Development and Characterization of Low-Phytate Pea. Crop. Sci. 2012, 52, 74–78. [Google Scholar] [CrossRef]
- Raboy, V.; Dickinson, D.B. Phytic acid levels in seeds of Glycine max and G. soja as influenced by phosphorus status. Crop. Sci. 1993, 33, 1300–1305. [Google Scholar] [CrossRef]
- Miller, G.A.; Youngs, V.L.; Oplinger, E.S. Effect of available soil-phosphorus and environment on the phytic acid concentration in oats. Cereal Chem. 1980, 57, 192–194. [Google Scholar]
- Saneoka, H.; Koba, T. Plant growth and phytic acid accumulation in seed as affected by phosphorus application in maize (Zea mays L.). Grassl. Sci. 2003, 48, 485–489. [Google Scholar]
- Fernández, V.; Brown, P.H. From plant surface to plant metabolism: The uncertain fate of foliar-applied nutrients. Front. Plant Sci. 2013, 4, 289. [Google Scholar] [CrossRef] [Green Version]
- Noack, S.R.; McBeath, T.M.; McLaughlin, M.J. Potential for foliar phosphorus fertilization of dryland cereal crops: A review. Crop. Pasture Sci. 2011, 62, 659–669. [Google Scholar] [CrossRef]
- Ling, F.; Silberbush, M. Response of maize to foliar vs. soil application of nitrogen-phosphorus-potassium fertilizers. J. Plant Nutr. 2007, 1, 2333–2342. [Google Scholar] [CrossRef]
- Mosali, J.; Desta, K.; Teal, R.; Freeman, K.; Martin, K.; Lawless, J.; Raun, W. Effect of foliar application of phosphorus on winter wheat grain yield, phosphorus uptake and use efficiency. J. Plant Nutr. 2006, 29, 2147–2163. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, Z.; Ahmad, R.; Saifullah; Ashraf, M.Y. Foliar Applied Phosphorous Enhanced Growth, Chlorophyll Contents, Gas Exchange Attributes and PUE in Wheat (Triticum aestivum L.). J. Plant Nutr. 2015, 38, 1929–1943. [Google Scholar] [CrossRef]
- Thapar, S.; Sekhon, B.S.; Atwal, A.; Singh, R. Phosphorus assimilation in mycorrhizal moong (Vigna radita L.) plants under different phosphorus levels. Plant Sci. 1990, 71, 209–214. [Google Scholar] [CrossRef]
- Lopez-Cantarero, I.; Lorente, F.A.; Romero, L. Are chlorophylls good indicators of nitrogen and phosphorus levels? J. Plant Nutr. 1994, 17, 979–990. [Google Scholar] [CrossRef]
- Fan, L.; Silberbush, M. Response of maize to foliar vs. soil application of nitrogen, phosphorus and potassium fertilizers. J. Plant Nutr. 2002, 25, 2333–2342. [Google Scholar]
- Shubhra, J.D.; Goswami, C.L.; Munjal, R. Influence of phosphorus application on water relations, biochemical parameters and gum content in cluster bean under water deficit. Biol. Plant. 2004, 48, 445–448. [Google Scholar] [CrossRef]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: Cambridge, MA, USA, 2012; p. 672. [Google Scholar]
- Carstensen, A.; Herdean, A.; Schmidt, S.B.; Sharma, A.; Spetea, C.; Pribil, M.; Husted, S. The impacts of phosphorus deficiency on the photosynthetic electron transport chain. Plant Physiol. 2018, 177, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.X.; Weng, X.Y.; Yang, Y. Effect of phosphorus deficiency on the photosynthetic characteristics of rice plants. Russ. J. Plant Physiol. 2007, 54, 741–748. [Google Scholar] [CrossRef]
- Xing, D.; Wu, Y. Effect of phosphorus deficiency on photosynthetic inorganic carbon assimilation of three climber plant species. Bot. Stud. 2014, 55, 60. [Google Scholar] [CrossRef] [Green Version]
- Taliman, N.A.; Dong, Q.; Echigo, K.; Raboy, V.; Saneoka, H. Effect of Phosphorus Fertilization on the Growth, Photosynthesis, Nitrogen Fixation, Mineral Accumulation, Seed Yield, and Seed Quality of a Soybean Low-Phytate Line. Plants 2019, 8, 119. [Google Scholar] [CrossRef] [Green Version]
- Fryer, M.J.; Andrews, J.R.; Oxborough, K.; Blowers, D.A.; Baker, N.R. Relationship between CO2 assimilation, photosynthetic electron transport, and active O2 metabolism in leaves of maize in the field during periods of low temperature. Plant Physiol. 1998, 116, 571–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thuynsma, R.; Kleinert, A.; Kossmann, J.; Valentine, A.J.; Hills, P.N. The effects of limiting phosphate on photosynthesis and growth of Lotus japonicus. S. Afr. J. Bot. 2016, 104, 244–248. [Google Scholar] [CrossRef]
- Shunmugam, A.S.K.; Bock, C.; Arganosa, G.C.; Georges, F.; Gray, G.R.; Warkentin, T.D. Accumulation of Phosphorus-Containing Compounds in Developing Seeds of Low-Phytate Pea (Pisum sativum L.) Mutants. Plants 2015, 4, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.J.; Muhammad, D.; Fahad, S.; Adnan, M.; Wahid, F.; Alamri, S.; Khan, F.; Dawar, K.M.; Irshad, I.; Danish, S.; et al. Phosphorus Nutrient Management through Synchronization of Application Methods and Rates in Wheat and Maize Crops. Plants 2020, 9, 1389. [Google Scholar]
- Rady, M.M.; El-Shewy, A.A.; Seif El-Yazal, M.A.; Abdelaal, K.E.S. Response of Salt-Stressed Common Bean Plant Performances to Foliar Application of Phosphorus (MAP). Int. Lett. Nat. Sci. 2018, 72, 7–20. [Google Scholar] [CrossRef]
- Singh, U.; Singh, B. Effect of basal and foliar application of diammonium phosphate in cognizance with phosphate-solubilizing bacteria on growth, yield and quality of rainfed chickpea (Cicer arietinum). Indian J. Agron. 2014, 59, 427–432. [Google Scholar]
- Krasilnikoff, G.; Gahoonia, T.; Erik-Nelson, N. Variation in phosphorus uptake by genotypes of cowpea (Vigna unguiculata L. Walp) due to differences in root and root hair length and induced rhizosphere processes. Plant Soil 2003, 251, 83–91. [Google Scholar]
- Bohn, L.; Meyer, A.S.; Rasmussen, S.K. Phytate: Impact on environment and human nutrition. A challenge for molecular breeding. J. Zhejiang Univ. Sci. B 2008, 9, 165–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimek-Kopyra, A.; Hakl, J.; Skladanka, J.; Dłużniewska, J. Influence of phosphorus nutrition on seed yield and quality of pea (Pisum sativum L.) cultivars across different seasons. Ital. J. Agron. 2019, 14, 208–213. [Google Scholar] [CrossRef] [Green Version]
- Musa, E.M.; Elsheikh, E.A.E.; Ahmed, I.A.M.; Babiker, E.E. Effect of intercropping, Bradyrhizobium inoculation and N, P fertilizers on yields, physical and chemical quality of cowpea seeds. Front. Agric. China 2011, 5, 543–557. [Google Scholar] [CrossRef]
- Kyei-Boahen, S.; Savala, C.E.N.; Chikoye, D.; Abaidoo, R. Growth and Yield Responses of Cowpea to Inoculation and Phosphorus Fertilization in Different Environments. Front. Plant Sci. 2017, 8, 646. [Google Scholar] [CrossRef] [Green Version]
- Malakondaiah, N.; Rajeswararao, G. Effect of foliar application of phosphorus on growth and mineral composition in peanut plants (Arachis hypogaea L.) under salt-stress. Plant Soil 1979, 52, 41–48. [Google Scholar] [CrossRef]
- Shankarlingappa, B.C.; Shivraj, B.; Vishwanath, K.P. Interaction effect of phosphorus and sulphur on uptake of nitrogen, phosphorus, potassium and sulphur by pigeonpea (Cajanus cajan). Indian J. Agron. 2000, 45, 348–352. [Google Scholar]
- Kumar, P.; Kumar, P.; Singh, T.; Singh, A.K.; Yadav, R.I. Effect of different potassium levels on mungbean under custard apple based Agri-Horti system. Afr. J. Agric. Res. 2014, 9, 728–734. [Google Scholar]
- Kumar, P.; Pandey, S.K.; Kumar, P. Effect of Different Phosphorus Levels on Nutrient Content, Uptake and Economics of Urd bean under Custard Apple based Agri-Horti System. J. AgriSearch 2015, 2, 88–93. [Google Scholar]
- Zhang, Y.Q.; Deng, Y.; Chen, R.Y.; Cui, Z.L.; Chen, X.P.; Yost, R.; Zhang, F.S.; Zou, C.Q. The reduction in zinc concentration of wheat grain upon increased phosphorus-fertilization and its mitigation by foliar zinc application. Plant Soil 2012, 361, 143–152. [Google Scholar] [CrossRef]
- Ova, E.A.; Kutman, U.B.; Ozturk, L.; Cakmak, I. High phosphorus supply reduced zinc concentration of wheat in native soil but not in autoclaved soil or nutrient solution. Plant Soil 2015, 393, 147–162. [Google Scholar] [CrossRef]
- Warkentin, T.; Vandenberg, A.; Banniza, S.; Slinkard, A. CDC Bronco field pea. Can. J. Plant Sci. 2005, 85, 649–650. [Google Scholar] [CrossRef] [Green Version]
- Blade, S.; Warkentin, T.; Vandenberg, A. Cutlass field pea. Can. J. Plant Sci. 2004, 84, 533–534. [Google Scholar]
- Zbíral, J. Analysis of Soils, I. Unified Techniques, 2nd ed.; Central Institute for Supervising and Testing in Agriculture: Brno, Czech Republic, 2002; p. 197. (In Czech) [Google Scholar]
- Netto, A.L.; Campostrini, E.; Goncalves de Oliverira, J.; Bressan-Smith, R.E. Photosynthetic pigments, nitrogen, chlorophyll a fluorescence and SPAD-502 readings in coffee leaves. Sci. Hortic. 2005, 104, 199–209. [Google Scholar] [CrossRef]
- Škarpa, P.; Klofáč, D.; Krčma, F.; Šimečková, J.; Kozáková, Z. Effect of Plasma Activated Water Foliar Application on Selected Growth Parameters of Maize (Zea mays L.). Water 2020, 12, 3545. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C.; Knapp, M. How to correctly determine the different chlorophyll fluorescence parameters and the chlorophyll fluorescence decrease ratio RFd of leaves with the PAM fluorometer. Photosynthetica 2005, 43, 379–393. [Google Scholar] [CrossRef]
- McKie, V.A.; McCleary, B.V. A Novel and Rapid Colorimetric Method for Measuring Total Phosphorus and Phytic Acid in Food and Animal Feeds. J. AOAC Int. 2016, 99, 738–743. [Google Scholar] [CrossRef] [Green Version]
- Mlejnkova, V.; Horky, P.; Kominkova, M.; Skladanka, J.; Hodulikova, L.; Adam, V.; Mlcek, J.; Jurikova, T.; Sochor, J. Biogenic amines and hygienic quality of lucerne silage. Open Life Sci. 2016, 11, 280–286. [Google Scholar] [CrossRef] [Green Version]
- Zbíral, J. Plant Analysis: Integrated Work Procedures; Central Institute for Supervising and Testing in Agriculture: Brno, Czech Republic, 2005; p. 192. (In Czech) [Google Scholar]
- StatSoft, Inc. STATISTICA (Data Analysis Software System), Version 12. 2013. Available online: www.statsoft.com (accessed on 5 May 2021).



{kind=link}
{kind=link}
| Genotype | Treatment | N-Tester Value | ΦPSII | RFd | |||
|---|---|---|---|---|---|---|---|
| T1 | T2 | T1 | T2 | T1 | T2 | ||
| 1-2347-144 | P0 | 448 ± 6 | 378 ± 5 | 0.801 ± 0.011 | 0.809 ± 0.014 | 2.04 ± 0.03 | 1.94 ± 0.04 |
| P1 | 458 ± 5 | 390 ± 4 * | 0.820 ± 0.008 * | 0.825 ± 0.010 | 2.13 ± 0.02 | 2.06 ± 0.05 * | |
| P2 | 464 ± 7 ** | 401 ± 4 ** | 0.818 ± 0.007 * | 0.831 ± 0.004 | 2.42 ± 0.20 ** | 2.07 ± 0.04 * | |
| P3 | 463 ± 6 * | 402 ± 3 ** | 0.822 ± 0.002 * | 0.833 ± 0.007 * | 2.42 ± 0.20 ** | 2.13 ± 0.07 ** | |
| 1-150-81 | P0 | 443 ± 10 | 371 ± 3 | 0.792 ± 0.015 | 0.803 ± 0.004 | 1.99 ± 0.02 | 1.93 ± 0.07 |
| P1 | 456 ± 10 * | 383 ± 7 * | 0.816 ± 0.004 ** | 0.813 ± 0.005 | 2.08 ± 0.04 | 2.00 ± 0.07 | |
| P2 | 464 ± 7 ** | 373 ± 9 | 0.816 ± 0.008 ** | 0.837 ± 0.005 ** | 2.11 ± 0.02 | 2.08 ± 0.06 * | |
| P3 | 475 ± 13 ** | 401 ± 7 ** | 0.829 ± 0.005 ** | 0.840 ± 0.007 ** | 2.16 ± 0.04 * | 2.10 ± 0.04 ** | |
| Cutlass | P0 | 439 ± 6 | 368 ± 8 | 0.803 ± 0.011 | 0.792 ± 0.018 | 2.06 ± 0.04 | 1.77 ± 0.09 |
| P1 | 452 ± 5 * | 374 ± 6 | 0.823 ± 0.008 * | 0.792 ± 0.009 | 2.02 ± 0.03 | 1.82 ± 0.07 | |
| P2 | 458 ± 5 ** | 402 ± 3 ** | 0.829 ± 0.005 ** | 0.815 ± 0.008 | 2.09 ± 0.06 | 1.97 ± 0.06 ** | |
| P3 | 458 ± 6 ** | 381 ± 5 * | 0.818 ± 0.019 | 0.789 ± 0.003 | 2.09 ± 0.02 | 1.76 ± 0.05 | |
| CDC Bronco | P0 | 447 ± 5 | 386 ± 8 | 0.804 ± 0.015 | 0.812 ± 0.043 | 2.00 ± 0.07 | 1.91 ± 0.04 |
| P1 | 463 ± 5 * | 394 ± 12 | 0.828 ± 0.004 ** | 0.827 ± 0.006 | 2.05 ± 0.10 | 1.94 ± 0.07 | |
| P2 | 465 ± 8 ** | 408 ± 4 ** | 0.825 ± 0.007 * | 0.816 ± 0.004 | 2.22 ± 0.08 ** | 2.02 ± 0.08 | |
| P3 | 468 ± 6 ** | 398 ± 6 * | 0.820 ± 0.011 * | 0.777 ± 0.008 ** | 2.07 ± 0.07 | 1.86 ± 0.13 | |
| Genotype | Treatment | Plant Height (cm) | Seed Weight (g/1000 Seeds) | Seed Number (Seed/Pot) |
|---|---|---|---|---|
| 1-2347-144 | P0 | 63 ± 5 | 193 ± 10 | 75.3 ± 3.8 |
| P1 | 66 ± 9 | 209 ± 20 | 81.7 ± 0.6 ** | |
| P2 | 72 ± 5 * | 220 ± 29 | 79.0 ± 2.0 | |
| P3 | 75 ± 8 ** | 212 ± 16 | 84.0 ± 1.0 ** | |
| 1-150-81 | P0 | 64 ± 7 | 185 ± 10 | 62.3 ± 1.5 |
| P1 | 69 ± 7 | 197 ± 30 | 67.3 ± 2.5 * | |
| P2 | 73 ± 17 * | 207 ± 26 | 69.3 ± 3.1 ** | |
| P3 | 76 ± 9 ** | 229 ± 22 ** | 71.3 ± 1.5 ** | |
| Cutlass | P0 | 72 ± 2 | 188 ± 20 | 85.0 ± 4.4 |
| P1 | 75 ± 7 | 217 ± 7 | 76.3 ± 1.5 ** | |
| P2 | 73 ± 4 | 212 ± 16 | 94.3 ± 4.5 ** | |
| P3 | 71 ± 3 | 181 ± 16 | 100.7 ± 2.3 ** | |
| CDC Bronco | P0 | 98 ± 18 | 234 ± 16 | 80.3 ± 1.5 |
| P1 | 109 ± 11 ** | 238 ± 9 | 86.0 ± 3.6 * | |
| P2 | 111 ± 9 ** | 240 ± 10 | 100.3 ± 2.5 ** | |
| P3 | 102 ± 9 | 226 ± 25 | 94.7 ± 1.5 ** |
| Genotype | Treatment | P (% DM) | Phytate (g/100 g DM) | Crude Protein (% DM) |
|---|---|---|---|---|
| 1-2347-144 | P0 | 0.36 ± 0.02 | 0.82 ± 0.07 | 22.2 ± 0.4 |
| P1 | 0.35 ± 0.02 | 0.82 ± 0.04 | 23.4 ± 0.7 | |
| P2 | 0.38 ± 0.06 | 0.90 ± 0.19 | 22.6 ± 0.4 | |
| P3 | 0.41 ± 0.08 | 1.00 ± 0.27 | 23.3 ± 2.0 | |
| 1-150-81 | P0 | 0.42 ± 0.02 | 0.82 ± 0.22 | 23.7 ± 0.8 |
| P1 | 0.42 ± 0.04 | 0.66 ± 0.07 | 22.4 ± 2.0 | |
| P2 | 0.39 ± 0.01 | 0.62 ± 0.04 | 20.9 ± 1.6 ** | |
| P3 | 0.45 ± 0.02 | 0.75 ± 0.19 | 21.9 ± 0.4 | |
| Cutlass | P0 | 0.30 ± 0.01 | 0.69 ± 0.05 | 20.1 ± 0.5 |
| P1 | 0.30 ± 0.02 | 0.69 ± 0.06 | 21.1 ± 0.3 | |
| P2 | 0.39 ± 0.03 ** | 1.01 ± 0.10 ** | 22.7 ± 1.1 ** | |
| P3 | 0.43 ± 0.03 ** | 1.13 ± 0.08 ** | 24.1 ± 1.2 ** | |
| CDC Bronco | P0 | 0.41 ± 0.03 | 1.04 ± 0.10 | 20.0 ± 0.2 |
| P1 | 0.41 ± 0.03 | 0.98 ± 0.07 | 20.4 ± 1.1 | |
| P2 | 0.41 ± 0.04 | 1.08 ± 0.07 | 20.7 ± 0.9 | |
| P3 | 0.46 ± 0.03 | 1.14 ± 0.10 | 20.7 ± 0.8 |
| Genotype | Treatment | Ash (% DM) | K (% DM) | Mg (% DM) | Ca (% DM) | Zn (mg/kg DM) |
|---|---|---|---|---|---|---|
| 1-2347-144 | P0 | 2.89 ± 0.08 | 0.92 ± 0.02 | 0.125 ± 0.005 | 0.036 ± 0.005 | 34.0 ± 5.3 |
| P1 | 2.84 ± 0.06 | 0.85 ± 0.07 | 0.118 ± 0.011 | 0.041 ± 0.003 | 29.8 ± 3.2 | |
| P2 | 2.96 ± 0.17 | 0.92 ± 0.05 | 0.121 ± 0.005 | 0.033 ± 0.003 | 25.8 ± 2.3 | |
| P3 | 3.14 ± 0.22 * | 1.01 ± 0.09 * | 0.125 ± 0.009 | 0.030 ± 0.004 | 26.0 ± 2.3 | |
| 1-150-81 | P0 | 3.26 ± 0.13 | 1.02 ± 0.04 | 0.121 ± 0.008 | 0.040 ± 0.007 | 28.7 ± 0.6 |
| P1 | 3.13 ± 0.29 | 0.99 ± 0.09 | 0.114 ± 0.010 | 0.035 ± 0.003 | 28.5 ± 1.2 | |
| P2 | 2.97 ± 0.06 * | 0.95 ± 0.00 | 0.112 ± 0.006 | 0.032 ± 0.007 * | 26.6 ± 5.8 * | |
| P3 | 3.09 ± 0.04 | 1.01 ± 0.00 | 0.121 ± 0.004 | 0.030 ± 0.004 ** | 25.6 ± 1.2 * | |
| Cutlass | P0 | 2.68 ± 0.06 | 0.82 ± 0.06 | 0.109 ± 0.002 | 0.032 ± 0.002 | 24.7 ± 2.4 |
| P1 | 2.68 ± 0.07 | 0.83 ± 0.01 | 0.114 ± 0.009 | 0.033 ± 0.005 | 25.6 ± 4.0 | |
| P2 | 2.96 ± 0.06 * | 0.90 ± 0.03 | 0.116 ± 0.002 | 0.035 ± 0.005 | 25.6 ± 3.7 | |
| P3 | 3.02 ± 0.18 ** | 0.87 ± 0.08 | 0.129 ± 0.007 ** | 0.045 ± 0.006 ** | 25.2 ± 1.6 | |
| CDC Bronco | P0 | 2.96 ± 0.14 | 0.93 ± 0.07 | 0.104 ± 0.004 | 0.030 ± 0.002 | 34.9 ± 3.7 |
| P1 | 2.96 ± 0.08 | 0.96 ± 0.03 | 0.111 ± 0.004 | 0.029 ± 0.002 | 34.4 ± 4.1 | |
| P2 | 3.02 ± 0.11 | 0.96 ± 0.04 | 0.116 ± 0.007 * | 0.029 ± 0.002 | 27.1 ± 2.4 * | |
| P3 | 3.08 ± 0.01 | 1.01 ± 0.01 | 0.108 ± 0.003 | 0.031 ± 0.005 | 29.1 ± 4.2 |
| Soil Parameter | Value |
|---|---|
| pH (CaCl2) | 6.09 |
| Soil oxidizable carbon (Cox) | 0.80% |
| Clay | 20% |
| Silt | 27% |
| Sand | 53% |
| CEC (Cation Exchange Capacity) | 164 mmol/kg |
| total N | 0.19% |
| Ammonium N (NH4+) | 1.48 mg/kg |
| Nitrate N (NO3−) | 17.2 mg/kg |
| Available P (Mehlich III) | 36.4 mg/kg * |
| Available K (Mehlich III) | 400 mg/kg |
| Available Ca (Mehlich III) | 2 720 mg/kg |
| Available Mg (Mehlich III) | 214 mg/kg |
| Chlorophyll Fluorescence Parameters | Pulse Type | Light Intensity (μmol/m2/s) | Phase | Duration (s) | 1st Pulse (s) | Pulse Interval (s) |
|---|---|---|---|---|---|---|
| ΦPSII | Saturation | 2400 | - | 1 pulse | ||
| RFd | Flash | 900 | L | 60 | 0.2 | 1 |
| DR | 88 | 1 | 1 | |||
| Saturation | 2400 | L | 60 | 7 | 12 | |
| DR | 88 | 11 | 26 | |||
| Actinic | 300 | L | 60 | - | - |
| Chlorophyll Fluorescence Parameters | Ref. | |
|---|---|---|
| ΦPSII | Fm − F0/Fm | [57] |
| RFd | Fd/Fs | [58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Škarpa, P.; Školníková, M.; Antošovský, J.; Horký, P.; Smýkalová, I.; Horáček, J.; Dostálová, R.; Kozáková, Z. Response of Normal and Low-Phytate Genotypes of Pea (Pisum sativum L.) on Phosphorus Foliar Fertilization. Plants 2021, 10, 1608. https://doi.org/10.3390/plants10081608
Škarpa P, Školníková M, Antošovský J, Horký P, Smýkalová I, Horáček J, Dostálová R, Kozáková Z. Response of Normal and Low-Phytate Genotypes of Pea (Pisum sativum L.) on Phosphorus Foliar Fertilization. Plants. 2021; 10(8):1608. https://doi.org/10.3390/plants10081608
Chicago/Turabian StyleŠkarpa, Petr, Marie Školníková, Jiří Antošovský, Pavel Horký, Iva Smýkalová, Jiří Horáček, Radmila Dostálová, and Zdenka Kozáková. 2021. "Response of Normal and Low-Phytate Genotypes of Pea (Pisum sativum L.) on Phosphorus Foliar Fertilization" Plants 10, no. 8: 1608. https://doi.org/10.3390/plants10081608
APA StyleŠkarpa, P., Školníková, M., Antošovský, J., Horký, P., Smýkalová, I., Horáček, J., Dostálová, R., & Kozáková, Z. (2021). Response of Normal and Low-Phytate Genotypes of Pea (Pisum sativum L.) on Phosphorus Foliar Fertilization. Plants, 10(8), 1608. https://doi.org/10.3390/plants10081608