
Molecular Characterization, Expression Analysis of Carotenoid, Xanthophyll, Apocarotenoid Pathway Genes, and Carotenoid and Xanthophyll Accumulation in Chelidonium majus L.

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification, Protein Nomenclature, and Sequence Analysis of CBP, XBP, and ABP Genes
2.2. Phylogenetic and Homology Analysis
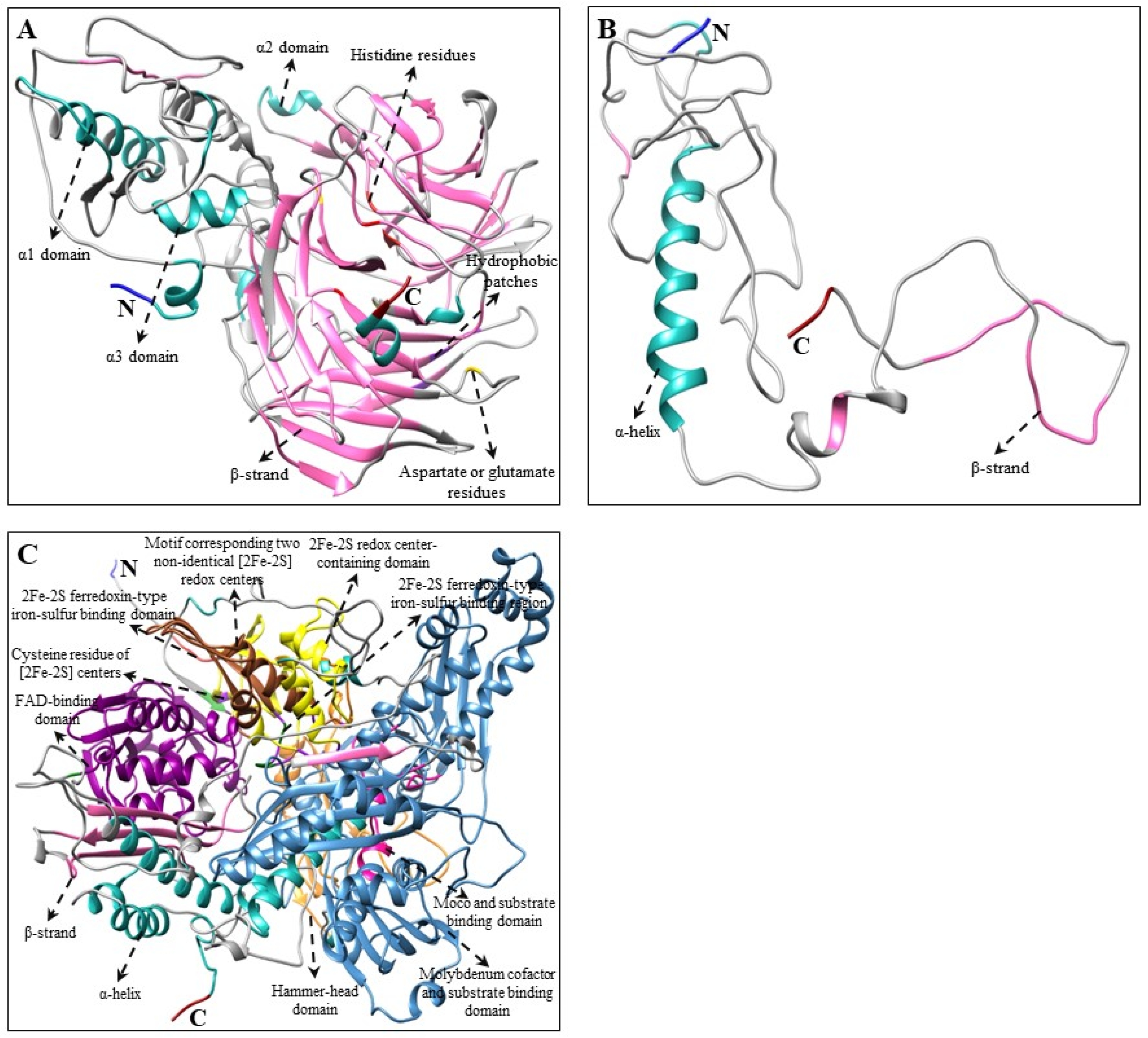
2.3. Multiple Alignments and Tertiary Structure Analysis of CBP, XBP, and ABP Genes
2.4. Subcellular Location Prediction of CBP, XBP, and ABP Genes by In Silico Analysis
2.5. CBP, XBP, and ABP Gene Expression Levels in Different Parts of C. majus
2.6. Carotenoid and Xanthophyll Level in Different Parts of C. majus
3. Materials and Methods
3.1. Plant Materials
3.2. Identification and Sequence Analysis of CBP, XBP, and ABP Genes
3.3. Structural Analysis of CBP, XBP, and ABP Genes
3.4. Phylogenetic Analysis and Percent Identity Matrix
3.5. RNA Extraction and cDNA Synthesis
3.6. CBP, XBP, and ABP Genes Expression
3.7. Carotenoid Extractions and HPLC Analysis
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Orvos, P.; Virág, L.; Tálosi, L.; Hajdú, Z.; Csupor, D.; Jedlinszki, N.; Szél, T.; Varró, A.; Hohmann, J. Effects of Chelidonium majus extracts and major alkaloids on hERG potassium channels and on dog cardiac action potential—A safety approach. Fitoterapia 2015, 100, 156–165. [Google Scholar] [CrossRef] [Green Version]
- Colombo, M.L.; Bosisio, E. Pharmacological activities of Chelidonium majus L. (papaveraceae). Pharmacol. Res. 1996, 33, 127–134. [Google Scholar] [CrossRef]
- Horváth, G.; Molnár, P.; Farkas, A.; Szabó, L.G.; Turcsi, E.; Deli, J. Separation and identification of carotenoids in flowers of Chelidonium majus L. and inflorescences of Solidago canadensis L. Chromatographia 2010, 71, 103–108. [Google Scholar] [CrossRef]
- Kędzia, B.; Łożykowska, K.; Gryszczyńska, A. Skład chemiczny i zawartość substancji biologicznie aktywnych w Chelidonium majus L. Postępy Fitoterapii 2013, 3, 174–181. [Google Scholar]
- Mikołajczak, P.Ł.; Kędzia, B.; Ożarowski, M.; Kujawski, R.; Bogacz, A.; Bartkowiak-Wieczorek, J.; Białas, W.; Gryszczyńska, A.; Buchwald, W.; Szulc, M. Evaluation of anti-inflammatory and analgesic activities of extracts from herb of Chelidonium majus L. Cent. Eur. J. Immunol. 2015, 40, 400. [Google Scholar] [CrossRef] [Green Version]
- Nawrot, R.; Zauber, H.; Schulze, W.X. Global proteomic analysis of Chelidonium majus and Corydalis cava (Papaveraceae) extracts revealed similar defense-related protein compositions. Fitoterapia 2014, 94, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Stancic-Rotaru, M.; Mititelu, M.; Crasmaru, M.; Balaban, D. Spectroanalytical profile of flavonoids from Chelidonium majus L. Rom. Biotechnol. Lett. 2002, 8, 1093–1100. [Google Scholar]
- Gilca, M.; Gaman, L.; Panait, E.; Stoian, I.; Atanasiu, V. Chelidonium majus—An integrative review: Traditional knowledge versus modern findings. J. Complement. Med. Res. 2010, 17, 241–248. [Google Scholar] [CrossRef]
- Khodabande, Z.; Jafarian, V.; Sariri, R. Antioxidant activity of Chelidonium majus extract at phenological stages. Appl. Biol. Chem. 2017, 60, 497–503. [Google Scholar] [CrossRef]
- Orland, A.; Knapp, K.; König, G.; Ulrich-Merzenich, G.; Knöß, W. Combining metabolomic analysis and microarray gene expression analysis in the characterization of the medicinal plant Chelidonium majus L. Phytomedicine 2014, 21, 1587–1596. [Google Scholar] [CrossRef]
- Wouters, A.; Lardon, F.; Gravekamp, C.; Apers, S.; Pieters, L. In vitro and in vivo investigations on the antitumour activity of Chelidonium majus. Phytomedicine 2015, 22, 1279–1287. [Google Scholar]
- Zielińska, S.; Czerwińska, M.E.; Dziągwa-Becker, M.; Dryś, A.; Kucharski, M.; Jezierska-Domaradzka, A.; Płachno, B.J.; Matkowski, A. Modulatory effect of Chelidonium majus extract and its alkaloids on LPS-stimulated cytokine secretion in human neutrophils. Molecules 2020, 25, 842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, D.; Sharma, A. A review on phytochemical and pharmacological potential of genus Chelidonium. Pharmacogn. J. 2013, 5, 184–190. [Google Scholar] [CrossRef]
- Molnár, P.; Kawase, M.; Motohashi, N. Isolation, crystallization and handling of carotenoids and (E/Z)-isomerization of carotenoids. In Functional Polyphenols and Carotenes with Antioxidative Action; Motohashi, N., Ed.; Research Signpost: Trivandrum, India, 2005; pp. 111–131. [Google Scholar]
- Sathasivam, R.; Radhakrishnan, R.; Kim, J.K.; Park, S.U. An update on biosynthesis and regulation of carotenoids in plants. S. Afr. J. Bot. 2020, 1–13. [Google Scholar] [CrossRef]
- Sathasivam, R.; Kermanee, P.; Roytrakul, S.; Juntawong, N. Isolation and molecular identification of β-carotene producing strains of Dunaliella salina and Dunaliella bardawil from salt soil samples by using species-specific primers and internal transcribed spacer (ITS) primers. Afr. J. Biotechnol. 2012, 11, 16677–16687. [Google Scholar]
- Sathasivam, R.; Pongpadung, P.; Praiboon, J.; Chirapart, A.; Trakulnaleamsai, S.; Roytrakul, S.; Juntawong, N. Optimizing NaCl and KNO3 concentrations for high β-carotene production in photobioreactor by Dunaliella salina KU11 isolated from saline soil sample. Chiang Mai J. Sci. 2018, 45, 106–115. [Google Scholar]
- Sathasivam, R.; Praiboon, J.; Chirapart, A.; Trakulnaleamsai, S.; Kermanee, P.; Roytrakul, S.; Juntawong, N. Screening, phenotypic and genotypic identification of β-carotene producing strains of Dunaliella salina from Thailand. Indian J. Geo-Mar. Sci. 2014, 43, 2198–2216. [Google Scholar]
- Sathasivam, R.; Radhakrishnan, R.; Hashem, A.; Abd_Allah, E.F. Microalgae metabolites: A rich source for food and medicine. Saudi J. Biol. Sci. 2019, 26, 709–722. [Google Scholar] [CrossRef]
- Sathasivam, R.; Ki, J.-S. A review of the biological activities of microalgal carotenoids and their potential use in healthcare and cosmetic industries. Mar. Drugs 2018, 16, 26. [Google Scholar] [CrossRef] [Green Version]
- Sathasivam, R.; Ki, J.-S. Differential transcriptional responses of carotenoid biosynthesis genes in the marine green alga Tetraselmis suecica exposed to redox and non-redox active metals. Mol. Biol. Rep. 2019, 46, 1167–1179. [Google Scholar] [CrossRef]
- Hughes, D.A. Effects of carotenoids on human immune function. Proc. Nutr. Soc. 1999, 58, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Stanley, L.; Yuan, Y.-W. Transcriptional regulation of carotenoid biosynthesis in plants: So many regulators, so little consensus. Front. Plant Sci. 2019, 10, 1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flowerika, A.A.; Kumar, J.; Thakur, N.; Pandey, A.; Pandey, A.K.; Upadhyay, S.K.; Tiwari, S. Characterization and expression analysis of phytoene synthase from bread wheat (Triticum aestivum L.). PLoS ONE 2016, 11, e0162443. [Google Scholar] [CrossRef] [PubMed]
- Karvouni, Z.; John, I.; Taylor, J.E.; Watson, C.F.; Turner, A.J.; Grierson, D. Isolation and characterization of a melon cDNA clone encoding phytoene synthase. Plant Mol. Biol. 1995, 27, 1153–1162. [Google Scholar] [CrossRef]
- Kato, M.; Ikoma, Y.; Matsumoto, H.; Sugiura, M.; Hyodo, H.; Yano, M. Accumulation of carotenoids and expression of carotenoid biosynthetic genes during maturation in citrus fruit. Plant Physiol. 2004, 134, 824–837. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.J.; Ko, K.C.; Kim, C.S.; Chung, W.I. Isolation and expression patterns of a cDNA encoding phytoene synthase in citrus. J. Plant Physiol. 2001, 158, 795–800. [Google Scholar] [CrossRef]
- Obrero, A.; Gonzalez-Verdejo, C.I.; Roman, B.; Gomez, P.; Die, J.V.; Ampomah-Dwamena, C. Identification, cloning, and expression analysis of three phytoene synthase genes from Cucurbita pepo. Biol. Plant. 2015, 59, 201–210. [Google Scholar] [CrossRef]
- Reddy, C.S.; Lee, S.H.; Yoon, J.S.; Kim, J.K.; Lee, S.W.; Hur, M.; Koo, S.C.; Kim, M.R.; Lee, W.M.; Jang, J.K.; et al. Molecular cloning and characterization of carotenoid pathway genes and carotenoid content in Ixeris dentata var. albiflora. Molecules 2017, 22, 1449. [Google Scholar] [CrossRef] [Green Version]
- Salvini, M.; Bernini, A.; Fambrini, M.; Pugliesi, C. cDNA cloning and expression of the phytoene synthase gene in sunflower. J. Plant Physiol. 2005, 162, 479–484. [Google Scholar] [CrossRef]
- Tuan, P.A.; Kim, Y.B.; Kim, J.K.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.U. Molecular characterization of carotenoid biosynthetic genes and carotenoid accumulation in Scutellaria baicalensis Georgi. J. Exp. Clin. Sci. 2015, 14, 146–157. [Google Scholar]
- Zhang, J.C.; Tao, N.G.; Xu, Q.; Zhou, W.J.; Cao, H.B.; Xu, J.A.; Deng, X.X. Functional characterization of Citrus PSY gene in Hongkong kumquat (Fortunella hindsii Swingle). Plant Cell Rep. 2009, 28, 1737–1746. [Google Scholar] [CrossRef]
- Li, C.; Ji, J.; Wang, G.; Li, Z.D.; Wang, Y.R.; Fan, Y.J. Over-expression of LcPDS, LcZDS, and LcCRTISO, genes from wolfberry for carotenoid biosynthesis, enhanced carotenoid accumulation, and salt tolerance in tobacco. Front. Plant Sci. 2020, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.Q.; Li, L.X.; Zhan, P.L.; Wang, Q. Cloning and functional characterization of phytoene desaturase in Andrographis paniculata. Zhongguo Zhong Yao Za Zhi 2015, 40, 3760–3765. [Google Scholar] [PubMed]
- Chen, Y.; Li, F.Q.; Wurtzel, E.T. Isolation and characterization of the Z-ISO gene encoding a missing component of carotenoid biosynthesis in plants. Plant Physiol. 2010, 153, 66–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Wang, Y.G.; Fu, J.X.; Bao, Z.Y.; Zhao, H.B. Transcriptomic analysis and carotenogenic gene expression related to petal coloration in Osmanthus fragrans ’Yanhong Gui’. Trees 2016, 30, 1207–1223. [Google Scholar] [CrossRef]
- Araya-Garay, J.; Feijoo-Siota, L.; Veiga-Crespo, P.; Sanchez-Perez, A.; Gonzalez Villa, T. Cloning and functional expression of zeta-carotene desaturase, a novel carotenoid biosynthesis gene from Ficus carica. Int. J. Microbiol. Adv. Immunol. 2014, 2, 32–40. [Google Scholar]
- Flores-Ortiz, C.; Alvarez, L.M.; Undurraga, A.; Arias, D.; Duran, F.; Wegener, G.; Stange, C. Differential role of the two ζ-carotene desaturase paralogs in carrot (Daucus carota): ZDS1 is a functional gene essential for plant development and carotenoid synthesis. Plant Sci. 2020, 291, 110327. [Google Scholar] [CrossRef]
- Kim, I.J.; Nam, T.S.; Kim, C.S.; Song, K.J.; Lee, D.H. Isolation and expression patterns of a zeta-carotene desaturase gene from citrus. Hortic. Environ. Biotechnol. 2010, 51, 551–555. [Google Scholar]
- Li, Z.D.; Wu, G.X.; Ji, J.; Wang, G.; Tian, X.W.; Gao, H.L. Cloning and expression of a ζ-carotene desaturase gene from Lycium chinense. J. Genet. 2015, 94, 287–294. [Google Scholar] [CrossRef]
- Li, P.R.; Zhang, S.J.; Zhang, S.F.; Li, F.; Zhang, H.; Liu, X.Y.; Wu, J.; Wang, X.W.; Sun, R.F. Carotenoid identification and molecular analysis of carotenoid isomerase-encoding BrCRTISO, the candidate gene for inner leaf orange coloration in Chinese cabbage. Mol. Breed. 2015, 35, 72. [Google Scholar] [CrossRef]
- Pinheiro, T.T.; Peres, L.E.P.; Purgatto, E.; Latado, R.R.; Maniero, R.A.; Martins, M.M.; Figueira, A. Citrus carotenoid isomerase gene characterization by complementation of the “Micro-Tom” tangerine mutant. Plant Cell Rep. 2019, 38, 623–636. [Google Scholar] [CrossRef]
- Cunningham, F.X.; Pogson, B.; Sun, Z.R.; McDonald, K.A.; DellaPenna, D.; Gantt, E. Functional analysis of the beta and epsilon lycopene cyclase enzymes of Arabidopsis reveals a mechanism for control of cyclic carotenoid formation. Plant Cell 1996, 8, 1613–1626. [Google Scholar] [CrossRef] [Green Version]
- Devitt, L.C.; Fanning, K.; Dietzgen, R.G.; Holton, T.A. Isolation and functional characterization of a lycopene beta-cyclase gene that controls fruit colour of papaya (Carica papaya L.). J. Exp. Bot. 2010, 61, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koc, I.; Filiz, E.; Tombuloglu, H. Comparative analysis of plant lycopene cyclases. Comput. Biol. Chem. 2015, 58, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.W.; Zhang, Y.; Zheng, X.J.; Zhu, K.J.; Xu, Q.; Deng, X.X. Isolation and functional characterization of a lycopene β-cyclase gene promoter from citrus. Front. Plant Sci. 2016, 7, 1367. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.; Wang, C.; Chen, X.; Zang, M.L.; Yuan, C.H.; Wang, X.T.; Wang, Q.; Li, M.; Li, X.Y.; Chen, L.; et al. The lycopene β-cyclase plays a significant role in provitamin A biosynthesis in wheat endosperm. BMC Plant Biol. 2015, 15, 112. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.C.; Zhou, W.J.; Xu, Q.; Tao, N.G.; Ye, J.L.; Guo, F.; Xu, J.; Deng, X.X. Two lycopene β-cyclases genes from sweet orange (Citrus sinensis L. Osbeck) encode enzymes with different functional efficiency during the conversion of lycopene-to-provitamin A. J. Integr. Agric. 2013, 12, 1731–1747. [Google Scholar] [CrossRef]
- Zhao, D.Q.; Zhou, C.H.; Sheng, Y.L.; Liang, G.H.; Tao, J. Molecular cloning and expression of phytoene synthase, lycopene beta-cyclase, and beta-carotene hydroxylase genes in persimmon (Diospyros kaki L.) fruits. Plant Mol. Biol. Rep. 2011, 29, 345–351. [Google Scholar] [CrossRef]
- Zhu, H.S.; Chen, M.D.; Wen, Q.F.; Li, Y.P. Isolation and characterization of the carotenoid biosynthetic genes LCYB, LCYE and CHXB from strawberry and their relation to carotenoid accumulation. Sci. Hortic. 2015, 182, 134–144. [Google Scholar] [CrossRef]
- Gao, Z.; Liu, Q.; Zheng, B.; Chen, Y. Molecular characterization and primary functional analysis of PeVDE, a violaxanthin de-epoxidase gene from bamboo (Phyllostachys edulis). Plant Cell Rep. 2013, 32, 1381–1391. [Google Scholar] [CrossRef]
- Li, X.; Zhao, W.; Sun, X.; Huang, H.; Kong, L.; Niu, D.; Sui, X.; Zhang, Z. Molecular cloning and characterization of violaxanthin de-epoxidase (CsVDE) in cucumber. PLoS ONE 2013, 8, e64383. [Google Scholar] [CrossRef]
- Jeknić, Z.; Morré, J.T.; Jeknić, S.; Jevremović, S.; Subotić, A.; Chen, T.H. Cloning and functional characterization of a gene for capsanthin-capsorubin synthase from tiger lily (Lilium lancifolium Thunb.’Splendens’). Plant Cell Physiol. 2012, 53, 1899–1912. [Google Scholar] [CrossRef]
- Ahrazem, O.; Rubio-Moraga, A.; Berman, J.; Capell, T.; Christou, P.; Zhu, C.F.; Gomez-Gomez, L. The carotenoid cleavage dioxygenase CCD2 catalysing the synthesis of crocetin in spring crocuses and saffron is a plastidial enzyme. New Phytol. 2016, 209, 650–663. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.C.; Molnar, P.; Schwab, W. Cloning and functional characterization of carotenoid cleavage dioxygenase 4 genes. J. Exp. Bot. 2009, 60, 3011–3022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigo, M.J.; Alquezar, B.; Alos, E.; Medina, V.; Carmona, L.; Bruno, M.; Al-Babili, S.; Zacarias, L. A novel carotenoid cleavage activity involved in the biosynthesis of citrus fruit-specific apocarotenoid pigments. J. Exp. Bot. 2013, 64, 4461–4478. [Google Scholar] [CrossRef]
- Tuan, P.A.; Kim, J.K.; Lee, S.; Chae, S.C.; Park, S.U. Molecular characterization of carotenoid cleavage dioxygenases and the effect of gibberellin, abscisic acid, and sodium chloride on the expression of genes involved in the carotenoid biosynthetic aathway and carotenoid accumulation in the callus of Scutellaria baicalensis Georgi. J. Agric. Food Chem. 2013, 61, 5565–5572. [Google Scholar]
- Zheng, X.J.; Xie, Z.Z.; Zhu, K.J.; Xu, Q.; Deng, X.X.; Pan, Z.Y. Isolation and characterization of carotenoid cleavage dioxygenase 4 genes from different citrus species. Mol. Genet. Genom. 2015, 290, 1589–1603. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.C.; Joseph, L.M.; Deng, W.T.; Liu, L.J.; Li, Q.B.; Cline, K.; McCarty, D.R. Molecular characterization of the Arabidopsis 9-cis epoxycarotenoid dioxygenase gene family. Plant J. 2003, 35, 44–56. [Google Scholar] [CrossRef]
- Sekimoto, H.; Seo, M.; Dohmae, N.; Takio, K.; Kamiya, Y.; Koshiba, T. Cloning and molecular characterization of plant aldehyde oxidase. J. Biol. Chem. 1997, 272, 15280–15285. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Peng, L.; Dong, S.; Xia, X.; Zhao, L. Transcriptome analysis of Chelidonium majus elaiosomes and seeds provide insights into fatty acid biosynthesis. PeerJ. 2019, 7, e6871. [Google Scholar] [CrossRef]
- Pourmazaheri, H.; Soorni, A.; Kohnerouz, B.B.; Dehaghi, N.K.; Kalantar, E.; Omidi, M.; Naghavi, M.R. Comparative analysis of the root and leaf transcriptomes in Chelidonium majus L. PLoS ONE 2019, 14, e0215165. [Google Scholar] [CrossRef]
- Lopez-Emparan, A.; Quezada-Martinez, D.; Zuniga-Bustos, M.; Cifuentes, V.; Iniguez-Luy, F.; Federico, M.L. Functional analysis of the Brassica napus L. phytoene synthase (PSY) gene family. PLoS ONE 2014, 9, e114878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, N.; Pandey, A.; Kumar, P.; Pandey, P.; Kesarwani, A.K.; Mantri, S.S.; Awasthi, P.; Tiwari, S. Regulation of banana phytoene synthase (MaPSY) expression, characterization and their modulation under various abiotic stress conditions. Front. Plant Sci. 2017, 8, 462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.-T.; Jia, L.-D.; Duan, M.-Z.; Chen, X.; Qiao, C.-L.; Ma, J.-Q.; Zhang, C.; Jing, F.-Y.; Zhang, S.-S.; Yang, B. Genome-wide identification and expression profiling of the carotenoid cleavage dioxygenase (CCD) gene family in Brassica napus L. PLoS ONE 2020, 15, e0238179. [Google Scholar]
- Zhe, Q.L.; Zheng, J.L.; Liu, J.H. Transcription activation of beta-carotene biosynthetic genes at the initial stage of stresses as an indicator of the increased β-carotene accumulation in isolated Dunaliella salina strain GY-H13. Aquat. Toxicol. 2020, 222, 105472. [Google Scholar] [CrossRef]
- Han, Y.; Zheng, Q.S.; Wei, Y.P.; Chen, J.; Liu, R.; Wan, H.J. In silico identification and analysis of phytoene synthase genes in plants. Genet. Mol. Res. 2015, 14, 9412–9422. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.L.; Wang, Y.C.; Qin, S. Molecular evolution of lycopene cyclases involved in the formation of carotenoids in eukaryotic algae. Plant Mol. Biol. Rep. 2011, 29, 1013–1020. [Google Scholar] [CrossRef]
- Zhu, Y.H.; Jiang, J.G.; Chen, Q. Characterization of cDNA of lycopene beta-cyclase responsible for a high level of beta-carotene accumulation in Dunaliella salina. Biochem. Cell Biol. 2008, 86, 285–292. [Google Scholar] [CrossRef]
- Zhu, Y.H.; Jiang, J.G.; Yan, Y.; Chen, X.W. Isolation and characterization of phytoene desaturase cDNA involved in the beta-carotene biosynthetic pathway in Dunaliella salina. J. Agric. Food Chem. 2005, 53, 5593–5597. [Google Scholar] [CrossRef]
- DePristo, M.A.; Weinreich, D.M.; Hartl, D.L. Missense meanderings in sequence space: A biophysical view of protein evolution. Nat. Rev. Genet. 2005, 6, 678–687. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Jhanwar, S.; Tyagi, A.K.; Jain, M. Genome-wide survey and expression analysis suggest diverse roles of glutaredoxin gene family members during development and response to various stimuli in rice. DNA Res. 2010, 17, 353–367. [Google Scholar] [CrossRef] [Green Version]
- Yan, P.; Gao, X.Z.; Shen, W.T.; Zhou, P. Cloning and expression analysis of phytoene desaturase and ζ-carotene desaturase genes in Carica papaya. Mol. Biol. Rep. 2011, 38, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Hugueney, P.; Badillo, A.; Chen, H.C.; Klein, A.; Hirschberg, J.; Camara, B.; Kuntz, M. Metabolism of cyclic carotenoids—A model for the alteration of this biosynthetic pathway in Capsicum annuum chromoplasts. Plant J. 1995, 8, 417–424. [Google Scholar] [CrossRef]
- Lao, Y.M.; Jin, H.; Zhou, J.; Zhang, H.J.; Cai, Z.H. Functional characterization of a missing branch component in Haematococcus pluvialis for control of algal carotenoid biosynthesis. Front. Plant Sci. 2017, 8, 1341. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouvier, F.; Keller, Y.; D’Harlingue, A.; Camara, B. Xanthophyll biosynthesis: Molecular and functional characterization of carotenoid hydroxylases from pepper fruits (Capsicum annuum L.). Biochim. Biophys. Acta Lipids Lipid Met. 1998, 1391, 320–328. [Google Scholar] [CrossRef]
- Charron, J.-B.F.; Ouellet, F.; Pelletier, M.; Danyluk, J.; Chauve, C.; Sarhan, F. Identification, expression, and evolutionary analyses of plant lipocalins. Plant Physiol. 2005, 139, 2017–2028. [Google Scholar] [CrossRef] [Green Version]
- Arnoux, P.; Morosinotto, T.; Saga, G.; Bassi, R.; Pignol, D. A structural basis for the pH-dependent xanthophyll cycle in Arabidopsis thaliana. Plant Cell 2009, 21, 2036–2044. [Google Scholar] [CrossRef] [Green Version]
- Bouvier, F.; Camara, B. Molecular analysis of carotenoid cyclase inhibition. Arch. Biochem. Biophys. 1997, 346, 53–64. [Google Scholar] [CrossRef]
- Mialoundama, A.S.; Heintz, D.; Jadid, N.; Nkeng, P.; Rahier, A.; Deli, J.; Camara, B.; Bouvier, F. Characterization of plant carotenoid cyclases as members of the flavoprotein family functioning with no net redox change. Plant Physiol. 2010, 153, 970–979. [Google Scholar] [CrossRef] [Green Version]
- Messing, S.A.J.; Gabelli, S.B.; Echeverria, I.; Vogel, J.T.; Guan, J.C.; Tan, B.C.; Klee, H.J.; McCarty, D.R.; Amzel, L.M. Structural insights into maize viviparous14, a key enzyme in the biosynthesis of the phytohormone abscisic acid. Plant Cell 2010, 22, 2970–2980. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, S.H.; Tan, B.C.; Gage, D.A.; Zeevaart, J.A.D.; McCarty, D.R. Specific oxidative cleavage of carotenoids by VP14 of maize. Science 1997, 276, 1872–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zdunek-Zastocka, E. Molecular cloning, characterization and expression analysis of three aldehyde oxidase genes from Pisum sativum L. Plant Physiol. Biochem. 2008, 46, 19–28. [Google Scholar] [CrossRef]
- Joyard, J.; Ferro, M.; Masselon, C.; Seigneurin-Berny, D.; Salvi, D.; Garin, J.; Rolland, N. Chloroplast proteomics and the compartmentation of plastidial isoprenoid biosynthetic pathways. Mol. Plant 2009, 2, 1154–1180. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Zhai, H.; Xue, L.Y.; Zhao, N.; He, S.Z.; Liu, Q.C. A lycopene β-cyclase gene, IbLCYB2, enhances carotenoid contents and abiotic stress tolerance in transgenic sweetpotato. Plant Sci. 2018, 272, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Li, P.R.; Zhang, S.J.; Zhang, S.F.; Li, F.; Zhang, H.; Cheng, F.; Wu, J.; Wang, X.W.; Sun, R.F. Carotenoid biosynthetic genes in Brassica rapa: Comparative genomic analysis, phylogenetic analysis, and expression profiling. BMC Genom. 2015, 16, 492. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Sola, M.A.; Rodriguez-Concepcion, M. Carotenoid biosynthesis in Arabidopsis: A colorful pathway. Arab. Book 2012, 10, e0158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuong, D.M.; Arasu, M.V.; Jeon, J.; Park, Y.J.; Kwon, S.J.; Al-Dhabi, N.A.; Park, S.U. Medically important carotenoids from Momordica charantia and their gene expressions in different organs. Saudi J. Biol. Sci. 2017, 24, 1913–1919. [Google Scholar] [CrossRef] [PubMed]
- Tuan, P.A.; Kim, J.K.; Kim, H.H.; Lee, S.Y.; Park, N.I.; Park, S.U. Carotenoid accumulation and characterization of cDNAs encoding phytoene synthase and phytoene desaturase in Garlic (Allium sativum). J. Agric. Food Chem. 2011, 59, 5412–5417. [Google Scholar] [CrossRef]
- Tuan, P.A.; Kim, J.K.; Park, N.I.; Lee, S.Y.; Park, S.U. Carotenoid content and expression of phytoene synthase and phytoene desaturase genes in bitter melon (Momordica charantia). Food Chem. 2011, 126, 1686–1692. [Google Scholar] [CrossRef]
- Tuan, P.A.; Kim, J.K.; Lee, J.; Park, W.T.; Kwon, D.Y.; Kim, Y.B.; Kim, H.H.; Kim, H.R.; Park, S.U. Analysis of carotenoid accumulation and expression of carotenoid biosynthesis genes in different organs of Chinese cabbage (Brassica rapa Subsp. pekinensis). Exp. Clin. Sci. J. 2012, 11, 508–516. [Google Scholar]
- Pillai, S.; Behra, R.; Nestler, H.; Suter, M.J.F.; Sigg, L.; Schirmer, K. Linking toxicity and adaptive responses across the transcriptome, proteome, and phenotype of Chlamydomonas reinhardtii exposed to silver. Proc. Natl. Acad. Sci. USA 2014, 111, 3490–3495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwanhausser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global quantification of mammalian gene expression control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Bai, C.; Bassie, L.; Nogareda, C.; Romagosa, I.; Twyman, R.M.; Christou, P.; Zhu, C. Zm PBF and Zm GAMYB transcription factors independently transactivate the promoter of the maize (Zea mays) β-carotene hydroxylase 2 gene. New Phytol. 2019, 222, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Koul, A.; Sharma, D.; Kaul, S.; Dhar, M.K. Identification and in silico characterization of cis-acting elements of genes involved in carotenoid biosynthesis in tomato. 3 Biotech 2019, 9, 287. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.L.; Guo, S.N.; Wen, F.; Zhang, X.L.; Wang, C.C.; Si, L.F.; Zheng, J.L.; Liu, J.H. Transcriptional and physiological responses of Dunaliella salina to cadmium reveals time-dependent turnover of ribosome, photosystem, and ROS-scavenging pathways. Aquat. Toxicol. 2019, 207, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Amiour, N.; Imbaud, S.; Clément, G.; Agier, N.; Zivy, M.; Valot, B.; Balliau, T.; Armengaud, P.; Quilleré, I.; Cañas, R. The use of metabolomics integrated with transcriptomic and proteomic studies for identifying key steps involved in the control of nitrogen metabolism in crops such as maize. J. Exp. Bot. 2012, 63, 5017–5033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balazadeh, S.; Schildhauer, J.; Araújo, W.L.; Munné-Bosch, S.; Fernie, A.R.; Proost, S.; Humbeck, K.; Mueller-Roeber, B. Reversal of senescence by N resupply to N-starved Arabidopsis thaliana: Transcriptomic and metabolomic consequences. J. Exp. Bot. 2014, 65, 3975–3992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, P.E.; Sreedasyam, A.; Trivedi, G.; Desai, S.; Dai, Y.; Cseke, L.J.; Collart, F.R. Multi-Omics approach identifies molecular mechanisms of plant-fungus mycorrhizal interaction. Front. Plant Sci. 2016, 6, 1061. [Google Scholar] [CrossRef]
- Maruyama, K.; Urano, K.; Yoshiwara, K.; Morishita, Y.; Sakurai, N.; Suzuki, H.; Kojima, M.; Sakakibara, H.; Shibata, D.; Saito, K. Integrated analysis of the effects of cold and dehydration on rice metabolites, phytohormones, and gene transcripts. Plant Physiol. 2014, 164, 1759–1771. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Zeng, D.; Yu, S.; Cui, C.; Li, J.; Li, H.; Chen, J.; Zhang, R.; Zhao, X.; Chen, L. From golden rice to aSTARice: Bioengineering astaxanthin biosynthesis in rice endosperm. Mol. Plant 2018, 11, 1440–1448. [Google Scholar] [CrossRef] [Green Version]
- Shumskaya, M.; Wurtzel, E.T. The carotenoid biosynthetic pathway: Thinking in all dimensions. Plant Sci. 2013, 208, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.; Tosatto, S.C.; Paladin, L.; Raj, S.; Richardson, L.J. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Geourjon, C.; Deleage, G. SOPMA: Significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments. Bioinformatics 1995, 11, 681–684. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins 2006, 64, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Emanuelsson, O.; Nielsen, H.; Heijne, G.V. ChloroP, a neural network-based method for predicting chloroplast transit peptides and their cleavage sites. Protein Sci. 1999, 8, 978–984. [Google Scholar] [CrossRef] [Green Version]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; Von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef] [Green Version]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.Y.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method—A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Park, C.H.; Xu, H.; Yeo, H.J.; Park, Y.E.; Hwang, G.-S.; Park, N.I.; Park, S.U. Enhancement of the flavone contents of Scutellaria baicalensis hairy roots via metabolic engineering using maize Lc and Arabidopsis PAP1 transcription factors. Metab. Eng. 2021, 64, 64–73. [Google Scholar] [CrossRef]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Park, C.H.; Yeo, H.J.; Park, S.Y.; Kim, J.K.; Park, S.U. Comparative phytochemical analyses and metabolic profiling of different phenotypes of Chinese cabbage (Brassica rapa ssp. pekinensis). Foods 2019, 8, 587. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Names | NCBI Accession No. | ORF (bp) | Length (aa) | ORF Type | MW (Kda) | pI | Signal Peptide |
|---|---|---|---|---|---|---|---|
| CmPSY | MW307341 | 912 | 303 | Full | 36.36 | 9.56 | No |
| CmPDS | MW307330 | 1752 | 583 | Full | 69.96 | 6.81 | No |
| CmZ-ISO | MW307331 | 734 | 244 | Partial | 29.28 | 6.71 | No |
| CmZDS | MW307344 | 1797 | 598 | Full | 71.76 | 7.89 | No |
| CmCrtISO | MW307336 | 1812 | 603 | Full | 72.36 | 8.79 | No |
| CmLCYB | MW307338 | 1518 | 505 | Full | 60.60 | 7.64 | No |
| CmLCYE | MW307339 | 1605 | 534 | Full | 64.08 | 6.49 | No |
| CmCHXB | MW307333 | 948 | 317 | Full | 38.04 | 8.75 | No |
| CmCHXE | MW307335 | 881 | 293 | Partial | 35.16 | 9.12 | No |
| CmZEP | MW307343 | 1998 | 665 | Full | 79.80 | 6.59 | No |
| CmVDE | MW307342 | 1464 | 487 | Full | 58.44 | 5.13 | No |
| CmCCS | MW307334 | 389 | 129 | Partial | 15.48 | 10.22 | No |
| CmCCD | MW307337 | 1836 | 611 | Full | 73.32 | 6.23 | No |
| CmNCED | MW307340 | 549 | 183 | Partial | 21.96 | 10.40 | No |
| CmAO | MW307332 | 4125 | 1374 | Full | 164.88 | 6.34 | No |
| Species Name | PSY | PDS | Z-ISO | ZDS | CrtISO | LCYB | LCYE | CHXB | CHXE | ZEP | VDE | CCS | CCD | NCED | AO |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Higher plants | |||||||||||||||
| C. majus | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| A. aestivalis | 72.46 | ||||||||||||||
| C. sinensis | 82.30 | 85.32 | 77.50 | 71.55 | |||||||||||
| M. cordata | 89.77 | ||||||||||||||
| P. somniferum | 70.26 | 78.00 | 81.40 | 81.60 | 82.56 | 76.39 | 70.65 | 79.53 | 68.05 | ||||||
| Chlorophyta | |||||||||||||||
| C. primus | 55.63 | 51.23 | 41.49 | 48.95 | 40.41 | ||||||||||
| C. reinhardtii | 63.60 | 56.36 | 55.93 | 44.96 | 49.08 | 30.13 | 30.72 | ||||||||
| M. pusilla | 27.78% | ||||||||||||||
| O. tauri | 38.70 | ||||||||||||||
| Bacteria | |||||||||||||||
| Calothrix sp. | 36.36 | ||||||||||||||
| D. bacterium | 35.66 | 44.03 | 32.86 | ||||||||||||
| Geitlerinema sp. | 66.03 | 35.82 | |||||||||||||
| O. cyanobacterium | 61.73 | 59.92 | |||||||||||||
| R. bacterium | 35.00 | ||||||||||||||
| Spirulina sp. | 46.00 | ||||||||||||||
| Dinoflagellates | |||||||||||||||
| S. microadriaticum | 40.50 | 25.27 | 33.20 | 35.62 | 25.44 | 27.39 | |||||||||
| Heterokonts | |||||||||||||||
| E. siliculosus | 49.65 | 28.23 | 35.78 | 32.15 | 40.44 | 23.63 | |||||||||
| F. cylindrus | 27.24 | 29.06 | 31.00 | ||||||||||||
| H. fermentalgiana | 33.16 | ||||||||||||||
| S. lomentaria | 59.75 |
| Gene Names | CELLO2GO | WoLF PSORT | TargetP | ChloroP 1.1 | Plant-PLoc | Consensus Prediction |
|---|---|---|---|---|---|---|
| CmPSY | MC | MC | Other | Other | NUC | MC/NUS/other |
| CmPDS | CP | CP | CP | CP | CP | CP |
| CmZ-ISO | PM | CP | CP | CP | CP | CP/PM |
| CmZDS | CP | CP | CP | CP | CP | CP |
| CmCrtISO | CP | CP | CP | CP | CP | CP |
| CmLCYB | MC | CP | Other | Other | CP | CP/MC/other |
| CmLCYE | PM | CP | CP | CP | CP | CP/PM |
| CmCHXB | CP | CP | CP | CP | CP | CP |
| CmCHXE | CP | CP | CP | CP | CP | CP |
| CmZEP | CP | Cytoplasmic | CP | Other | CP | CP/cytoplasmic/other |
| CmVDE | Cytoplasmic | ER | Thylakoid | Other | CP | CP/ER/thylakoid/other |
| CmCCS | MC | CP/MC | CP | CP | CP | CP/MC |
| CmCCD | CP | Cytoplasmic | Other | CP | CP | CP/cytoplasmic/other |
| CmNCED | NUC | CP | Other | CP | CP | CP/NUC/other |
| CmAO | CP | Cytoplasmic | Other | Other | CP | CP/cytoplasmic/other |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sathasivam, R.; Yeo, H.J.; Park, C.H.; Choi, M.; Kwon, H.; Sim, J.E.; Park, S.U.; Kim, J.K. Molecular Characterization, Expression Analysis of Carotenoid, Xanthophyll, Apocarotenoid Pathway Genes, and Carotenoid and Xanthophyll Accumulation in Chelidonium majus L. Plants 2021, 10, 1753. https://doi.org/10.3390/plants10081753
Sathasivam R, Yeo HJ, Park CH, Choi M, Kwon H, Sim JE, Park SU, Kim JK. Molecular Characterization, Expression Analysis of Carotenoid, Xanthophyll, Apocarotenoid Pathway Genes, and Carotenoid and Xanthophyll Accumulation in Chelidonium majus L. Plants. 2021; 10(8):1753. https://doi.org/10.3390/plants10081753
Chicago/Turabian StyleSathasivam, Ramaraj, Hyeon Ji Yeo, Chang Ha Park, Minsol Choi, Haejin Kwon, Ji Eun Sim, Sang Un Park, and Jae Kwang Kim. 2021. "Molecular Characterization, Expression Analysis of Carotenoid, Xanthophyll, Apocarotenoid Pathway Genes, and Carotenoid and Xanthophyll Accumulation in Chelidonium majus L." Plants 10, no. 8: 1753. https://doi.org/10.3390/plants10081753
APA StyleSathasivam, R., Yeo, H. J., Park, C. H., Choi, M., Kwon, H., Sim, J. E., Park, S. U., & Kim, J. K. (2021). Molecular Characterization, Expression Analysis of Carotenoid, Xanthophyll, Apocarotenoid Pathway Genes, and Carotenoid and Xanthophyll Accumulation in Chelidonium majus L. Plants, 10(8), 1753. https://doi.org/10.3390/plants10081753