
Effects of Epichloë sinensis Endophyte and Host Ecotype on Physiology of Festuca sinensis under Different Soil Moisture Conditions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
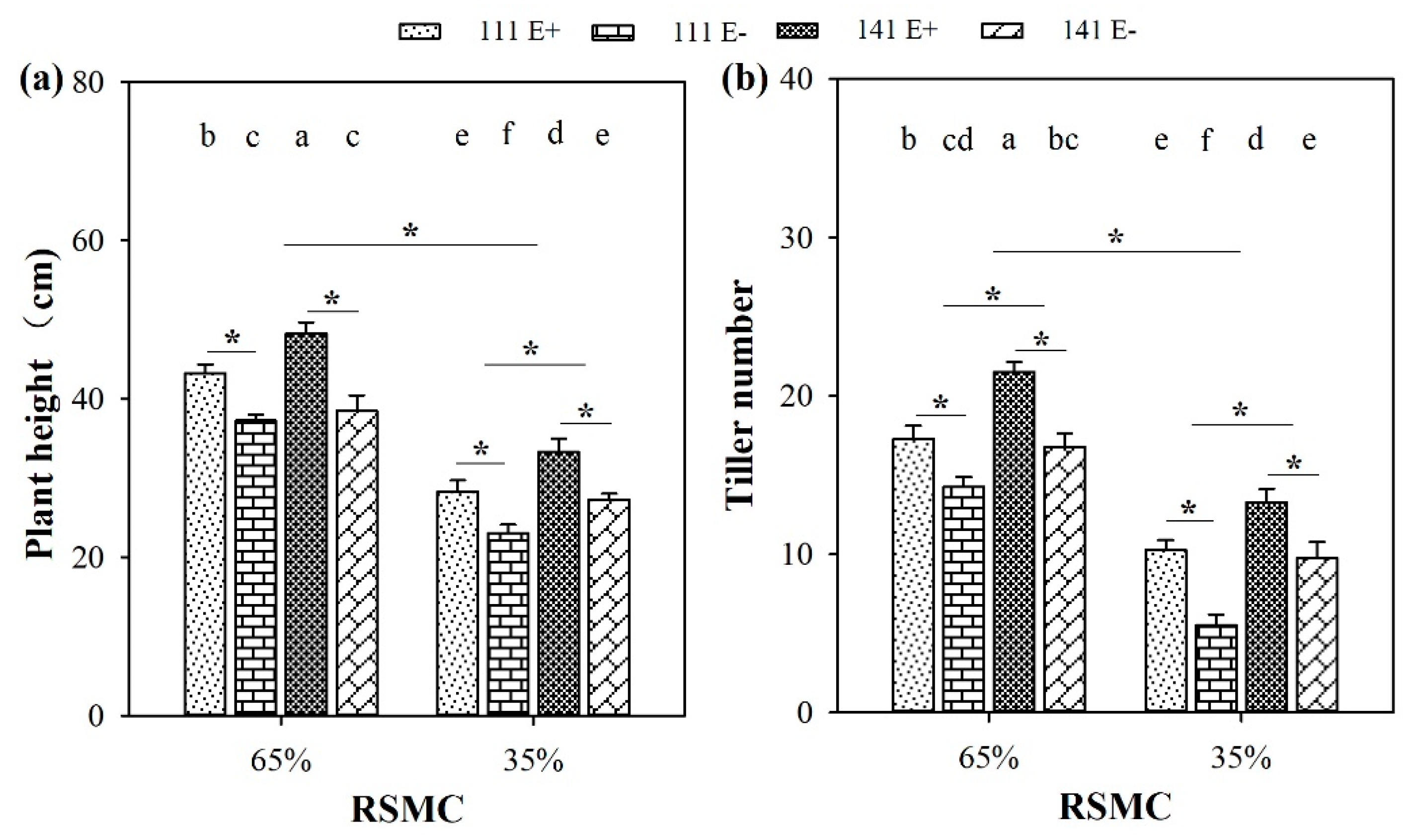
2.1. Plant Height and Tiller Number
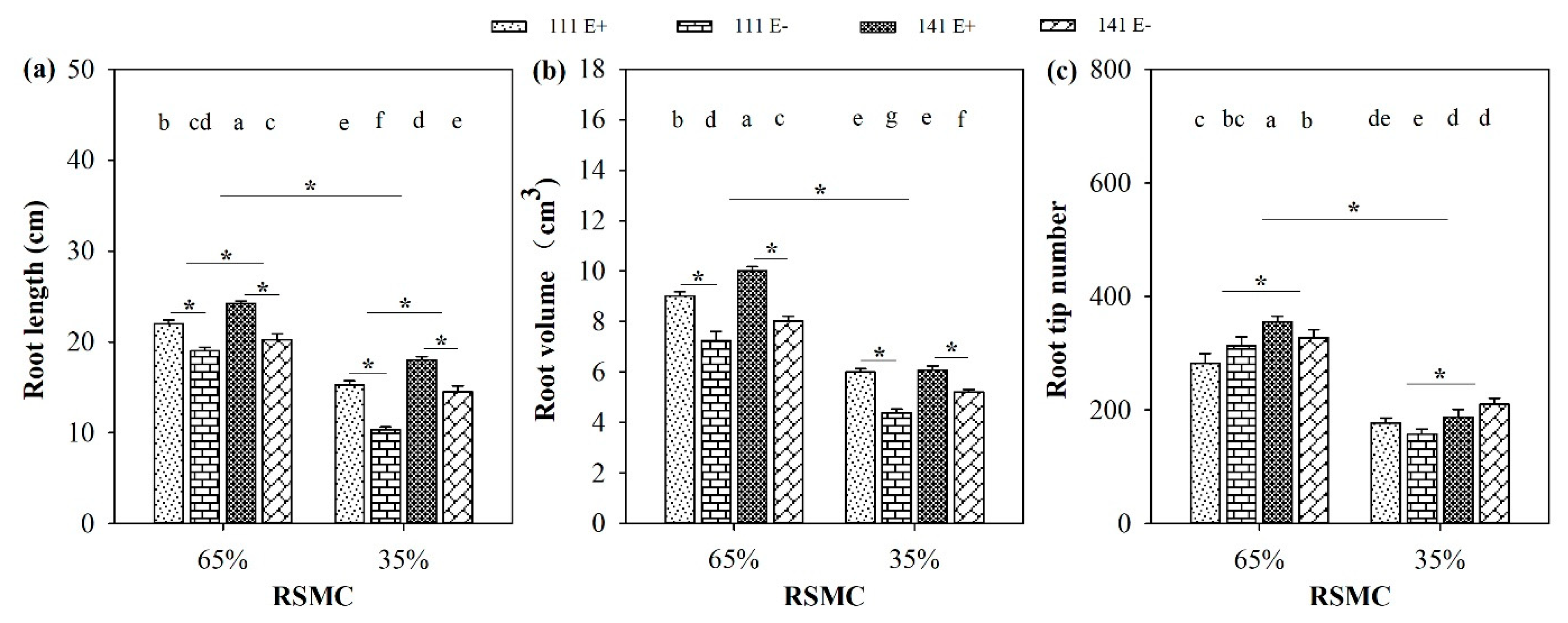
2.2. Root Indices
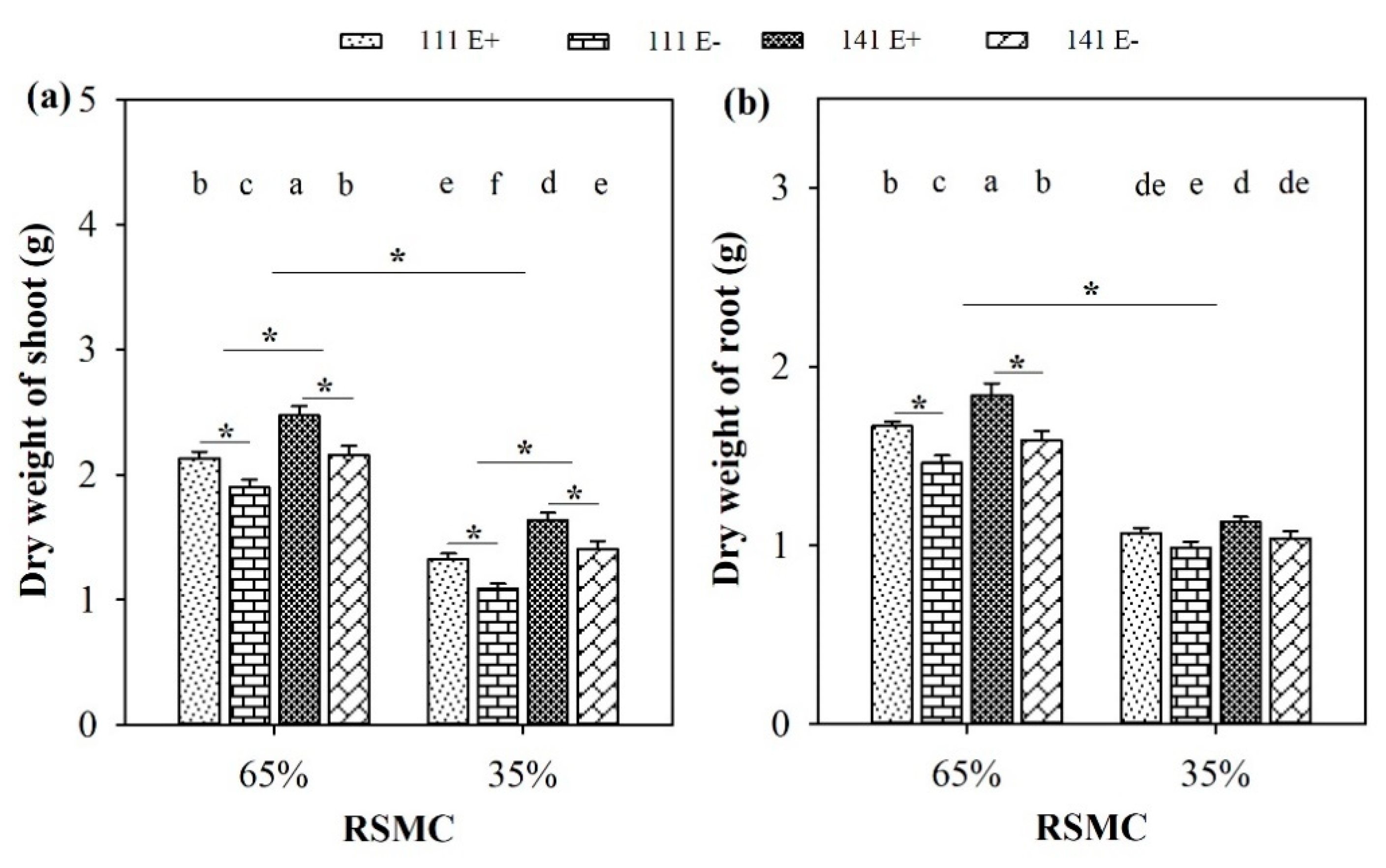
2.3. Biomass of Shoot and Root
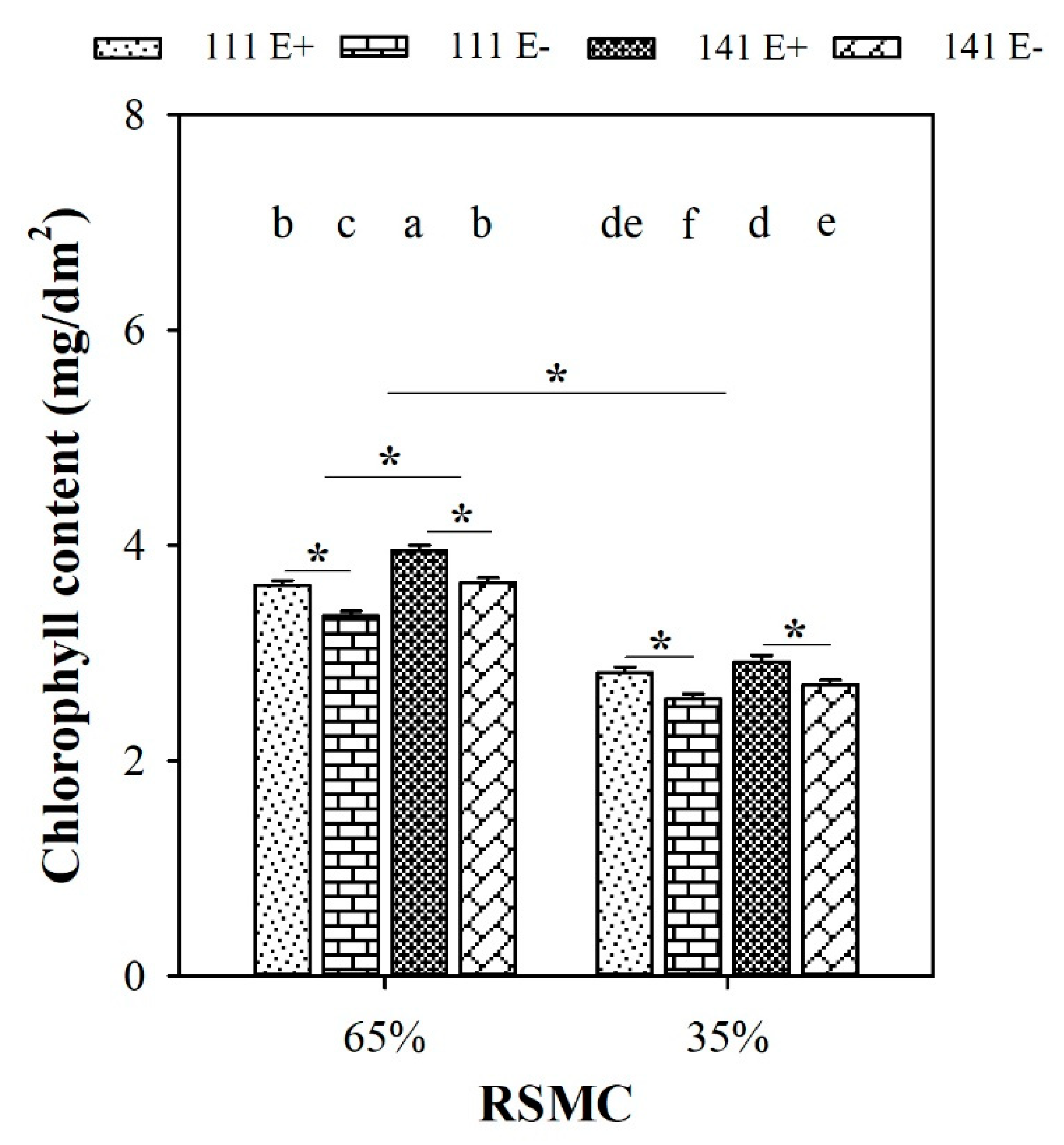
2.4. Chlorophyll Content
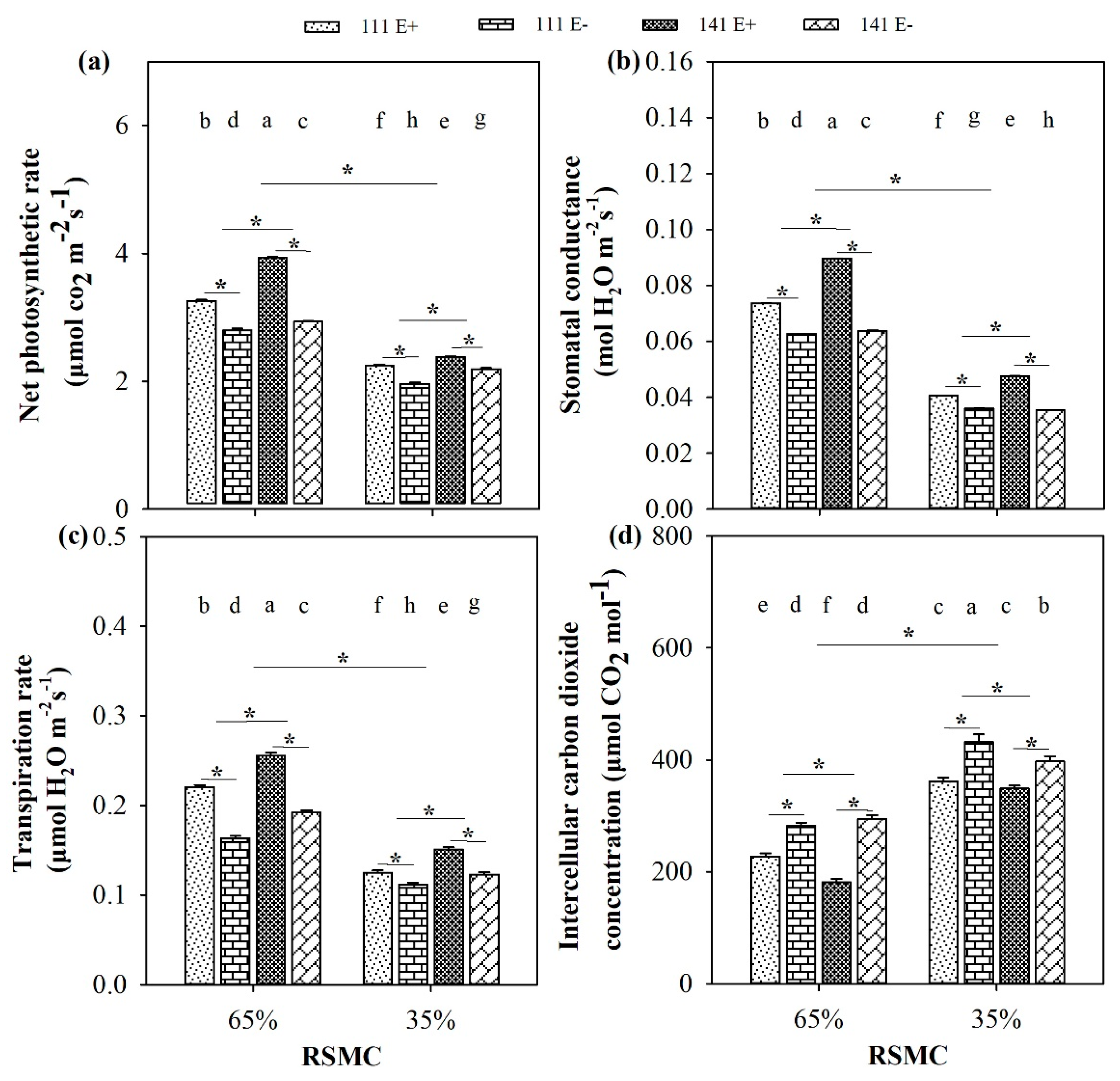
2.5. Photosynthetic Indices
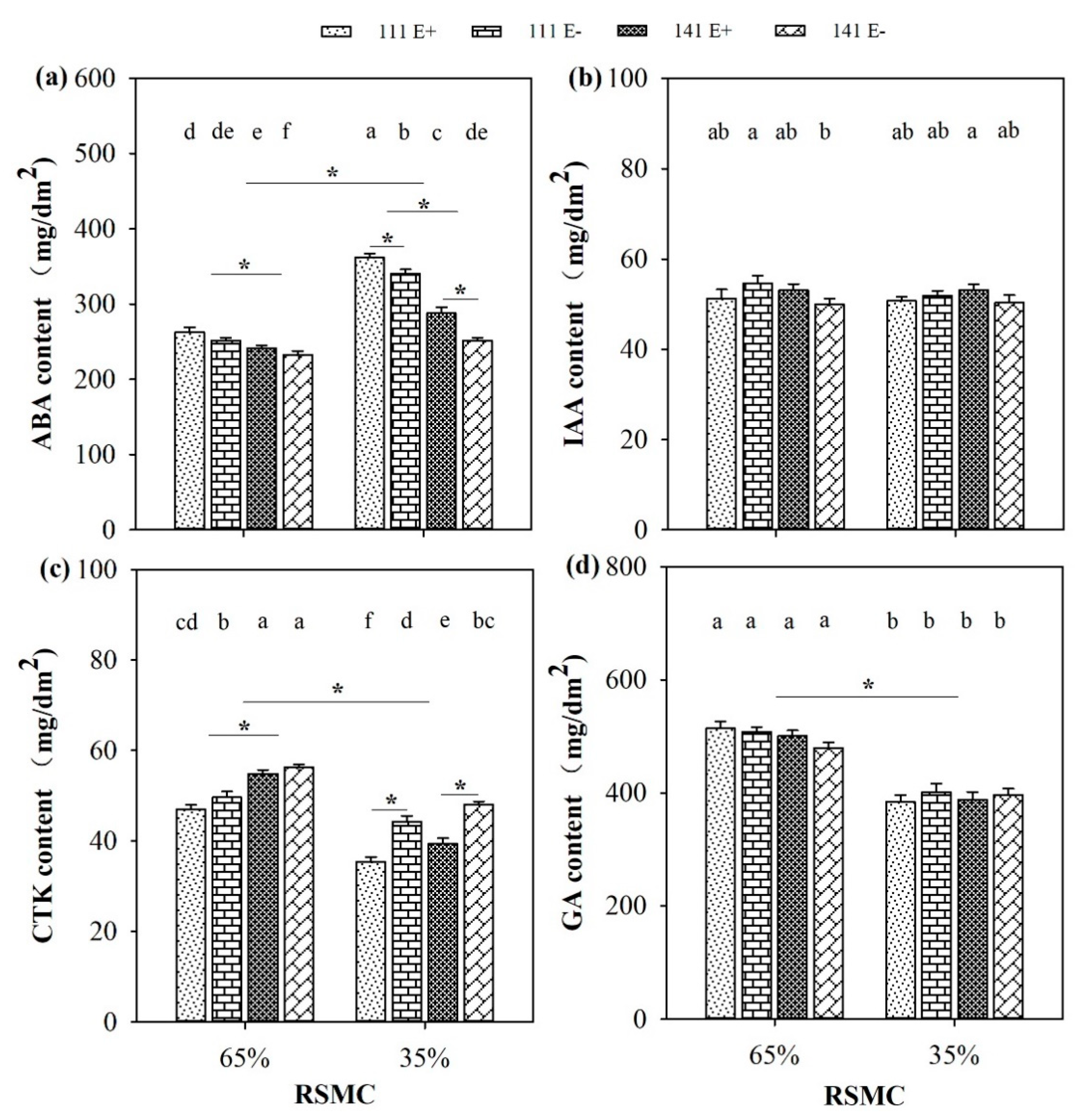
2.6. Phytohormones
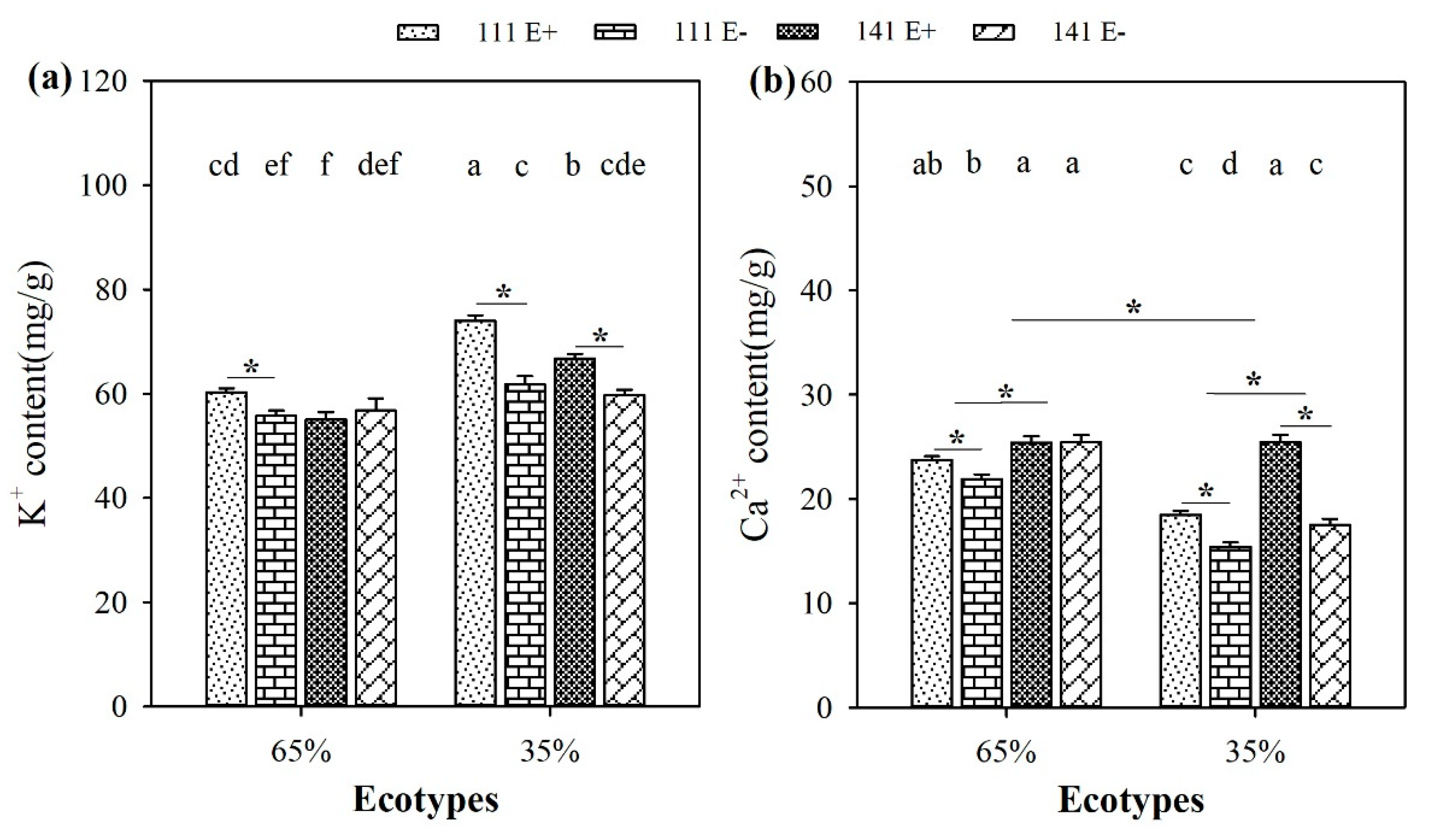
2.7. Contents of K+ and Ca2+
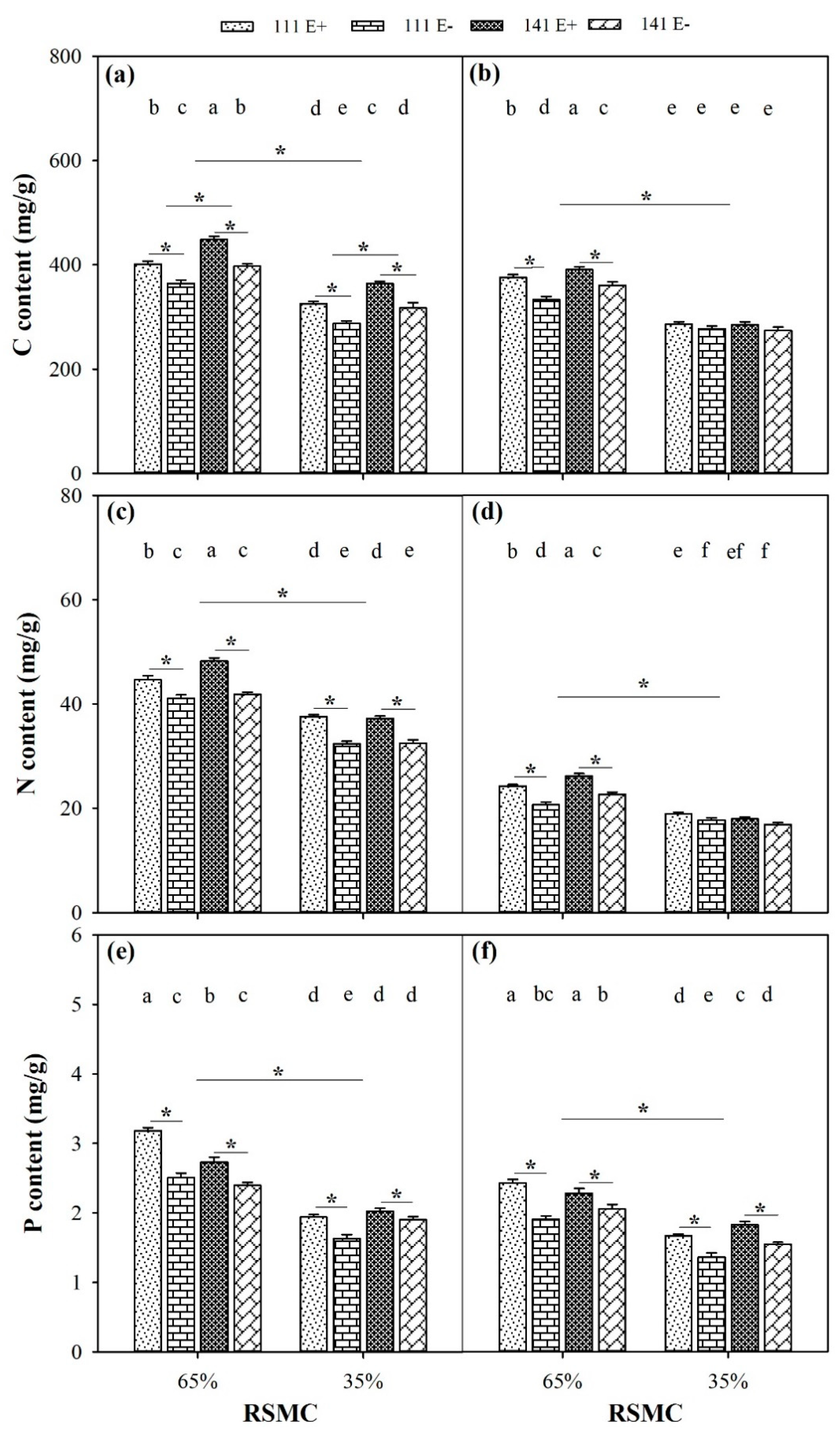
2.8. Contents of C, N, and P
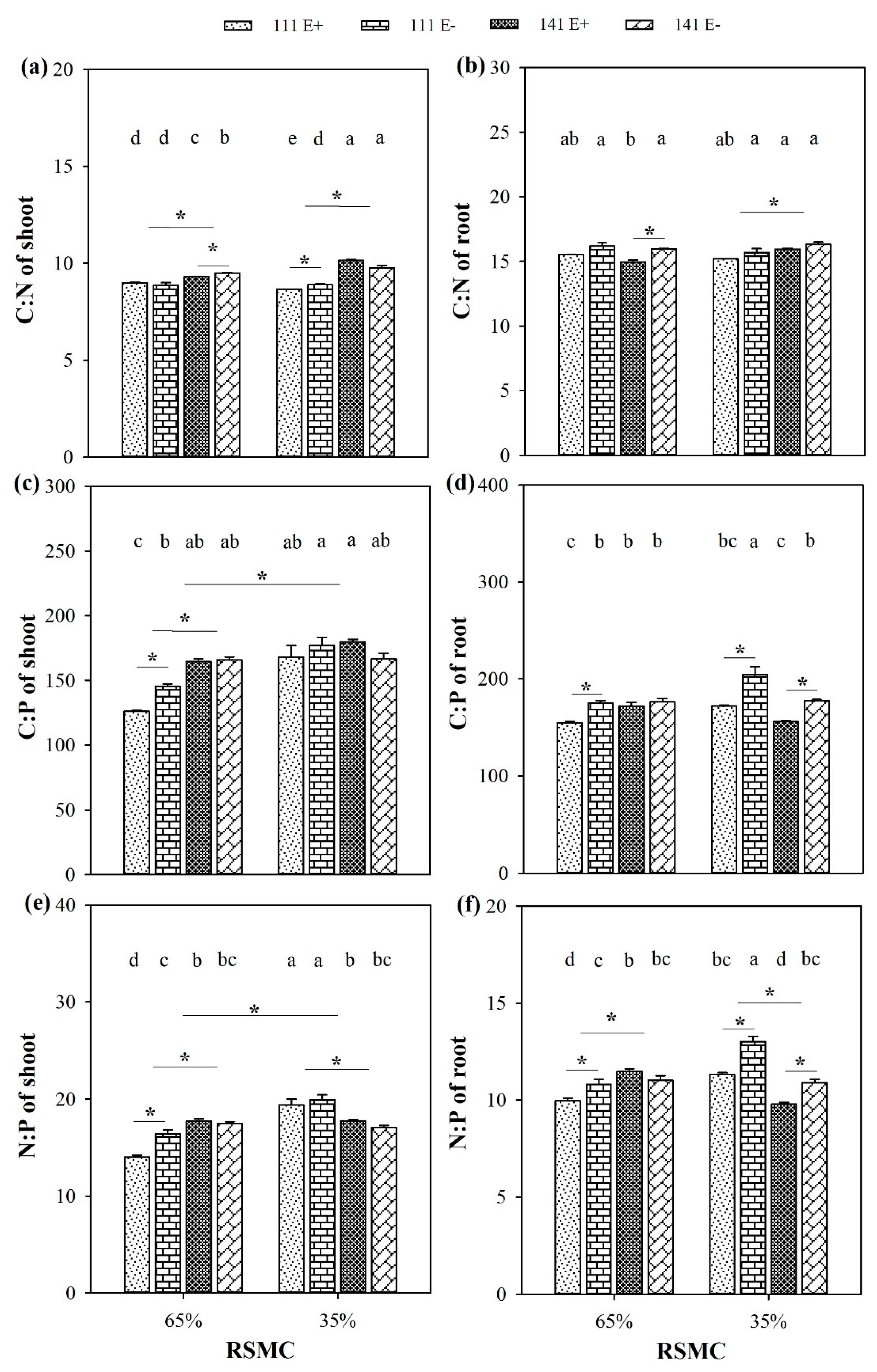
2.9. Ecological Stoichiometry
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Experimental Design
4.3. Measurement Protocols
4.3.1. Plant Performance
4.3.2. Chlorophyll Content
4.3.3. Photosynthetic Indices
4.3.4. Phytohormones
4.3.5. Potassium (K+) and Calcium (Ca2+) Ion Contents
4.3.6. C, N, and P Contents
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Bray, E.A.; Bailey-Serres, J.; Weretilnyk, E. Responses to abiotic stresses. In Biochemistry and Molecular Biology of Plants; American Society of Plant Physiologists: Rockville, MD, USA, 2000; pp. 1158–1249. [Google Scholar]
- Arraudeau, M.A. Breeding strategies for drought resistance. In Drought Resistance in Cereals; Baker, F.W.C., Ed.; CAB International: Wallingford, UK, 1989; pp. 107–116. [Google Scholar]
- Hosseini, F.; Mosaddeghi, M.R.; Hajabbasi, M.A.; Sabzalian, M.R. Role of fungal endophyte of tall fescue (Epichloë coenophiala) on water availability, wilting point and integral energy in texturally-different soils. Agric. Water Manag. 2016, 163, 197–211. [Google Scholar] [CrossRef]
- Karcher, D.E.; Richardson, M.D.; Hignight, K.; Rush, D. Drought tolerance of tall fescue populations selected for high root/shoot ratios and summer survival. Crop. Sci. 2008, 48, 771–777. [Google Scholar] [CrossRef]
- Clarke, J.M.; Durley, R.C. The responses of plants to drought stress. In Water Stress on Plants; Simpson, G.M., Ed.; Praeger Publishers: New York, NY, USA, 1981; pp. 89–139. [Google Scholar]
- Malinowski, D.P.; Belesky, D.P. Adaptations of endophytE−infected cool-season grasses to environmental stresses: Mechanisms of drought and mineral stress tolerance. Crop. Sci. 2000, 40, 923. [Google Scholar] [CrossRef]
- Leuchtmann, A.; Bacon, C.W.; Schardl, C.L.; White, J.F.; Tadych, M. Nomenclatural realignment of Neotyphodium speices with genus Epichloë. Mycologia 2014, 106, 202–215. [Google Scholar] [CrossRef] [PubMed]
- Schardl, C.L.; Leuchtmann, A.; Spiering, M.J. Symbioses of grasses with seedborne fungal endophytes. Annu. Rev. Plant Biol. 2004, 55, 315–340. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.B.; Krauss, J. Symbiosis between grasses and asexual fungal endophytes. Curr. Opin. Plant Biol. 2005, 8, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Hahn, H.; Mcmanus, M.T.; Warnstorff, K.; Monahan, B.J.; Young, C.A.; Davies, E. Neotyphodium fungal endophytes confer physiological protection to perennial ryegrass (lolium perenne L.) subjected to a water deficit. Environ. Exp. Bot. 2008, 63, 183–199. [Google Scholar] [CrossRef]
- Clay, K.; Schardl, C. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. Am. Nat. 2002, 160 (Suppl. S4), S99–S127. [Google Scholar] [CrossRef]
- Song, M.L.; Chai, Q.; Li, X.Z.; Yao, X.; Li, C.J.; Christensen, M.; Nan, Z.B. An asexual epichlo endophyte modifies the nutrient stoichiometry of wild barley (hordeum brevisubulatum) under salt stress. Plant Soil 2015, 387, 153–165. [Google Scholar] [CrossRef]
- Redman, R.S. Thermotolerance Generated by Plant/Fungal Symbiosis. Science 2002, 298, 1581. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.X.; Li, C.J.; Nan, Z.B. Effects of cadmium stress on growth and anti-oxidative systems in Achnatherum inebrians symbiotic with Neotyphodium gansuense. J. Hazard. Mater. 2010, 175, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, L.M.; Popay, A.J.; Finch, S.C.; Clearwater, M.J.; Cave, V.M. Temperature and plant genotype alter alkaloid concentrations in ryegrass infected with an Epichloë endophyte and this affects an insect herbivore. Front. Plant Sci. 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Song, M.L.; Li, X.Z.; Saikkonen, K.; Li, C.J.; Nan, Z.B. An asexual Epichloë endophyte enhances waterlogging tolerance of hordeum brevisubulatum. Fungal Ecol. 2015, 13, 44–52. [Google Scholar] [CrossRef]
- Kuldau, G.; Bacon, C. Clavicipitaceous endophytes: Their ability to enhance resistance of grasses to multiple stresses. Biol. Control 2008, 46, 57–71. [Google Scholar] [CrossRef]
- Lin, W.H.; Kuang, Y.; Wang, J.J.; Duan, D.D.; Xu, W.B.; Ma, B.H.; Tian, P. Effects of seasonal variation on the alkaloids of different ecotypes of Epichloë endophytE−Festuca sinensis associations. Front. Microbiol. 2019, 10, 1695. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.J.; Lin, W.H.; Zhao, Y.T.; Meng, C.; Ma, A.W.; Xue, L.H.; Kuang, Y.; Tian, P. Physiological and biochemical responses of Festuca sinensis seedlings to temperature and soil moisture stress. Funct. Plant Biol. 2017, 44, 1007. [Google Scholar] [CrossRef]
- Zhou, L.Y.; Li, C.J.; Zhang, X.X.; Johnson, R.; Bao, G.S.; Yao, X.; Chai, Q. Effects of cold shocked Epichloë infected Festuca sinensis on ergot alkaloid accumulation. Fungal Ecol. 2015, 14, 99–104. [Google Scholar] [CrossRef]
- Song, H.; Nan, Z.B.; Song, Q.Y.; Xia, C.; Li, X.Z.; Yao, X.; Xu, W.B.; Tian, P. Advances in research on Epichloë endophytes in chinese native grasses. Front. Microbiol. 2016, 7, 1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.Y.; Zhang, X.X.; Li, C.J.; Christensen, M.J.; Nan, Z.B. Antifungal activity and phytochemical investigation of the asexual endophyte of Epichloë sp. from Festuca sinensis. Sci. China Life Sci. 2015, 58, 821–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.Y.; Zhong, S.; Duo, H.M.; Qiao, F. Antimicrobial activity and composition of volatile substance of Epichloë sp. endophyte isolated from Festuca sinensis. Nat. Prod. Res. Dev. 2019, 31, 1543–1551. [Google Scholar]
- Peng, Q.Q.; Li, C.J.; Song, M.L.; Nan, Z.B. Effects of seed hydropriming on growth of Festuca sinensis infected with Neotyphodium endophyte. Fungal Ecol. 2013, 6, 83–91. [Google Scholar] [CrossRef]
- Tian, P.; Xu, W.B.; Li, C.J.; Song, H.; Nan, Z.B. Phylogenetic relationship and taxonomy of a hybrid epichlo species symbiotic with Festuca sinensis. Mycol. Prog. 2020, 19, 1069–1081. [Google Scholar] [CrossRef]
- Wang, J.J.; Zhou, Y.P.; Lin, W.H.; Li, M.M.; Wang, M.N.; Wang, Z.G.; Kuang, Y.; Tian, P. Effect of an Epichloë endophyte on adaptability to water stress in Festuca sinensis. Fungal Ecol. 2017, 30, 39–47. [Google Scholar] [CrossRef]
- Gundel, P.E.; Irisarri, J.; Fazio, L.; Casas, C.; Pérez, L.I. Inferring field performance from drought experiments can be misleading: The case of symbiosis between grasses and Epichloë fungal endophytes. J. Arid Environ. 2016, 132, 60–62. [Google Scholar] [CrossRef]
- Saari, S.; Faeth, S.H. Hybridization of Neotyphodium endophytes enhances competitive ability of the host grass. New Phytol. 2012, 195, 231–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, K.H. Effects of endophyte infection on drought stress tolerance of Lolium perenne accessions from the mediterranean region. Environ. Exp. Bot. 2011, 71, 337. [Google Scholar] [CrossRef]
- Kannadan, S.; Rudgers, J.A. Endophyte symbiosis benefits a rare grass under low water availability. Funct. Ecol. 2010, 22, 706–713. [Google Scholar] [CrossRef]
- Malinowski, D.; Leuchtmann, A.; Schmidt, D.; NÖsberger, J. Growth and water status in meadow fescue is affected by Neotyphodium and Phialophora species endophytes. Agron. J. 1997, 89, 673–678. [Google Scholar] [CrossRef]
- Elbersen, H.W.; West, C.P. Growth and water relations of field grown tall fescue as influenced by drought and endophyte. Grass Forage Sci. 1996, 51, 333–342. [Google Scholar] [CrossRef]
- Vázquez-dE−Aldana, B.R.; García-Ciudad, A.; García-Criado, B.; VicentE−Tavera, S.; Zabalgogeazcoa, I. Fungal endophyte (Epichloë festucae) alters the nutrient content of Festuca rubra regardless of water availability. PLoS ONE 2013, 8, e84539. [Google Scholar] [CrossRef] [PubMed]
- Elmi, A.A.; West, C.P. Endophyte infection effects on stomatal conductance, osmotic adjustment and drought recovery of tall fescue. New Phytol. 1995, 131, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Inukai, Y.; Sakamoto, T.; Ueguchi-Tanaka, M.; Shibata, Y.; Gomi, K.; Umemura, I.; Hasegawa, Y.; Ashikari, M.; Kitano, H.; Matsuoka, M. Crown rootless1, which is essential for crown root formation in rice, is a target of an auxin response factor in auxin signaling. Plant Cell 2005, 17, 1387–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabzalian, M.R.; Mirlohi, A. Neotyphodium endophytes trigger salt resistance in tall and meadow fescues. J. Plant Nutr. Soil Sci. 2010, 173, 952–957. [Google Scholar] [CrossRef]
- Nagabhyru, P.; Dinkins, R.D.; Wood, C.L.; Bacon, C.W.; Schardl, C.L. Tall fescue endophyte effects on tolerance to water-deficit stress. BMC Plant Biol. 2013, 13, 127. [Google Scholar] [CrossRef] [Green Version]
- Hesse, U.; Schoberlein, W.; Wittenmayert, L.; Forster, K.; Warnstorn, K.; Diepenbrock, W.; Merbacht, W. Effect of Neotyphodium endophytes on growth reproduction and drought-stress tolerance of three Lolium perenne L. genotypes. Grass Forage Sci. 2003, 58, 407–415. [Google Scholar] [CrossRef]
- Rozpądek, P.; Wężowicz, K.; Nosek, M.; Ważny, R.; Tokarz, K.; Lembicz, M.; Miszalski, Z.; Turnau, K. The fungal endophyte Epichloë typhina improves photosynthesis efficiency of its host orchard grass (Dactylis glomerata). Planta 2015, 242, 1025–1035. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Gao, Y.B.; Chen, S.P.; Ren, A.Z. Effects of Endophyte Infection on Photosynthesis, Transpiration and Water Use Efficiency of Lolium perenne L. Under Drought Stress. Chin. J. Plant Ecol. 2001, 25, 537–543. [Google Scholar]
- Morse, L.J.; Faeth, S.H.; Day, T.A. Neotyphodium interactions with a wild grass are driven mainly by endophyte haplotype. Funct. Ecol. 2007, 21, 813–822. [Google Scholar] [CrossRef]
- Xia, C.; Christensen, M.J.; Zhang, X.X.; Nan, Z.B. Effect of Epichloë gansuensis endophyte and transgenerational effects on the water use efficiency, nutrient and biomass accumulation of achnatherum inebrians under soil water deficit. Plant Soil 2018, 424, 1–17. [Google Scholar] [CrossRef]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Itai, C. Role of phytohormones in plant responses to stresses. In Plant Response to Environmental Stresses: From Phytohormones to Genome Reorganization; Lerner, H.R., Ed.; Marcel Dekker: New York, NY, USA, 1999; pp. 287–301. [Google Scholar]
- Zhang, X.; Ervin, E.H.; Evanylo, G.K. Impact of Biosolids on Hormone Metabolism in Drought-Stressed Tall Fescue. Crop. Sci. 2009, 49, 1893–1901. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and toletance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, S.; Davies, W.J. Drought, ozone, aba and ethylene: New insights from cell to plant to community. Plant Cell Environ. 2010, 33, 510–525. [Google Scholar] [CrossRef] [PubMed]
- Pospisilova, J. Participation of phytohormones in the stomatal regulation of gas exchange during water stress. Biol. Plant 2003, 46, 491–506. [Google Scholar] [CrossRef]
- Chow, B.; McCourt, P. Hormone signalling from a developmental context. J. Exp. Bot. 2004, 55, 247–251. [Google Scholar] [CrossRef]
- Pospisilova, J.; Vagner, M.; Malbeck, J.; Travnickova, A.; Batkova, P. Interactions between abscisic acid and cytokinins during water stress and subsequent rehydration. Biol. Plant 2005, 49, 533–540. [Google Scholar] [CrossRef]
- Acharya, B.R.; Assmann, S.M. Hormone interactions in stomatal function. Plant Mol. Biol. 2009, 69, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, R.; Watanabe, Y.; Fujita, Y.; Le, D.T.; Kojima, M. Analysis of Cytokinin Mutants and Regulation of Cytokinin Metabolic Genes Reveals Important Regulatory Roles of Cytokinins in Drought, Salt and Abscisic Acid Responses, and Abscisic Acid Biosynthesis. Plant Cell 2011, 23, 2169–2183. [Google Scholar] [CrossRef] [Green Version]
- Werner, T.; Nehnevajova, E.; Kollmer, I.; Novak, O.; Strnad, M.; Kramer, U.; Schmulling, T. Root-specific reduction of cytokinin causes enhanced root growth, drought tolerance, and leaf mineral enrichment in arabidopsis and tobacco. Plant Cell 2010, 22, 3905–3920. [Google Scholar] [CrossRef] [Green Version]
- Werner, T.; Köllmer, I.; Bartrina, I.; Holst, K.; Schmülling, T. New insights into the biology of cytokinin degradation. Plant Biol. 2006, 8, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Jones, M.L.; Banowetz, G.M.; Clark, D.G. Overproduction of cytokinins in petunia flowers transformed with P (SAG12)-IPT delays corolla senescence and decreases sensitivity to ethylene. Plant Physiol. 2003, 132, 2174–2183. [Google Scholar] [CrossRef] [Green Version]
- Zdunek, E.; Lips, S.H. Transport and accumulation rates of abscisic acid and aldehyde oxidase activity in Pisum sativum L. in response to suboptimal growth conditions. J. Exp. Bot. 2001, 52, 1269–1276. [Google Scholar] [CrossRef] [Green Version]
- Bao, G.S.; Song, M.L.; Wang, Y.Q. Interactive effects of different densities of Pedicularis kansuensis parasitism and Epichloë endophyte infection on the endogenous hormone levels and alkaloid contents of Stipa purpurea. Acta Prataculturae Sin. 2020, 29, 147–156. [Google Scholar]
- Chen, S.P.; Gao, Y.B.; Liang, Y.; Ren, A.Z. Effect of endophyte infection on the concentrations of free proline and aba of leaves of Lolium perenne L. under water stress. Acta Ecol. Sin. 2001, 21, 1964–1972, (In Chinese with English Abstract). [Google Scholar]
- Yan, Z.L.; Xuan, C.X.; Niu, J.Y.; Xi, L.L.; Liu, J.H.; Zhao, T.W. Effect of drought stress and water recovery on endogenous hormone content in roots of pea. Chin. J. Eco-Agric. 2009, 17, 297–301, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Woo, Y.M.; Park, H.J.; Yang, J.I.; Park, J.J.; Back, K.; Park, Y.M.; An, G. Constitutively wilted 1, a member of the rice YUCCA gene family, is required for maintaining water homeostasis and all appropriate root to shoot ratio. Plant Mol. Biol. 2007, 65, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.C.; Jiang, J.G. Osmotic adjustment and plant adaptation to environmental changes related to drought and salinity. Environ. Rev. 2010, 18, 309–319. [Google Scholar] [CrossRef]
- Dacosta, M.; Huang, B. Osmotic adjustment associated with variation in bentgrass tolerance to drought stress. J. Am. Soc. Hortic. Sci. 2006, 131, 338–344. [Google Scholar] [CrossRef]
- White, P.J. Improving potassium acquisition and utilisation by crop plants. J. Plant Nutr. Soil Sci. 2013, 176, 305–316. [Google Scholar] [CrossRef]
- Wakeel, A.; Farooq, M.; Qadir, M.; Schubert, S. Potassium substitution by sodium in plants. Crit. Rev. Plant Sci. 2011, 30, 401–413. [Google Scholar] [CrossRef]
- Bayat, F.; Mirlohi, A.; Khodambashi, M. Effects of endophytic fungi on some drought tolerance mechanisms of tall fescue in a hydroponics culture. Russ. J. Plant Phys. 2009, 56, 510–516. [Google Scholar] [CrossRef]
- Hu, X.; Jiang, M.; Zhang, J.; Zhang, A.; Lin, F.; Tan, M. Calcium-calmodulin is required for abscisic acid-induced antioxidant defense and functions both upstream and downstream of H2O2 production in leaves of maize (Zea mays) plants. New Phytol. 2007, 173, 27–38. [Google Scholar] [CrossRef]
- Rentel, M.C.; Knight, M.R. Oxidative stress-induced calcium signaling in Arabidopsis. Plant Physiol. 2004, 135, 1471–1479. [Google Scholar] [CrossRef] [Green Version]
- Berman-Frank, I.; Dubinsky, Z. Balanced growth in aquatic plants: Myth or reality? Phytoplankton use the imbalance between carbon assimilation and biomass production to their strategic advantage. Bioscience 1999, 49, 29–37. [Google Scholar] [CrossRef]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2011, 62, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Sardans, J.; Rivas-Ubach, A.; Peñuelas, J. The C:N:P stoichiometry of organisms and ecosystems in changing world: A review and perspectives. Perspect. Plant Ecol. 2012, 14, 33–47. [Google Scholar] [CrossRef]
- Khan, M.H.; Meghvansi, M.K.; Gupta, R.; Veer, V.; Singh, L.; Kalita, M.C. Foliar spray with vermiwash modifies the arbuscular mycorrhizal dependency and nutrient stoichiometry of bhut jolokia (Capsicum assamicum). PLoS ONE 2014, 9, e92318. [Google Scholar] [CrossRef]
- Faeth, S.H.; Fagan, W.F. Fungal endophytes: Common host plant symbionts but uncommonmutualists. Integr. Comp. Biol. 2002, 42, 360–368. [Google Scholar] [CrossRef] [Green Version]
- Belesky, D.P.; Stuedemann, J.A.; Plattner, R.D.; Wilkinson, S.R. Ergopeptine alkaloids in grazed tall fescue. Agron. J. 1988, 80, 209–212. [Google Scholar] [CrossRef] [Green Version]
- Fujita, Y.; Robroek, B.; Ruiter, P.; Heil, G.W.; Wassen, M.J. Increased N affects P uptake of eight grassland species: The role of root surface phosphatase activity. Oikos 2010, 119, 1665–1673. [Google Scholar] [CrossRef]
- Hessen, D.O.; Jensen, T.C.; Kyle, M.; Elser, J.J. RNA responses to N- and P-limitation; reciprocal regulation of stoichiometry and growth rate in Brachionus. Functl. Ecol. 2007, 21, 956–962. [Google Scholar] [CrossRef]
- Vrede, T.; Dobberfuhl, D.R.; Kooijman, S.A.L.M.; Elser, J.J. Fundamental connections among organism C:N:P stoichiometry, macromolecular composition, and growth. Ecology 2004, 85, 1217–1229. [Google Scholar] [CrossRef] [Green Version]
- Elser, J.J.; O’brien, W.J.; Dobberfuhl, D.R.; Dowling, T.E. The evolution of ecosystem processes: Growth rate and elemental stoichiometry of a key herbivore in temperate and arctic habitats. J. Evol. Biol. 2000, 13, 845–853. [Google Scholar] [CrossRef] [Green Version]
- Elser, J.J.; Dobberfuhl, D.R.; Mac Kay, N.A.; Schampel, J.H. Organism size, life history, and N:P stoichiometry. Bioscience 1996, 46, 674–684. [Google Scholar] [CrossRef] [Green Version]
- Elser, J.J.; Hamilton, A. Stoichiometry and the new biology: The future is now. PLoS Biol. 2007, 5, e181. [Google Scholar] [CrossRef]
- Queiroz, R.; Cazetta, J.O. Proline and trehalose in maize seeds germinating under low osmotic potentials. Rev. Bras. Eng. Agric. Ambient. 2016, 20, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Blanch, M.; Sanchez-Ballesta, M.T.; Escribano, M.I.; Merodio, C. The relationship between bound water and carbohydrate reserves in association with cellular integrity in fragaria vesca stored under different conditions. Food Bioprocess Technol. 2015, 8, 875–884. [Google Scholar] [CrossRef]
- Ren, F.; Zhou, H.K.; Zhao, X.Q.; Han, F.; Shi, L.N.; Duan, J.C.; Zhao, J.Z. Influence of simulated warming using OTC on physiological–biochemical characteristics of elymus nutans in alpine meadow on qinghai-tibetan plateau. Acta Ecol. Sin. 2010, 30, 166–171. [Google Scholar] [CrossRef]
- Wang, Q.H.; Dong, S.L.; Tian, X.L.; Wang, F. Effects of circadian rhythms of fluctuating temperature on growth and biochemical composition of Ulva pertusa. Hydrobiologia 2007, 586, 313–319. [Google Scholar] [CrossRef]
- Schardl, C.L.; Grossman, R.B.; Nagabhyru, P.; Faulkner, J.R.; Mallik, U.P. Loline alkaloids: Currencies of mutualism. Phytochemistry 2007, 68, 980–996. [Google Scholar] [CrossRef] [PubMed]
- Schardl, C.L.; Young, C.A.; Hesse, U.; Amyotte, S.G.; Andreeva, K.; Calie, P.J.; Fleetwood, D.J.; Haws, D.C.; Moore, N.; Oeser, B.; et al. Plant-symbiotic fungi as chemical engineers: Multi-genome analysis of the clavicipitaceae reveals dynamics of alkaloid loci. PLoS Genet. 2013, 9, e1003323. [Google Scholar] [CrossRef] [Green Version]
- Blankenship, J.D.; Houseknecht, J.B.; Pal, S.; Bush, L.P.; Grossman, R.B.; Schardl, C.L. Biosynthetic precursors of fungal pyrrolizidines, the loline alkaloids. ChemBioChem 2005, 6, 1016–1022. [Google Scholar] [CrossRef]
- Bush, L.P.; Fannin, F.F.; Siegel, M.R.; Dahlman, D.L.; Burton, H.R. Chemistry, occurrence and biological effects of pyrrolizidine alkaloids associated with endophytE−grass associations. Agric. Ecosyst. Environ. 1993, 44, 81–102. [Google Scholar] [CrossRef]
- Bacon, C.W. Abiotic stress tolerances (moisture, nutrients) and photosynthesis in endophyte infected tall fescue. Agric. Ecosyst. Environ. 1993, 44, 123–141. [Google Scholar] [CrossRef]
- Kennedy, C.W.; Bush, L.P. Effect of environmental and management factors on the accumulation of N-acetyl and N-formyl loline alkaloids in tall fescue. Crop. Sci. 1983, 23, 547–552. [Google Scholar] [CrossRef]
- Malinowski, D.; Leuchtmann, A.; Schmidt, D.; NÖsberger, J. Symbiosis with Neotyphodium uncinatum endophyte may increase the competitive ability of meadow fescue. Agron. J. 1997, 89, 833–839. [Google Scholar] [CrossRef]
- Zhang, X.X.; Li, C.J.; Nan, Z.B. Effects of salt and drought stress on alkaloid production in endophytE−infected drunken horse grass (Achnatherum inebrians). Biochem. Syst. Ecol. 2011, 39, 471–476. [Google Scholar] [CrossRef]
- Saikkonen, K.; Lehtonen, P.; Helander, M.; Koricheva, J.; Faeth, S.H. Model systems in ecology: Dissecting the endophytegrass literature. Trends Plant Sci. 2006, 11, 428–433. [Google Scholar] [CrossRef]
- Cheplick, G.P. Recovery from drought stress in Lolium perenne (Poaceae): Are fungal endophytes detrimental? Am. J. Bot. 2004, 91, 1960–1968. [Google Scholar] [CrossRef] [PubMed]
- Faeth, S.H.; Sullivan, T.J. Mutualistic asexual endophytes in a native grass are usually parasitic? Am. Nat. 2003, 161, 310–325. [Google Scholar] [CrossRef] [PubMed]
- Hesse, U.; Schöberlein, W.; Wittenmayer, L.; Förster, K.; Warnstorff, K.; Diepenbrock, W.; Merbach, W. Influence of water supply and endophyte infection (Neotyphodium spp.) on vegetative and reproductive growth of two Lolium perenne L. genotypes. Eur. J. Agron. 2005, 22, 45–54. [Google Scholar] [CrossRef]
- Nan, Z.B. Incidence and distribution of endophytic fungi in seeds and plants of some native and introduced grasses in China. Acta Prataculturae Sin. 1996, 5, 13–17, (In Chinese with English Abstract). [Google Scholar]
- Harrington, J.T.; Mexal, J.G.; Fisher, J.T. Volume Displacement Provides a Quick and Accurate Way to Quantify New Root Production. Tree Plant. Notes 2012, 45, 121–124. [Google Scholar]
- Hanway, J.J.; Heidel, H. Soil analysis methods as used in lowa state college soil testing laboratory. Iowa Agric. 1952, 57, 1–31. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Li, M.; Lin, W.; Nan, Z.; Tian, P. Effects of Epichloë sinensis Endophyte and Host Ecotype on Physiology of Festuca sinensis under Different Soil Moisture Conditions. Plants 2021, 10, 1649. https://doi.org/10.3390/plants10081649
Xu W, Li M, Lin W, Nan Z, Tian P. Effects of Epichloë sinensis Endophyte and Host Ecotype on Physiology of Festuca sinensis under Different Soil Moisture Conditions. Plants. 2021; 10(8):1649. https://doi.org/10.3390/plants10081649
Chicago/Turabian StyleXu, Wenbo, Miaomiao Li, Weihu Lin, Zhibiao Nan, and Pei Tian. 2021. "Effects of Epichloë sinensis Endophyte and Host Ecotype on Physiology of Festuca sinensis under Different Soil Moisture Conditions" Plants 10, no. 8: 1649. https://doi.org/10.3390/plants10081649
APA StyleXu, W., Li, M., Lin, W., Nan, Z., & Tian, P. (2021). Effects of Epichloë sinensis Endophyte and Host Ecotype on Physiology of Festuca sinensis under Different Soil Moisture Conditions. Plants, 10(8), 1649. https://doi.org/10.3390/plants10081649